
Investigating Internalization of Reporter-Protein-Functionalized Polyhedrin Particles by Brain Immune Cells
 ,
,
Abstract
:1. Introduction
- (1)
- To establish the extent to which PODS are internalised by microglia;
- (2)
- To assess whether acute toxic effects or microglial activation are induced by PODS internalisation;
- (3)
- To document intracellular localisation of PODS.
2. Materials and Methods
2.1. Materials
2.2. Characterisation of PODS Particles
2.3. Primary Mouse Microglia Culture
2.4. Incubation of PODS with Microglia
2.5. Fixation and Immunostaining of Cultures
2.6. Microscopy and Image Analysis
2.7. Statistical Analyses
3. Results
3.1. Characterisation of PODS Particles
3.2. No Acute Toxicity Was Evident in Microglia Incubated with PODS Particles
3.3. Morphometry of Microglia Incubated with PODS Was Unchanged versus Controls
3.4. Perinuclear PODS Occasionally Distorted Cell Nuclei
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kang, T.; Cha, G.D.; Park, O.K.; Cho, H.R.; Kim, M.; Lee, J.; Kim, D.; Lee, B.; Chu, J.; Koo, S.; et al. Penetrative and Sustained Drug Delivery Using Injectable Hydrogel Nanocomposites for Postsurgical Brain Tumor Treatment. ACS Nano 2023, 17, 5435–5447. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Mehta, A.; Tong, Z.; Esser, L.; Voelcker, N.H. Development of Polymeric Nanoparticles for Blood–Brain Barrier Transfer—Strategies and Challenges. Adv. Sci. 2021, 8, 1–32. [Google Scholar] [CrossRef] [PubMed]
- Bahlakeh, G.; Rahbarghazi, R.; Mohammadnejad, D.; Abedelahi, A.; Karimipour, M. Current knowledge and challenges associated with targeted delivery of neurotrophic factors into the central nervous system: Focus on available approaches. Cell Biosci. 2021, 11, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Robinson, J.; Lu, P. Optimization of trophic support for neural stem cell grafts in sites of spinal cord injury. Exp. Neurol. 2017, 291, 87–97. [Google Scholar] [CrossRef] [PubMed]
- Marsh, S.E.; Blurton-Jones, M. Neural stem cell therapy for neurodegenerative disorders: The role of neurotrophic support. Neurochem. Int. 2017, 106, 94–100. [Google Scholar] [CrossRef] [PubMed]
- Thorne, R.G.; Frey, W.H. Delivery of neurotrophic factors to the central nervous system: Pharmacokinetic considerations. Clin. Pharmacokinet. 2001, 40, 907–946. [Google Scholar] [CrossRef] [PubMed]
- Saltzman, W.M.; Mak, M.W.; Mahoney, M.J.; Duenas, E.T.; Cleland, J.L. Intracranial delivery of recombinant nerve growth factor: Release kinetics and protein distribution for three delivery systems. Pharm. Res. 1999, 16, 232–240. [Google Scholar] [CrossRef]
- Krewson, C.E.; Klarman, M.L.; Saltzman, W.M. Distribution of nerve growth factor following direct delivery to brain interstitium. Brain Res. 1995, 680, 196–206. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Cooke, M.J.; Sachewsky, N.; Morshead, C.M.; Shoichet, M.S. Bioengineered sequential growth factor delivery stimulates brain tissue regeneration after stroke. J. Control. Release 2013, 172, 1–11. [Google Scholar] [CrossRef]
