
Tackling Antibiotic Resistance: Exploring 5-Fluorouracil as a Promising Antimicrobial Strategy for the Treatment of Streptococcus suis Infection
 ,
,
Abstract
:Simple Summary
Abstract

1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
2.2. Construction of thyA Deletion Mutant Strain of S. suis
2.3. Cytotoxicity Assay
2.4. Resistance-Development Studies
2.5. Determination of Minimum Inhibitory Concentration (MIC) and Minimum Bactericide Concentration (MBC)
2.6. Bacterial Growth Curves
2.7. Colony-Forming Unit Assay
2.8. Scanning Electron Microscope (SEM)
2.9. Transmission Electron Microscope (TEM)
2.10. Lactate Dehydrogenase (LDH) Assay
2.11. Determination of Genomic DNA Integrity
2.12. RNA Extraction and Quantitative Reverse Transcription Polymerase Chain Reaction (qRT-PCR)
2.13. Animal Experiments
2.14. Statistical Analysis
3. Results
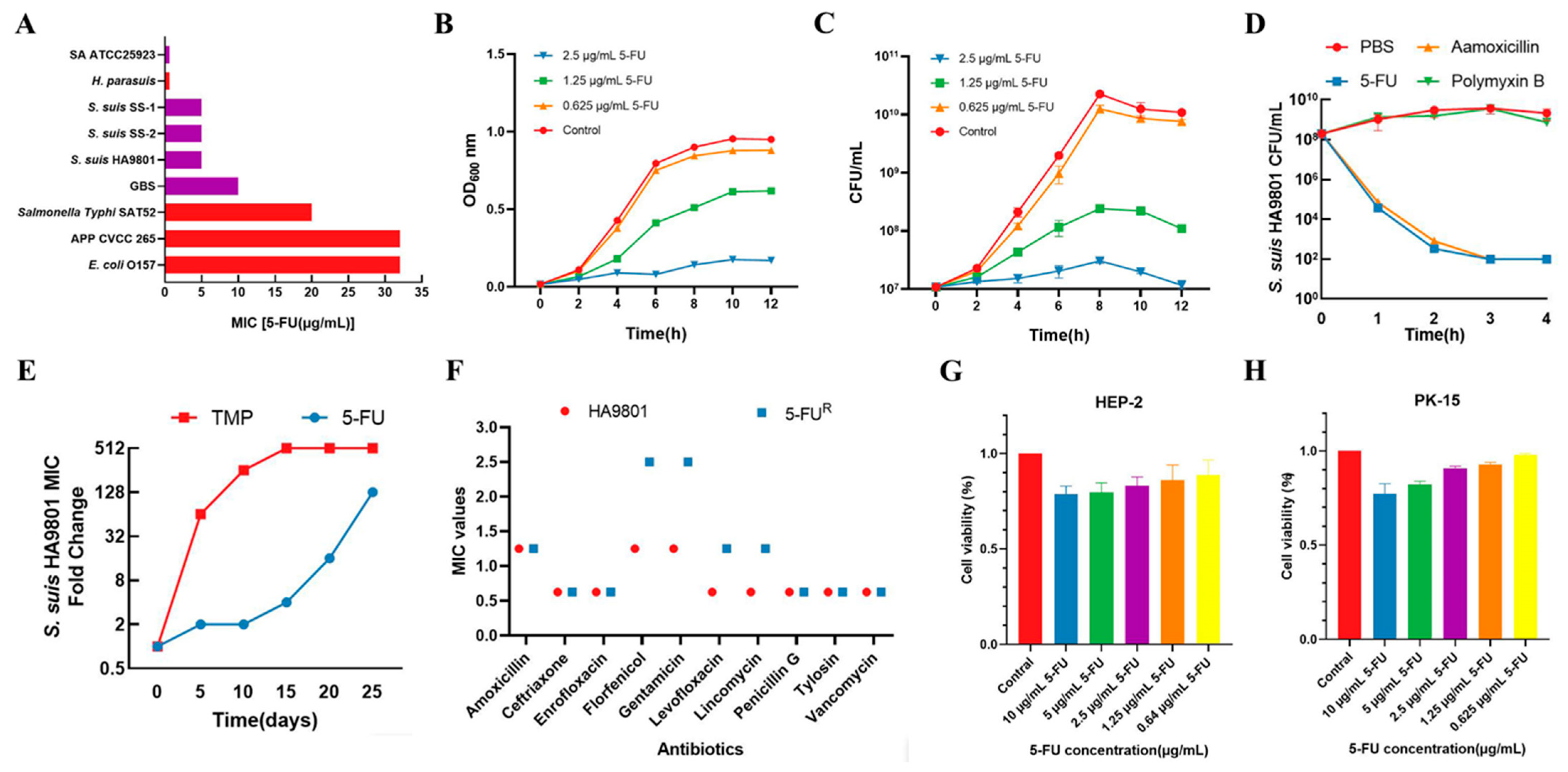
3.1. 5-FU Exhibits Effective Antibacterial Activity and Safety In Vitro
3.2. 5-FU Disrupts the Cell Wall and Bacterial Membrane Morphology and Induces DNA Damage in S. suis
3.3. 5-FU Causes Damage to S. suis by Inhibiting Thymidine Synthesis
3.4. 5-FU Reduces S. suis Colonization of Tissues in Mice
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gottschalk, M.; Xu, J.; Calzas, C.; Segura, M. Streptococcus suis: A new emerging or an old neglected zoonotic pathogen? Future Microbiol. 2010, 5, 371–391. [Google Scholar] [CrossRef]
- Wang, J.; Liang, P.; Sun, H.; Wu, Z.; Gottschalk, M.; Qi, K.; Zheng, H. Comparative transcriptomic analysis reveal genes involved in the pathogenicity increase of Streptococcus suis epidemic strains. Virulence 2022, 13, 1455–1470. [Google Scholar] [CrossRef]
- Zhong, X.; Ma, J.; Bai, Q.; Zhu, Y.; Zhang, Y.; Gu, Q.; Pan, Z.; Liu, G.; Wu, Z.; Yao, H. Identification of the RNA-binding domain-containing protein RbpA that acts as a global regulator of the pathogenicity of Streptococcus suis serotype 2. Virulence 2022, 13, 1304–1314. [Google Scholar] [CrossRef]
- Feng, Y.; Zhang, H.; Ma, Y.; Gao, G.F. Uncovering newly emerging variants of Streptococcus suis, an important zoonotic agent. Trends Microbiol. 2010, 18, 124–131. [Google Scholar] [CrossRef]
