
Nicotinamide Supplementation Mitigates Oxidative Injury of Bovine Intestinal Epithelial Cells through Autophagy Modulation
 and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Cell Culture
2.3. Effect of Nicotinamide on Bovine Intestinal Epithelial Cells Mediated by H2O2
2.4. Cell Viability Assay
2.5. Determination of Lactate Dehydrogenase and Total Antioxidant Capacity
2.6. RNA Extraction and Real-Time Quantitative PCR
2.7. Protein Extraction and Western Blotting
2.8. Detection of Autophagy and Mitochondrial Membrane Potential
2.9. Immunofluorescence
2.10. Inhibition of Autophagy in Bovine Intestinal Epithelial Cells
2.11. Statistical Analysis
3. Results
3.1. Effect of Nicotinamide on Oxidative Stress Bovine Intestinal Epithelial Cells Challenged by H2O2
3.2. Antioxidant Enzymes
3.3. Tight Junction Proteins
3.4. Effect of Nicotinamide on Autophagy of Bovine Intestinal Epithelial Cells Challenged by H2O2
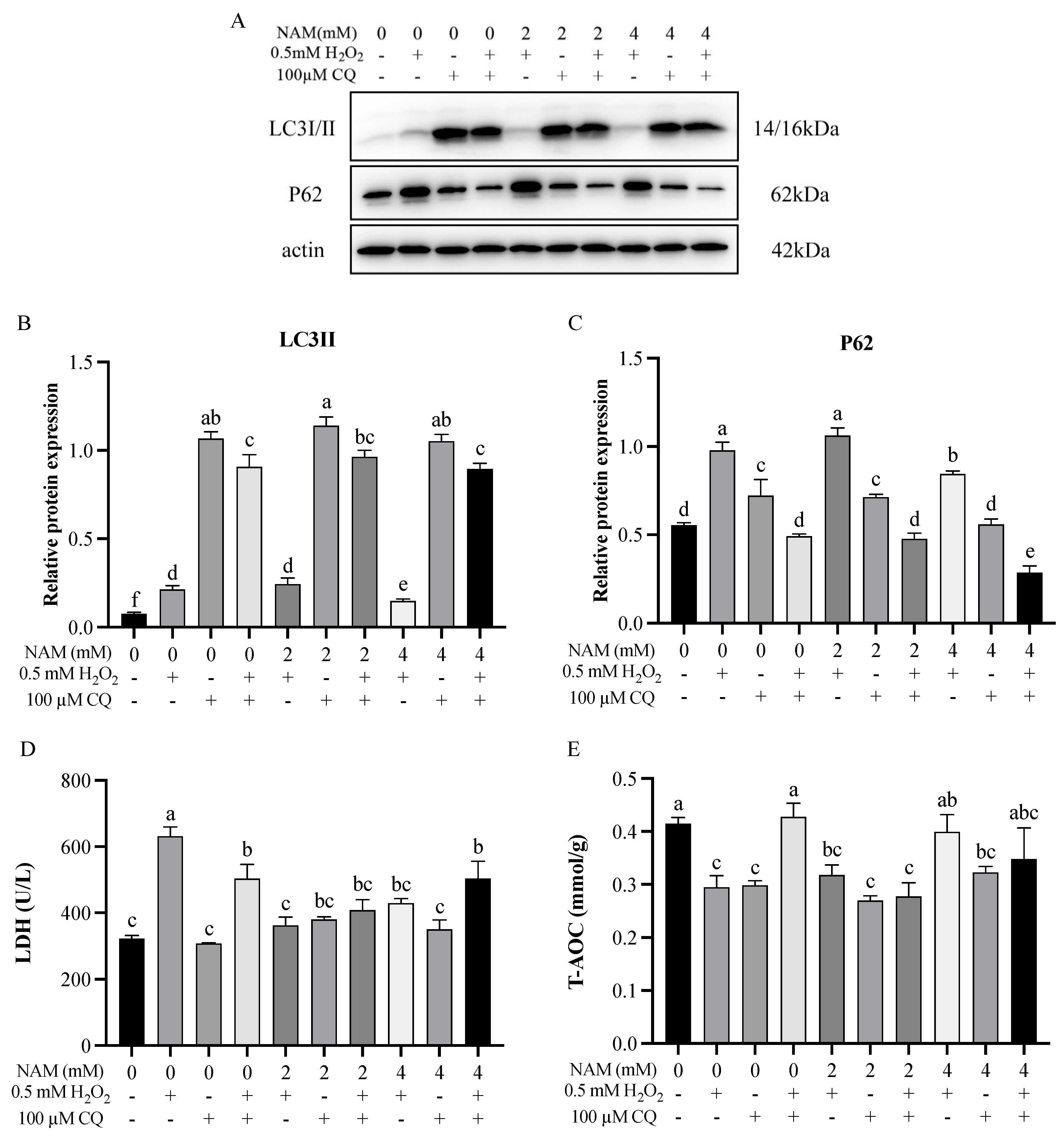
3.5. Effect of Autophagy Inhibitor and Nicotinamide on Bovine Intestinal Epithelial Cells Challenged by H2O2
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wei, X.; Li, D.; Feng, C.; Mao, H.; Zhu, J.; Cui, Y.; Yang, J.; Gao, H.; Wang, C. Effects of Hydrogen Peroxide and L-Tryptophan on Antioxidative Potential, Apoptosis, and Mammalian Target of Rapamycin Signaling in Bovine Intestinal Epithelial Cells. J. Dairy. Sci. 2022, 105, 10007–10019. [Google Scholar] [CrossRef]
- Lueschow, S.R.; McElroy, S.J. The Paneth Cell: The Curator and Defender of the Immature Small Intestine. Front. Immunol. 2020, 11, 587. [Google Scholar] [CrossRef] [PubMed]
- Peterson, L.W.; Artis, D. Intestinal Epithelial Cells: Regulators of Barrier Function and Immune Homeostasis. Nat. Rev. Immunol. 2014, 14, 141–153. [Google Scholar] [CrossRef] [PubMed]
- Mao, H.; Zhang, Y.; Ji, W.; Yun, Y.; Wei, X.; Cui, Y.; Wang, C. Leucine Protects Bovine Intestinal Epithelial Cells from Hydrogen Peroxide-Induced Apoptosis by Alleviating Oxidative Damage. J. Sci. Food Agric. 2022, 102, 5903–5912. [Google Scholar] [CrossRef]
- Sun, Z.; Liu, H.; Yang, Z.; Shao, D.; Zhang, W.; Ren, Y.; Sun, B.; Lin, J.; Xu, M.; Nie, S. Intestinal Trefoil Factor Activates the PI3K/Akt Signaling Pathway to Protect Gastric Mucosal Epithelium from Damage. Int. J. Oncol. 2014, 45, 1123–1132. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Sun, R.; Wang, G.; Chen, Z.; Li, Y.; Zhao, Y.; Liu, D.; Zhao, H.; Zhang, F.; Yao, J.; et al. SIRT3-Mediated Deacetylation of PRDX3 Alleviates Mitochondrial Oxidative Damage and Apoptosis Induced by Intestinal Ischemia/Reperfusion Injury. Redox Biol. 2020, 28, 101343. [Google Scholar] [CrossRef]