- Van Damme, P.; Tilkin, P.; Mercer, K.J.; Terryn, J.; D’Hondt, A.; Herne, N.; Tousseyn, T.; Claeys, K.G.; Thal, D.R.; Zachrisson, O.; et al. Intracerebroventricular delivery of vascular endothelial growth factor in patients with amyotrophic lateral sclerosis, a phase I study. Brain Commun. 2020, 2, 1–10. [Google Scholar] [CrossRef]
- Eriksdotter Jönhagen, M.; Nordberg, A.; Amberla, K.; Bäckman, L.; Ebendal, T.; Meyerson, B.; Olson, L.; Seiger, Å.; Shigeta, M.; Theodorsson, E.; et al. Intracerebroventricular Infusion of Nerve Growth Factor in Three Patients with Alzheimer’s Disease. Dement. Geriatr. Cogn. Disord. 1998, 9, 246–257. [Google Scholar] [CrossRef] [PubMed]
- Emerich, D.F.; Kordower, J.H.; Chu, Y.; Thanos, C.; Bintz, B.; Paolone, G.; Wahlberg, L.U. Widespread Striatal Delivery of GDNF from Encapsulated Cells Prevents the Anatomical and Functional Consequences of Excitotoxicity. Neural Plast. 2019, 2019, 6286197. [Google Scholar] [CrossRef]
- Chang, H.; Heuer, R.A.; Oleksijew, A.M.; Coots, K.S.; Roque, C.B.; Nella, K.T.; McGuire, T.L.; Matsuoka, A.J. An engineered three-dimensional stem cell niche in the inner ear by applying a nanofibrillar cellulose hydrogel with a sustained-release neurotrophic factor delivery system. Acta Biomater. 2020, 108, 111–127. [Google Scholar] [CrossRef] [PubMed]
- Géral, C.; Angelova, A.; Lesieur, S. From molecular to nanotechnology strategies for delivery of neurotrophins: Emphasis on brain-derived neurotrophic factor (BDNF). Pharmaceutics 2013, 5, 127–167. [Google Scholar] [CrossRef]
- Coulibaly, F.; Chiu, E.; Ikeda, K.; Gutmann, S.; Haebel, P.W.; Schulze-Briese, C.; Mori, H.; Metcalf, P. The molecular organization of cypovirus polyhedra. Nature 2007, 446, 97–101. [Google Scholar] [CrossRef]
- Ji, X.; Sutton, G.; Evans, G.; Axford, D.; Owen, R.; Stuart, D.I. How baculovirus polyhedra fit square pegs into round holes to robustly package viruses. EMBO J. 2010, 29, 505–514. [Google Scholar] [CrossRef] [PubMed]
- Mori, H. Expression of Bombyx movi cytoplasmic polyhedrosis virus polyhedrin in insect cells by using a baculovirus expression vector, and its assembly into polyhedra Polyhedrin. J. Gen. Virol. 1993, 74, 99–102. [Google Scholar] [CrossRef] [PubMed]
- Healy, A.M.; Worku, Z.A.; Kumar, D.; Madi, A.M. Pharmaceutical solvates, hydrates and amorphous forms: A special emphasis on cocrystals. Adv. Drug Deliv. Rev. 2017, 117, 25–46. [Google Scholar] [CrossRef]
- Coulibaly, F.; Chiu, E.; Gutmann, S.; Rajendran, C.; Haebel, P.W.; Ikeda, K.; Mori, H.; Ward, V.K.; Schulze-Briese, C.; Metcalf, P. The atomic structure of baculovirus polyhedra reveals the independent emergence of infectious crystals in DNA and RNA viruses. Proc. Natl. Acad. Sci. USA 2009, 106, 22205–22210. [Google Scholar] [CrossRef]
- Rohrmann, G.F. Polyhedrin Structure. J. Gen. Virol. 1986, 67, 1499–1513. [Google Scholar] [CrossRef]
- Abe, S.; Maity, B.; Ueno, T. Design of a confined environment using protein cages and crystals for the development of biohybrid materials. Chem. Commun. 2016, 52, 6496–6512. [Google Scholar] [CrossRef] [PubMed]