- Seitz, M.; Valentin-Weigand, P.; Willenborg, J. Use of Antibiotics and Antimicrobial Resistance in Veterinary Medicine as Exemplified by the Swine Pathogen Streptococcus suis. Curr. Top. Microbiol. Immunol. 2016, 398, 103–121. [Google Scholar]
- Tan, C.; Zhang, A.; Chen, H.; Zhou, R. Recent Proceedings on Prevalence and Pathogenesis of Streptococcus suis. Curr. Issues Mol. Biol. 2019, 32, 473–520. [Google Scholar] [CrossRef]
- Theuretzbacher, U.; Gottwalt, S.; Beyer, P.; Butler, M.; Czaplewski, L.; Lienhardt, C.; Moja, L.; Paul, M.; Paulin, S.; Rex, J.H.; et al. Analysis of the clinical antibacterial and antituberculosis pipeline. Lancet Infect. Dis. 2019, 19, e40–e50. [Google Scholar] [CrossRef]
- Årdal, C.; Balasegaram, M.; Laxminarayan, R.; McAdams, D.; Outterson, K.; Rex, J.H.; Sumpradit, N. Antibiotic development—economic, regulatory and societal challenges. Nat. Rev. Microbiol. 2019, 18, 267–274. [Google Scholar] [CrossRef]
- Kinnings, S.L.; Liu, N.; Buchmeier, N.; Tonge, P.J.; Xie, L.; Bourne, P.E. Drug discovery using chemical systems biology: Repositioning the safe medicine Comtan to treat multi-drug and extensively drug resistant tuberculosis. PLoS Comput. Biol. 2009, 5, e1000423. [Google Scholar] [CrossRef]
- Wilson, P.M.; Danenberg, P.V.; Johnston, P.G.; Lenz, H.-J.; Ladner, R.D. Standing the test of time: Targeting thymidylate biosynthesis in cancer therapy. Nat. Rev. Clin. Oncol. 2014, 11, 282–298. [Google Scholar] [CrossRef]
- Longley, D.B.; Harkin, D.P.; Johnston, P.G. 5-Fluorouracil: Mechanisms of action and clinical strategies. Nat. Rev. Cancer 2003, 3, 330–338. [Google Scholar] [CrossRef]
- Cohen, S.S.; Flaks, J.G.; Barner, H.D.; Loeb, M.R.; Lichtenstein, J. The Mode of Action of 5-Fluorouracil and Its Derivatives. Proc. Natl. Acad. Sci. USA 1958, 44, 1004–1012. [Google Scholar] [CrossRef]
- Sedlmayer, F.; Woischnig, A.K.; Unterreiner, V.; Fuchs, F.; Baeschlin, D.; Khanna, N.; Fussenegger, M. 5-Fluorouracil blocks quorum-sensing of biofilm-embedded methicillin-resistant Staphylococcus aureus in mice. Nucleic Acids Res. 2021, 49, e73. [Google Scholar] [CrossRef]