- Bogan, K.L.; Brenner, C. Nicotinic Acid, Nicotinamide, and Nicotinamide Riboside: A Molecular Evaluation of NAD+ Precursor Vitamins in Human Nutrition. Annu. Rev. Nutr. 2008, 28, 115–130. [Google Scholar] [CrossRef]
- Wei, X.; Yin, Q.; Zhao, H.; Cao, Y.; Cai, C.; Yao, J. Metabolomics for the Effect of Biotin and Nicotinamide on Transition Dairy Cows. J. Agric. Food Chem. 2018, 66, 5723–5732. [Google Scholar] [CrossRef]
- Song, S.B.; Jang, S.-Y.; Kang, H.T.; Wei, B.; Jeoun, U.-W.; Yoon, G.S.; Hwang, E.S. Modulation of Mitochondrial Membrane Potential and ROS Generation by Nicotinamide in a Manner Independent of SIRT1 and Mitophagy. Mol. Cells 2017, 40, 503–514. [Google Scholar] [CrossRef]
- Wei, X.; Yin, Q.; Zhao, H.; He, J.; Cao, Y.; Yao, J. Nicotinamide Supplementation during Postpartum and Peripartum Modulates Hepatic Energy and Lipid Metabolism, Oxidative Status, and Metabolomics Profile, as Well as Lipids in the Adipose Tissue of Goats. Anim. Feed. Sci. Technol. 2021, 274, 114849. [Google Scholar] [CrossRef]
- Chen, H.; Chen, K.; Zhao, F.; Guo, Y.; Liang, Y.; Wang, Z.; Liu, T.; Chen, S. Macroautophagy Involved in Testosterone Synthesis in Leydig Cells of Male Dairy Goat (Capra hircus). Theriogenology 2022, 180, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Wu, D.; Zhou, S.; Hu, H.; Li, F.; Guan, Z.; Zhan, X.; Gao, Y.; Wang, P.; Rao, Z. MLKL Deficiency Attenuated Hepatocyte Oxidative DNA Damage by Activating Mitophagy to Suppress Macrophage CGAS-STING Signaling during Liver Ischemia and Reperfusion Injury. Cell Death Discov. 2023, 9, 58. [Google Scholar] [CrossRef] [PubMed]
- Yue, K.; Pu, X.; Loor, J.J.; Jiang, Q.; Dong, J.; Shen, T.; Li, G.; Gao, W.; Lei, L.; Du, X.; et al. Impaired Autophagy Aggravates Oxidative Stress in Mammary Gland of Dairy Cows with Clinical Ketosis. J. Dairy. Sci. 2022, 105, 6030–6040. [Google Scholar] [CrossRef]
- Dammak, A.; Pastrana, C.; Martin-Gil, A.; Carpena-Torres, C.; Peral Cerda, A.; Simovart, M.; Alarma, P.; Huete-Toral, F.; Carracedo, G. Oxidative Stress in the Anterior Ocular Diseases: Diagnostic and Treatment. Biomedicines 2023, 11, 292. [Google Scholar] [CrossRef]
- Zhang, S.; Yang, G.; Guan, W.; Li, B.; Feng, X.; Fan, H. Autophagy Plays a Protective Role in Sodium Hydrosulfide-Induced Acute Lung Injury by Attenuating Oxidative Stress and Inflammation in Rats. Chem. Res. Toxicol. 2021, 34, 857–864. [Google Scholar] [CrossRef] [PubMed]
