
Evaluating Different Methods to Establish Biodiverse Swards of Native Grasses and Wildflowers for Pasturelands
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sites
2.2. Experimental Design
2.3. Measurements
2.4. Data Analysis
3. Results
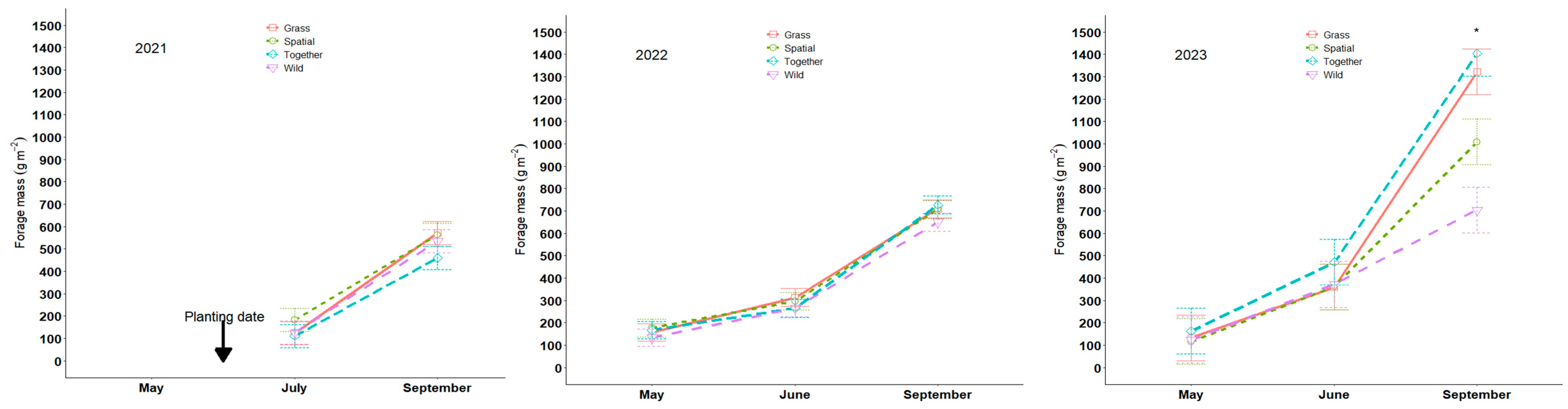
3.1. Stand Density and Forage Mass
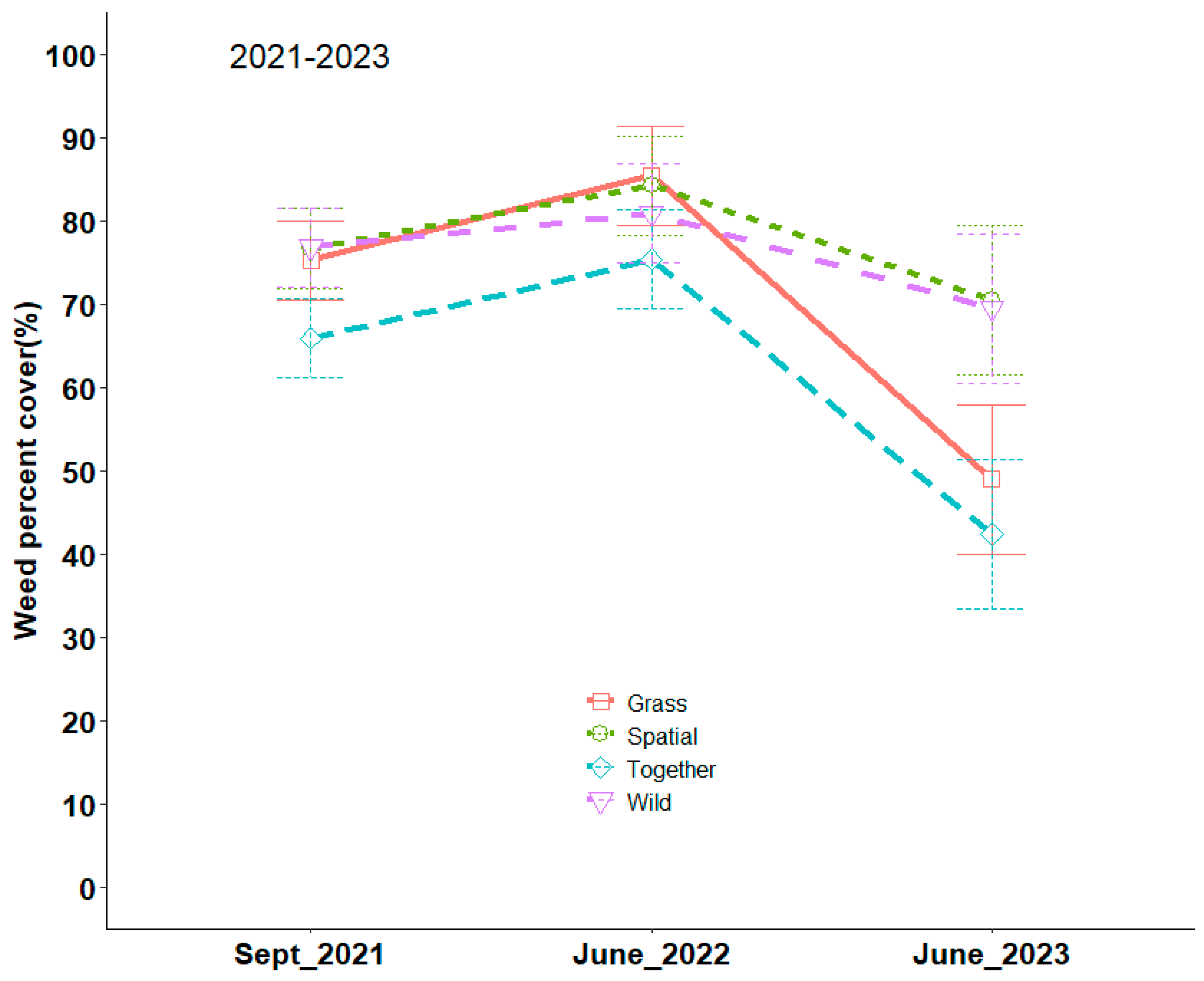
3.2. Species Composition
3.3. Bloom Density
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hoveland, C.S. Importance and economic significance of the Acremonium endophytes to performance of animals and grass plant. Agric. Ecosyst. Environ. 1993, 44, 3–12. [Google Scholar] [CrossRef]
- Tracy, B.; Maughan, M.; Post, N.; Faulkner, D. Integrating Annual and Perennial Warm-season Grasses in a Temperate Grazing System. Crop Sci. 2010, 50, 2171–2177. [Google Scholar] [CrossRef]
- Keyser, P.D.; Ashworth, A.J.; Allen, F.L.; Bates, G.E. Dormant-Season Planting and Seed-Dormancy Impacts on Switchgrass Establishment and Yield. Crop Sci. 2016, 56, 474–483. [Google Scholar] [CrossRef]
- Keyser, P.D.; Buehler, D.A.; Fike, J.H.; Finke, D.L.; Fuhlendorf, S.D.; Martin, J.A.; Naumann, H.D.; Smith, S.R. The Birds and the Bees: Producing Beef and Conservation Benefits on Working Grasslands. Agronomy 2022, 12, 1934. [Google Scholar] [CrossRef]
- Sollenberger, L.E.; Kohmann, M.M.; Dubeux, J.C.B., Jr.; Silveira, M.L. Grassland Management Affects Delivery of Regulating and Supporting Ecosystem Services. Crop Sci. 2019, 59, 441–459. [Google Scholar] [CrossRef]
- Sanderson, M.A.; Liebig, M.A. Forages and the Environment. In Forages 2020; Moore, K.J., Collins, M., Nelson, C.J., Redfearn, D.D., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2020; pp. 249–259. [Google Scholar] [CrossRef]
- Biesmeijer, J.C.; Roberts, S.P.; Reemer, M.; Ohlemüller, R.; Edwards, M.; Peeters, T.; Schaffers, A.P.; Potts, S.G.; Kleukers, R.; Thomas, C.D.; et al. Parallel declines in pollinators and insect-pollinated plants in Britain and the Netherlands. Science 2006, 313, 351–354. [Google Scholar] [CrossRef]
- Savage, J.; Woodcock, B.A.; Bullock, J.M.; Nowakowski, M.; Tallowin, J.R.B.; Pywell, R.F. Management to Support Multiple Ecosystem Services from Productive Grasslands. Sustainability 2021, 13, 6263. [Google Scholar] [CrossRef]