- Di Bonaventura, G.; Lupetti, V.; Di Giulio, A.; Muzzi, M.; Piccirilli, A.; Cariani, L.; Pompilio, A. Repurposing High-Throughput Screening Identifies Unconventional Drugs with Antibacterial and Antibiofilm Activities against Pseudomonas aeruginosa under Experimental Conditions Relevant to Cystic Fibrosis. Microbiol. Spectr. 2023, 11, e0035223. [Google Scholar] [CrossRef]
- Oe, C.; Hayashi, H.; Hirata, K.; Kawaji, K.; Hashima, F.; Sasano, M.; Furuichi, M.; Usui, E.; Katsumi, M.; Suzuki, Y.; et al. Pyrimidine Analogues as a New Class of Gram-Positive Antibiotics, Mainly Targeting Thymineless-Death Related Proteins. ACS Infect. Dis. 2020, 6, 1490–1500. [Google Scholar] [CrossRef]
- Bean, B.; Tomasz, A. Inhibitory effects and metabolism of 5-fluoropyrimidine derivatives in pneumococcus. J. Bacteriol. 1971, 106, 412–420. [Google Scholar] [CrossRef]
- Ueda, A.; Attila, C.; Whiteley, M.; Wood, T.K. Uracil influences quorum sensing and biofilm formation in Pseudomonas aeruginosa and fluorouracil is an antagonist. Microb. Biotechnol. 2009, 2, 62–74. [Google Scholar] [CrossRef]
- Singh, V.; Brecik, M.; Mukherjee, R.; Evans, J.C.; Svetlíková, Z.; Blaško, J.; Surade, S.; Blackburn, J.; Warner, D.F.; Mikušová, K.; et al. The Complex Mechanism of Antimycobacterial Action of 5-Fluorouracil. Chem. Biol. 2015, 22, 63–75. [Google Scholar] [CrossRef]
- Wang, Y.; Gong, S.; Dong, X.; Li, J.; Grenier, D.; Yi, L. In vitro Mixed Biofilm of Streptococcus suis and Actinobacillus pleuropneumoniae Impacts Antibiotic Susceptibility and Modulates Virulence Factor Gene Expression. Front. Microbiol. 2020, 11, 507. [Google Scholar] [CrossRef]
- Gao, M.X.; Zuo, J.; Shen, Y.M.; Yuan, S.; Gao, S.J.; Wang, Y.X.; Wang, Y.; Yi, L. Modeling Co-Infection by and Reveals Influences on Biofilm Formation and Host Response. Animals 2023, 13, 1511. [Google Scholar] [CrossRef]
- Peng, M.; Xu, Y.; Dou, B.; Yang, F.; He, Q.; Liu, Z.; Gao, T.; Liu, W.; Yang, K.; Guo, R.; et al. The adcA and lmb Genes Play an Important Role in Drug Resistance and Full Virulence of Streptococcus suis. Microbiol. Spectr. 2023, 11, e0433722. [Google Scholar] [CrossRef]