- Ji, Y.; He, Y.; Yang, Y.; Dai, Z.; Wu, Z. Hydroxyproline Alleviates 4-Hydroxy-2-Nonenal-Induced DNA Damage and Apoptosis in Porcine Intestinal Epithelial Cells. Anim. Nutr. 2022, 9, 7–15. [Google Scholar] [CrossRef]
- Mauthe, M.; Orhon, I.; Rocchi, C.; Zhou, X.; Luhr, M.; Hijlkema, K.-J.; Coppes, R.P.; Engedal, N.; Mari, M.; Reggiori, F. Chloroquine Inhibits Autophagic Flux by Decreasing Autophagosome-Lysosome Fusion. Autophagy 2018, 14, 1435–1455. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Chang, R.; Tang, Y.; Luo, S.; Jiang, C.; Jia, H.; Xu, Q.; Dong, Z.; Liang, Y.; Loor, J.J.; et al. Transcription Factor EB (TFEB)-Mediated Autophagy Protects Bovine Mammary Epithelial Cells against H2O2-Induced Oxidative Damage in Vitro. J. Anim. Sci. Biotechnol. 2021, 12, 35. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, Y.; Zhang, X.; Lu, Y.; Chen, H. New Insights in Intestinal Oxidative Stress Damage and the Health Intervention Effects of Nutrients: A Review. J. Funct. Foods 2020, 75, 104248. [Google Scholar] [CrossRef]
- Cheng, W.-H. Green Tea: An Ancient Antioxidant Drink for Optimal Health? J. Nutr. 2019, 149, 1877–1879. [Google Scholar] [CrossRef]
- Mitchell, S.J.; Bernier, M.; Aon, M.A.; Cortassa, S.; Kim, E.Y.; Fang, E.F.; Palacios, H.H.; Ali, A.; Navas-Enamorado, I.; Di Francesco, A.; et al. Nicotinamide Improves Aspects of Healthspan, but Not Lifespan, in Mice. Cell Metab. 2018, 27, 667–676.e4. [Google Scholar] [CrossRef] [PubMed]
- Nunes, C.; Freitas, V.; Almeida, L.; Laranjinha, J. Red Wine Extract Preserves Tight Junctions in Intestinal Epithelial Cells under Inflammatory Conditions: Implications for Intestinal Inflammation. Food Funct. 2019, 10, 1364–1374. [Google Scholar] [CrossRef] [PubMed]
- Kuo, W.-T.; Odenwald, M.A.; Turner, J.R.; Zuo, L. Tight Junction Proteins Occludin and ZO-1 as Regulators of Epithelial Proliferation and Survival. Ann. N. Y. Acad. Sci. 2022, 1514, 21–33. [Google Scholar] [CrossRef] [PubMed]
- Weiss, G.A.; Hennet, T. Mechanisms and Consequences of Intestinal Dysbiosis. Cell Mol. Life Sci. 2017, 74, 2959–2977. [Google Scholar] [CrossRef] [PubMed]
- Yi, Z.; Tan, X.; Wang, Q.; Huang, P.; Li, Y.; Ding, X.; Li, J.; Huang, J.; Yang, H.; Yin, Y. Dietary Niacin Affects Intestinal Morphology and Functions via Modulating Cell Proliferation in Weaned Piglets. Food Funct. 2021, 12, 7402–7414. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Zhou, Y.; Pang, N.; Hu, Q.; Li, Q.; Sun, Y.; Ding, Y.; Gu, Y.; Xiao, Y.; Gao, M.; et al. NAD Supplement Alleviates Intestinal Barrier Injury Induced by Ethanol Via Protecting Epithelial Mitochondrial Function. Nutrients 2022, 15, 174. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Wang, H.; Fang, S.; Xu, C. Roles of Endoplasmic Reticulum Stress and Autophagy on H2O2-induced Oxidative Stress Injury in HepG2 Cells. Mol. Med. Rep. 2018, 18, 4163–4174. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Li, L.; Wang, S.; Yu, C.; Xiao, B.; Lin, L.; Cong, W.; Cheng, J.; Yang, W.; Sun, W.; et al. H2O2 Treatment or Serum Deprivation Induces Autophagy and Apoptosis in Naked Mole-Rat Skin Fibroblasts by Inhibiting the PI3K/Akt Signaling Pathway. Oncotarget 2016, 7, 84839–84850. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wang, C.; Zhu, J.; Lin, Q.; Yu, M.; Wen, J.; Feng, J.; Hu, C. Sodium Butyrate Ameliorates Oxidative Stress-Induced Intestinal Epithelium Barrier Injury and Mitochondrial Damage through AMPK-Mitophagy Pathway. Oxid. Med. Cell Longev. 2022, 2022, 3745135. [Google Scholar] [CrossRef]
- Komatsu, M.; Kurokawa, H.; Waguri, S.; Taguchi, K.; Kobayashi, A.; Ichimura, Y.; Sou, Y.-S.; Ueno, I.; Sakamoto, A.; Tong, K.I.; et al. The Selective Autophagy Substrate P62 Activates the Stress Responsive Transcription Factor Nrf2 through Inactivation of Keap1. Nat. Cell Biol. 2010, 12, 213–223. [Google Scholar] [CrossRef]
- Shvets, E.; Fass, E.; Scherz-Shouval, R.; Elazar, Z. Correction: The N-Terminus and Phe52 Residue of LC3 Recruit P62/SQSTM1 into Autophagosomes. J. Cell Sci. 2022, 135, jcs260853. [Google Scholar] [CrossRef] [PubMed]
- Desagher, S.; Martinou, J.-C. Solange Desagher and Jean-Claude Martinou: Executioners of Cell Death. Trends Cell Biol. 2016, 26, 560–562. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.; Hussien, R.; Brooks, G.A. H2O2-Induced Mitochondrial Fragmentation in C2C12 Myocytes. Free Radic. Biol. Med. 2010, 49, 1646–1654. [Google Scholar] [CrossRef] [PubMed]
- Cao, S.; Wang, C.; Yan, J.; Li, X.; Wen, J.; Hu, C. Curcumin Ameliorates Oxidative Stress-Induced Intestinal Barrier Injury and Mitochondrial Damage by Promoting Parkin Dependent Mitophagy through AMPK-TFEB Signal Pathway. Free Radic. Biol. Med. 2020, 147, 8–22. [Google Scholar] [CrossRef] [PubMed]
- Wan, J.; Weiss, E.; Ben Mkaddem, S.; Mabire, M.; Choinier, P.-M.; Picq, O.; Thibault-Sogorb, T.; Hegde, P.; Pishvaie, D.; Bens, M.; et al. LC3-Associated Phagocytosis Protects against Inflammation and Liver Fibrosis via Immunoreceptor Inhibitory Signaling. Sci. Transl. Med. 2020, 12, eaaw8523. [Google Scholar] [CrossRef] [PubMed]