- Chiu, E.; Coulibaly, F.; Metcalf, P. Insect virus polyhedra, infectious protein crystals that contain virus particles. Curr. Opin. Struct. Biol. 2012, 22, 234–240. [Google Scholar] [CrossRef] [PubMed]
- Mori, H.; Shukunami, C.; Furuyama, A.; Notsu, H.; Nishizaki, Y.; Hiraki, Y. Immobilization of Bioactive Fibroblast Growth Factor-2 into Cubic Proteinous Microcrystals (Bombyx mori Cypovirus Polyhedra) That Are Insoluble in a Physiological Cellular Environment. J. Biol. Chem. 2007, 282, 17289–17296. [Google Scholar] [CrossRef] [PubMed]
- Wendler, A.; James, N.; Jones, M.H.; Pernstich, C. Phagocytosed Polyhedrin-Cytokine Cocrystal Nanoparticles Provide Sustained Secretion of Bioactive Cytokines from Macrophages. BioDesign Res. 2021, 2021, 9816485. [Google Scholar] [CrossRef] [PubMed]
- Whitty, C.; Pernstich, C.; Marris, C.; McCaskie, A.; Jones, M.; Henson, F. Sustained delivery of the bone morphogenetic proteins BMP-2 and BMP-7 for cartilage repair and regeneration in osteoarthritis. Osteoarthr. Cartil. Open 2022, 4, 100240. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, G.; Ueda, T.; Shimoyama, J.; Ijiri, H.; Omi, Y.; Yube, H.; Sugita, Y.; Kubo, K.; Maeda, H.; Kinoshita, Y.; et al. Bone regeneration by polyhedral microcrystals from silkworm virus. Sci. Rep. 2012, 2, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Matsuzaki, Y.; Maruta, R.; Takaki, K.; Kotani, E.; Kato, Y.; Yoshimura, R.; Endo, Y.; Whitty, C.; Pernstich, C.; Gandhi, R.; et al. Sustained neurotrophin release from protein nanoparticles mediated by matrix metalloproteinases induces the alignment and differentiation of nerve cells. Biomolecules 2019, 9, 510. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, G.A. Matrix Metalloproteinases and Extracellular Matrix in the Central Nervous System. In Primer on Cerebrovascular Diseases; Elsevier: Amsterdam, The Netherlands, 2017; pp. 291–295. ISBN 9780128030585. [Google Scholar]
- Nella, K.T.; Norton, B.M.; Chang, H.; Heuer, R.A.; Roque, C.B.; Matsuoka, A.J. Bridging the electrode-neuron gap: Finite element modeling of in vitro neurotrophin gradients to optimize neuroelectronic interfaces in the inner ear. Acta Biomater. 2022, 151, 360–378. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, S.I.; Weinberg, D.; Al-Shakli, A.F.; Fernandes, A.R.; Yiu, H.H.P.; Telling, N.D.; Roach, P.; Chari, D.M. ‘Stealth’ nanoparticles evade neural immune cells but also evade major brain cell populations: Implications for PEG-based neurotherapeutics. J. Control. Release 2016, 224, 136–145. [Google Scholar] [CrossRef]
- Tsui, C.; Koss, K.; Churchward, M.A.; Todd, K.G. Biomaterials and glia: Progress on designs to modulate neuroinflammation. Acta Biomater. 2019, 83, 13–28. [Google Scholar] [CrossRef]
- Basit, R.H.; Tzerakis, N.; Jenkins, S.I.; Chari, D.M. In vitro model of traumatic brain injury to screen neuro-regenerative biomaterials. Mater. Sci. Eng. C 2021, 128, 112253. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, S.I.; Pickard, M.R.; Furness, D.N.; Yiu, H.H.P.; Chari, D.M. Differences in magnetic particle uptake by CNS neuroglial subclasses: Implications for neural tissue engineering. Nanomedicine 2013, 8, 951–968. [Google Scholar] [CrossRef] [PubMed]