- Ghajar, S.M.; Wagner, J.F.; O’Rourke, M.; Tracy, B.F. Evaluating methods to establish biodiverse pasturelands with native grasses and wildflowers. Nativ. Plants J. 2022, 23, 65–74. [Google Scholar] [CrossRef]
- Wagner, J.; Ghajar, S.; O’rourke, M.; Tracy, B. Bee-Friendly Beef: Developing Biodiverse Pastures to Increase Ecosystem Services. In Proceedings of the XXIV International Grassland Congress/XI International Rangeland Congress (Sustainable Use of Grassland and Rangeland Resources for Improved Livelihoods) 2021, Nairobi, Kenya, 25–29 October 2021. [Google Scholar]
- Richard, M.P.; Morrison, J.I.; McCurdy, J.D. Effects of preemergence herbicides on establishment of little bluestem and sideoats grama golf course rough. Crop Forage Turfgrass Manag. 2020, 6, e20051. [Google Scholar] [CrossRef]
- Angelella, G.M.; O’Rourke, M.E. Pollinator Habitat Establishment after Organic and No-till Seedbed Preparation Methods. HortScience 2017, 52, 1349–1355. [Google Scholar] [CrossRef]
- Angelella, G.M.; Stange, L.; Scoggins, H.L.; O’Rourke, M.E. Pollinator Refuge Establishment and Conservation Value: Impacts of Seedbed Preparations, Seed Mixtures, and Herbicides. HortScience 2019, 54, 445–451. [Google Scholar] [CrossRef]
- Dickson, T.L.; Busby, W.H. Forb Species Establishment Increases with Decreased Grass Seeding Density and with Increased Forb Seeding Density in a Northeast Kansas, U.S.A. Exp. Prairie Restoration. Restor. Ecol. 2009, 17, 597–605. [Google Scholar] [CrossRef]
- Jaramillo, D.M.; Dubeux, J.C.B., Jr.; Mackowiak, C.; Sollenberger, L.E.; DiLorenzo, N.; Rowland, D.L.; Blount, A.R.S.; Santos, E.R.S.; Garcia, L.; Ruiz-Moreno, M. Annual and Perennial Peanut Mixed with ‘Pensacola’ Bahiagrass in North Florida. Crop Sci. 2018, 58, 982–992. [Google Scholar] [CrossRef]
- Glidden, A.J.; Sherrard, M.E.; Meissen, J.C.; Myers, M.C.; Elgersma, K.J.; Jackson, L.L. Planting time, first-year mowing, and seed mix design influence ecological outcomes in agroecosystem revegetation projects. Restor. Ecol. 2023, 31, e13818. [Google Scholar] [CrossRef]
- Kimball, S.; Lulow, M.; Sorenson, Q.; Balazs, K.; Fang, Y.-C.; Davis, S.J.; O’Connell, M.; Huxman, T.E. Cost-effective ecological restoration. Restor. Ecol. 2015, 23, 800–810. [Google Scholar] [CrossRef]