- Cocco, S.; Leone, A.; Roca, M.S.; Lombardi, R.; Piezzo, M.; Caputo, R.; Ciardiello, C.; Costantini, S.; Bruzzese, F.; Sisalli, M.J.; et al. Inhibition of Autophagy by Chloroquine Prevents Resistance to PI3K/AKT Inhibitors and Potentiates Their Antitumor Effect in Combination with Paclitaxel in Triple Negative Breast Cancer Models. J. Transl. Med. 2022, 20, 290. [Google Scholar] [CrossRef] [PubMed]
- Bik, E.; Mateuszuk, L.; Orleanska, J.; Baranska, M.; Chlopicki, S.; Majzner, K. Chloroquine-Induced Accumulation of Autophagosomes and Lipids in the Endothelium. Int. J. Mol. Sci. 2021, 22, 2401. [Google Scholar] [CrossRef]
- Javaid, H.M.A.; Lim, H.; Shin, S.; Huh, J.Y. Inhibition of Autophagy with Chloroquine Dysregulates Mitochondrial Quality Control and Energetics in Adipocytes. Arch. Pharm. Res. 2022, 45, 731–742. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene 1 | Primer Sequences (5′-3′) |
|---|---|
| β-actin | F: CGGGAAATCGTCCGTGAC R: CCGTGTTGGCGTAGAGGT |
| CAT | F: AGATACTCCAAGGCGAAGGTG R: AAAGCCACGAGGGTCACGAAC |
| GPX1 | F: AGTGCGAGGTGARATGGCGAGARA R: TGGGCARAARATCCCTGGAGAGCA |
| SOD2 | F: CGTCGCCGAGGAGAAGTA R: CCAGCAGGGGGATAAGA |
| Claudin-2 | F: CCAGGCCATGATGGTGACAT R: GAAGAAGACTCCGCCCACAA |
| Occludin | F: ACGCAGGAAGTGCCTTTGGTAGC R: GCAGCCATGGCCAGCAGGAA |
| ZO-1 | F: GAAAGATGTTTATCGTCGCATCGT R: ATTCCTTCTCATATTCAAAATGGGTTCTGA |
| Items 1 | Treatments 2 | SEM | p-Value | |||
|---|---|---|---|---|---|---|
| CON | OS | OS2N | OS4N | |||
| Antioxidant enzymes | ||||||
| CAT | 1.01 a | 0.72 b | 0.87 ab | 1.09 a | 0.10 | 0.023 |
| GPX1 | 1.00 | 1.20 | 1.22 | 1.11 | 0.14 | 0.436 |
| SOD2 | 1.01 | 0.72 | 1.16 | 0.97 | 0.17 | 0.087 |
| Tight junction proteins | ||||||
| Occludin | 1.00 | 0.76 | 0.93 | 0.94 | 0.05 | 0.258 |
| Claudin-2 | 1.00 a | 0.62 b | 1.12 a | 0.96 a | 0.08 | 0.015 |
| ZO-1 | 1.00 | 1.07 | 0.83 | 0.99 | 0.05 | 0.119 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, Y.; Feng, C.; Zhang, Y.; Hu, K.; Wang, C.; Wei, X. Nicotinamide Supplementation Mitigates Oxidative Injury of Bovine Intestinal Epithelial Cells through Autophagy Modulation. Animals 2024, 14, 1483. https://doi.org/10.3390/ani14101483
Guo Y, Feng C, Zhang Y, Hu K, Wang C, Wei X. Nicotinamide Supplementation Mitigates Oxidative Injury of Bovine Intestinal Epithelial Cells through Autophagy Modulation. Animals. 2024; 14(10):1483. https://doi.org/10.3390/ani14101483
Chicago/Turabian StyleGuo, Yihan, Changdong Feng, Yiwei Zhang, Kewei Hu, Chong Wang, and Xiaoshi Wei. 2024. "Nicotinamide Supplementation Mitigates Oxidative Injury of Bovine Intestinal Epithelial Cells through Autophagy Modulation" Animals 14, no. 10: 1483. https://doi.org/10.3390/ani14101483