- Soenen, S.I.J.H.; De Cuyper, M. Assessing iron oxide nanoparticle toxicity in vitro: Current status and future prospects. Nanomedicine 2010, 5, 1261–1275. [Google Scholar] [CrossRef] [PubMed]
- Cell Guidance Systems PP303 PODS GFP Data Sheet. Available online: https://cellgs.e2ecdn.co.uk/Downloads/PP303.pdf (accessed on 23 January 2024).
- Fernandes, A.R.; Chari, D.M. A multicellular, neuro-mimetic model to study nanoparticle uptake in cells of the central nervous system. Integr. Biol. 2014, 6, 855–861. [Google Scholar] [CrossRef]
- McCarthy, K.D.; de Vellis, J. Preparation of separate astroglial and oligodendroglial cell cultures from rat cerebral tissue. J. Cell Biol. 1980, 85, 890–902. [Google Scholar] [CrossRef]
- Jenkins, S.I.; Roach, P.; Chari, D.M. Development of a nanomaterial bio-screening platform for neurological applications. Nanomedicine NBM 2014, 11, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, S.I.; Pickard, M.R.; Chari, D.M. Magnetic Nanoparticle Mediated Gene Delivery in Oligodendroglial Cells: A Comparison of Differentiated Cells Versus Precursor Forms. Nano Life 2012, 3, 1243001. [Google Scholar] [CrossRef]
- Hutter, E.; Boridy, S.; Labrecque, S.; Lalancette-Hébert, M.; Kriz, J.; Winnik, F.M.; Maysinger, D. Microglial Response to Gold Nanoparticles. ACS Nano 2010, 4, 2595–2606. [Google Scholar] [CrossRef]
- Canton, I.; Battaglia, G. Endocytosis at the nanoscale. Chem. Soc. Rev. 2012, 41, 2718–2739. [Google Scholar] [CrossRef]
- Cannon, G.J.; Swanson, J.A. The macrophage capacity for phagocytosis. J. Cell Sci. 1992, 101, 907–913. [Google Scholar] [CrossRef] [PubMed]
- Swanson, J.A.; Watts, C. Macropinocytosis. Trends Cell Biol. 1995, 5, 424–428. [Google Scholar] [CrossRef] [PubMed]
- Manzanares, D.; Ceña, V. Endocytosis: The nanoparticle and submicron nanocompounds gateway into the cell. Pharmaceutics 2020, 12, 371. [Google Scholar] [CrossRef] [PubMed]
- Rennick, J.J.; Johnston, A.P.R.; Parton, R.G. Key principles and methods for studying the endocytosis of biological and nanoparticle therapeutics. Nat. Nanotechnol. 2021, 16, 266–276. [Google Scholar] [CrossRef]
- Bannunah, A.M.; Vllasaliu, D.; Lord, J.; Stolnik, S. The mechanisms of nanoparticle internalization and transport across an intestinal epithelial cell model: Effect of size and surface charge. Mol. Pharm. 2014, 11, 4363–4373. [Google Scholar] [CrossRef]
- Ding, L.; Zhu, X.; Wang, Y.; Shi, B.; Ling, X.; Chen, H.; Nan, W.; Barrett, A.; Guo, Z.; Tao, W.; et al. Intracellular Fate of Nanoparticles with Polydopamine Surface Engineering and a Novel Strategy for Exocytosis-Inhibiting, Lysosome ImpairmentBased Cancer Therapy. Nano Lett. 2017, 17, 6790–6801. [Google Scholar] [CrossRef]




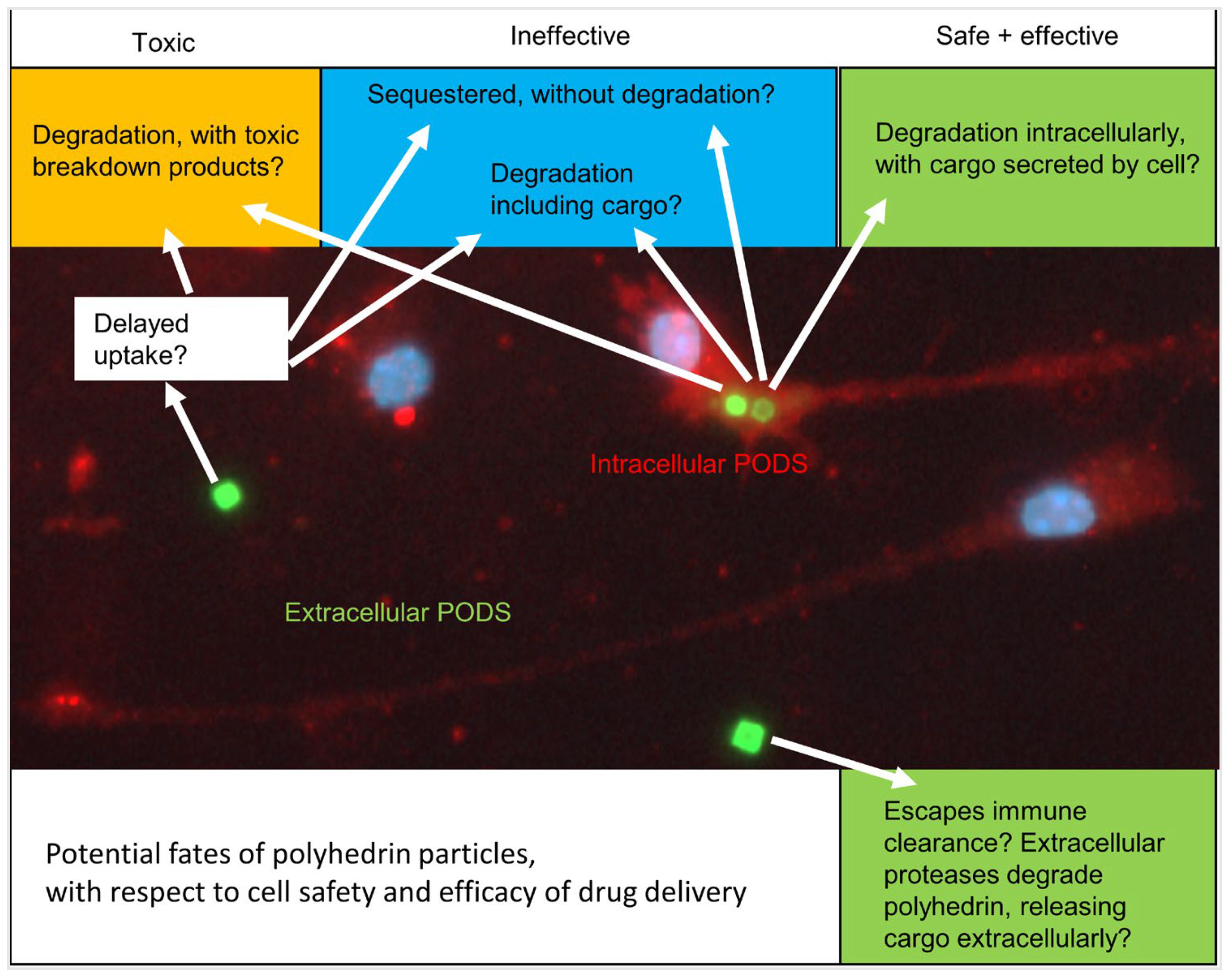

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Type, Source (Author) [Ref] | PODS Type/Cargo | PODS Concentration | Exposure Time (Days) | Uptake | Toxicity | Cell Responses to PODS (Other Notes) |
|---|---|---|---|---|---|---|