- Vogel, K.P.; Masters, R.A. Frequency grid—A simple tool for measuring grassland establishment. Range Manag. 2001, 54, 653–655. [Google Scholar] [CrossRef]
- Woodcock, B.A.; Savage, J.; Bullock, J.M.; Nowakowski, M.; Orr, R.; Tallowin, J.R.B.; Pywell, R.F. Enhancing floral resources for pollinators in productive agricultural grasslands. Biol. Conserv. 2014, 171, 44–51. [Google Scholar] [CrossRef]
- Daubenmire, R.F. Canopy coverage method of vegetation analysis. Northwest Sci. 1959, 33, 43–67. [Google Scholar]
- Xu, S.; Jagadamma, S.; Cui, S.; Oakes, R.N.; Kubesch, J.O.C. Forage species composition influenced soil health in organic forage transitioning systems. Agric. Ecosyst. Environ. 2023, 342, 108228. [Google Scholar] [CrossRef]
- Seahra, S.; Yurkonis, K.A.; Newman, J.A. Seeding tallgrass prairie in monospecific patches promotes native species establishment and cover. Restor. Ecol. 2019, 27, 82–91. [Google Scholar] [CrossRef]
- Grman, E.; Zirbel, C.R.; Bauer, J.T.; Groves, A.M.; Bassett, T.; Brudvig, L.A. Super-abundant C4 grasses are a mixed blessing in restored prairies. Restor. Ecol. 2021, 29, e13281. [Google Scholar] [CrossRef]
- Robert, A.M.; Scott, J.N.; Roch, E.G.; Daniel, D.B.; Robert, N.S. Imidazolinone Herbicides Improve Restoration of Great Plains Grasslands. Weed Technol. 1996, 10, 392–403. [Google Scholar]
- Barnes, T.G. Strategies to Convert Exotic Grass Pastures to Tall Grass Prairie Communities. Weed Technol. 2004, 18, 1364–1370. [Google Scholar] [CrossRef]
- Wiese, J.L.; Keren, E.N.; Menalled, F.D. Tolerance of native wildflower species to postemergence herbicides. Nativ. Plants J. 2011, 12, 31–36. [Google Scholar] [CrossRef]
- Beran, D.D.; Gaussoin, R.E.; Masters, R.A. Native Wildflower Establishment with Imidazolinone Herbicides. HortScience 1999, 34, 283–286. [Google Scholar] [CrossRef]
- Li, Z.; Xu, S.; Rajan, N.; Nair, S.; Jagadamma, S.; Nave, R.; Kubesch, J.; Bates, G.; McIntosh, D.; Chen, C.; et al. Productivity and nutritive value of no-input minimum tillage organic forage systems. Nutr. Cycl. Agroecosyst. 2022, 124, 335–357. [Google Scholar] [CrossRef]
- Bonin, C.L.; Lal, R.; Tracy, B.F. Evaluation of Perennial Warm-Season Grass Mixtures Managed for Grazing or Biomass Production. Crop Sci. 2014, 54, 2373–2385. [Google Scholar] [CrossRef]
- Hong, C.O.; Owens, V.N.; Lee, D.K.; Boe, A. Switchgrass, Big Bluestem, and Indiangrass Monocultures and Their Two- and Three-Way Mixtures for Bioenergy in the Northern Great Plains. BioEnergy Res. 2013, 6, 229–239. [Google Scholar] [CrossRef]
- Gail, W.T.; Hartnett, D.C. Effects of mycorrhizae on plant growth and Dynamics in Experimental Tallgrass Prairie Microcosms. Am. J. Bot. 1997, 84, 478–482. [Google Scholar] [CrossRef]
- Wilson, G.W.T.; Hartnett, D.C. Interspecific variation in plant responses to mycorrhizal colonization in tallgrass prairie. Am. J. Bot. 1998, 85, 1732–1738. [Google Scholar] [CrossRef]
- Zhang, G.; Murray, C.J.; Clair, A.L.S.; Cass, R.P.; Dolezal, A.G.; Schulte, L.A.; Toth, A.L.; O’Neal, M.E. Native vegetation embedded in landscapes dominated by corn and soybean improves honey bee health and productivity. J. Appl. Ecol. 2023, 60, 1032–1043. [Google Scholar] [CrossRef]
- Kubesch, J. Evaluating Native Warm-Season Grass and Wildflower Mixtures for Beef Cattle Production in the Mid-Atlantic. Ph.D. Thesis, Virginia Tech, Blacksburg, VA, USA, 2023. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
| Location | pH | Buffer pH | P2O5 (mg kg−1) | K2O (mg kg−1) | Organic Matter (%) |
|---|---|---|---|---|---|
| Kentland | 6.6 | 6.4 | 27.5 | 66.0 | 3.7 |
| SVAREC | 6.4 | 6.2 | 20.5 | 36.5 | 7.2 |
| Species | 2021 | 2022 | 2023 |
|---|---|---|---|
| Big bluestem (Andropogon gerardii) | 0.39 (±0.53) | 2.05 (±1.57) | 6.02 (±3.68) |
| Little bluestem (Schizachyrium scoparium) | 0.00 (±0.00) | 1.86 (±1.15) | 3.42 (±1.66) |
| Indiangrass (Sorghastrum nutans) | 0.46 (±0.41) | 3.69 (±1.94) | Together: 24.38 (±4.29) A Grass and Spatial: 13.50 (±4.29) AB Wild: 1.38 (±4.29) B |
| Lanceleaf coreopsis (Coreopsis lanceolata) | 0.06 (±0.16) | 0.49 (±0.68) | 0.19 (±0.34) |
| Perennial blue flax (Linum perenne) | 0.00 (±0.00) | 1.27 (±1.50) | 0.33 (±0.46) |
| Ohio spiderwort (Tradescantia ohiensis) | 0.00 (±0.00) | 1.66 (±1.10) | 4.38 (±2.83) |
| Black-eyed Susan (Rudbeckia hirta) | 0.88 (±0.95) | 2.44 (±1.81) | 3.69 (±2.84) |
| Purple coneflower (Echinacea purpurea) | 0.56 (±0.71) | 1.25 (±0.89) | 1.42 (±1.14) |
| Anise hyssop (Agastache foeniculum) | 0.33 (±0.38) | 0.16 (±0.24) | 0.00 (±0.00) |