| Rat PC12 cell line (Matsuzaki) [27] | NGF, EGFP | 5 or 10 × 104 PODS/coverslip (7 × 104 cells) | 5 (no media change) | Not assessed | “no evidence of inflammatory or foreign-body reaction” in vivo (PODS in collagen scaffold, implanted into bone) | Did not address direct cell responses to PODS. Cells avoided dried patch of PODS, applied before cell seeding. Drug delivery seemed effective. (Reported protease-induced pores in PODS.) |
| hESC-derived otic neuronal progenitors (Chang) [13] | hBDNF | 80 × 104 PODS/well | 7 | Not assessed | Not directly assessed; no toxicity noted | Did not address direct cell responses to PODS Drug delivery seemed effective (consistent hBDNF release over 7 d, assessed by ELISA) |
| THP1-derived macrophages, M0, M1, M2 (Wendler) [24] | empty, EGFP, IL-6, FGF2, −10 | 5, 10 or 15 PODS/cell | 1, 4 | Yes | 15 PODS/cell did not show toxicity, up to 96 h; but “apoptotic bodies” at ~50 PODS/cell | All phenotypes (M0, M1, M2) showed uptake; ‘almost all PODS at 24 h’. No effects on cellular function reported. IL-6 secretion by M1 was unaltered by empty-/FGF10-PODS uptake (lack of inflammation). IL-6-PODS: IL-6 in media from cell culture was 8–30% of IL-6-PODS alone. |
| Mouse primary bone marrow monocytes (Wendler) [24] | M-CSF, GM-CSF | 5 or 10 PODS/cell | 1 | Yes | ”seemingly without negatively influencing their behavior” | Cargo protein remained bioactive after macrophage uptake; possibly secreted by macrophages? PODS suggested to survive acidic conditions of phagolysosomes. |
| Human chondrocytes; ‘non-professional phagocytes’ (Whitty) [25] | empty, BMP-2, -7 | 50 ng/mL, 25–200 ng/mL | 14 | Yes | No | Uptake reported as “phagocytosis”, although endocytotic mechanism not specifically assessed. Extent of uptake only assessed as ‘efficient’. (Increased proliferation when treated with PODS-BMP2 and -BMP7) |
| Spinal ganglion cells, from hiPSCs (Nella) [29] | hBDNF | 2 or 80 × 104 PODS/well | 7 | Not assessed | Not assessed | Drug delivery/release produced cellular responses. (Serum was required to release BDNF from PODS in cell-free conditions; Authors speculate that proteases are necessary for cargo release.) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Parwana, K.A.K.; Kaur Gill, P.; Njanike, R.; Yiu, H.H.P.; Adams, C.F.; Chari, D.M.; Jenkins, S.I. Investigating Internalization of Reporter-Protein-Functionalized Polyhedrin Particles by Brain Immune Cells. Materials 2024, 17, 2330. https://doi.org/10.3390/ma17102330
Parwana KAK, Kaur Gill P, Njanike R, Yiu HHP, Adams CF, Chari DM, Jenkins SI. Investigating Internalization of Reporter-Protein-Functionalized Polyhedrin Particles by Brain Immune Cells. Materials. 2024; 17(10):2330. https://doi.org/10.3390/ma17102330
Chicago/Turabian StyleParwana, Krishma A. K., Priyapreet Kaur Gill, Runyararo Njanike, Humphrey H. P. Yiu, Chris F. Adams, Divya Maitreyi Chari, and Stuart Iain Jenkins. 2024. "Investigating Internalization of Reporter-Protein-Functionalized Polyhedrin Particles by Brain Immune Cells" Materials 17, no. 10: 2330. https://doi.org/10.3390/ma17102330