| Grey-headed coneflower (Ratibida pinnata) | 0.14 (±0.22) | 1.72 (±1.39) | 5.64 (±3.04) |
| Maximilian sunflower (Helianthus maximilliani) | 0.83 (±0.60) | 0.00 (±0.00) | 0.00 (±0.00) |
| Rigid goldenrod (Solidago rigida) | 0.00 (±0.00) | 0.03 (±0.04) | 0.39 (±0.56) |
| Annual gaillardia (Gaillardia pulchella) | 4.23 (±2.74) | 2.73 (±2.93) | 0.08 (±0.16) |
| Species | 2021 | 2022 | 2023 |
|---|---|---|---|
| Lanceleaf coreopsis (Coreopsis lanceolata) | 0.00 (±0.00) | 0.31 (±0.42) | 0.10 (±0.21) |
| Perennial blue flax (Linum perenne) | 0.00 (±0.00) | 0.85 (±0.88) | 0.58 (±0.76) |
| Ohio spiderwort (Tradescantia ohiensis) | 0.00 (±0.00) | 1.31 (±1.36) | 5.58 (±8.60) |
| Black-eyed Susan (Rudbeckia hirta) | 0.81 (±0.83) | 5.40 (±4.01) | 8.48 (±4.65) |
| Purple coneflower (Echinacea purpurea) | 0.00 (±0.00) | 0.21 (±0.25) | 0.35 (±0.44) |
| Anise hyssop (Agastache foeniculum) | 1.06 (±1.59) | 0.71 (±1.41) | 0.00 (±0.00) |
| Grey-headed coneflower (Ratibida pinnata) | 0.00 (±0.00) | 0.58 (±0.83) | 10.83 (±12.36) |
| Maximilian sunflower (Helianthus maximilliani) | 0.06 (±0.13) | 0.04 (±0.08) | 0.00 (±0.00) |
| Rigid goldenrod (Solidago rigida) | 0.00 (±0.00) | 0.00 (±0.00) | 1.56 (±3.11) |
| Annual gaillardia (Gaillardia pulchella) | 3.13 (±2.84) | 2.79 (±2.54) | 0.00 (±0.00) |
| White clover (Trifolium repens) | 0.00 (±0.00) | 0.21 (±0.50) | 0.00 (±0.00) |
| Red clover (Trifolium pratensis) | 0.00 (±0.00) | 0.00 (±0.00) | 0.00 (±0.00) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kubesch, J.O.C.; Greiner, S.P.; Pent, G.J.; Reid, J.L.; Tracy, B.F. Evaluating Different Methods to Establish Biodiverse Swards of Native Grasses and Wildflowers for Pasturelands. Agronomy 2024, 14, 1041. https://doi.org/10.3390/agronomy14051041
Kubesch JOC, Greiner SP, Pent GJ, Reid JL, Tracy BF. Evaluating Different Methods to Establish Biodiverse Swards of Native Grasses and Wildflowers for Pasturelands. Agronomy. 2024; 14(5):1041. https://doi.org/10.3390/agronomy14051041
Chicago/Turabian StyleKubesch, Jonathan O. C., Scott P. Greiner, Gabriel J. Pent, J. Leighton Reid, and Benjamin F. Tracy. 2024. "Evaluating Different Methods to Establish Biodiverse Swards of Native Grasses and Wildflowers for Pasturelands" Agronomy 14, no. 5: 1041. https://doi.org/10.3390/agronomy14051041