- Takamatsu, D.; Osaki, M.; Sekizaki, T. Thermosensitive suicide vectors for gene replacement in Streptococcus suis. Plasmid 2001, 46, 140–148. [Google Scholar] [CrossRef]
- Koeth, L.M.; DiFranco-Fisher, J.M.; McCurdy, S. A Reference Broth Microdilution Method for Dalbavancin In Vitro Susceptibility Testing of Bacteria that Grow Aerobically. J. Vis. Exp. 2015, 103, 53028. [Google Scholar]
- Li, J.; Fan, Q.; Jin, M.; Mao, C.; Zhang, H.; Zhang, X.; Sun, L.; Grenier, D.; Yi, L.; Hou, X.; et al. Paeoniflorin reduce luxS/AI-2 system-controlled biofilm formation and virulence in Streptococcus suis. Virulence 2021, 12, 3062–3073. [Google Scholar] [CrossRef]
- Rasool, K.; Helal, M.; Ali, A.; Ren, C.E.; Gogotsi, Y.; Mahmoud, K.A. Antibacterial Activity of Ti3C2Tx MXene. ACS Nano 2016, 10, 3674–3684. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, Y.; Li, J.; Gong, S.; Sun, L.; Grenier, D.; Li, Y. Pdh is involved in the cell division and Normal septation of Streptococcus suis. Microbiol. Res. 2019, 228, 126304. [Google Scholar] [CrossRef]
- Xiong, Z.; Zhang, X.; Zhang, S.; Lei, L.; Ma, W.; Li, D.; Wang, W.; Zhao, Q.; Xing, B. Bacterial toxicity of exfoliated black phosphorus nanosheets. Ecotoxicol. Environ. Saf. 2018, 161, 507–514. [Google Scholar] [CrossRef]
- Xu, Z.; Qiu, Z.; Liu, Q.; Huang, Y.; Li, D.; Shen, X.; Fan, K.; Xi, J.; Gu, Y.; Tang, Y.; et al. Converting organosulfur compounds to inorganic polysulfides against resistant bacterial infections. Nat. Commun. 2018, 9, 3713. [Google Scholar] [CrossRef]
- Li, J.; Fan, Q.; Zuo, J.; Xue, B.; Zhang, X.; Wei, Y.; Sun, L.; Grenier, D.; Yi, L.; Hou, X.; et al. Paeoniflorin combined with norfloxacin ameliorates drug-resistant Streptococcus suis infection. J. Antimicrob. Chemother. 2022, 77, 3275–3282. [Google Scholar] [CrossRef]
- Liao, Z.Y.; Sordet, O.; Zhang, H.L.; Kohlhagen, G.; Antony, S.; Gmeiner, W.H.; Pommier, Y. A novel polypyrimidine antitumor agent FdUMP[10] induces thymineless death with topoisomerase I-DNA complexes. Cancer Res. 2005, 65, 4844–4851. [Google Scholar] [CrossRef]
- Boshoff, H.I.; Reed, M.B.; Barry, C.E., 3rd; Mizrahi, V. DnaE2 polymerase contributes to in vivo survival and the emergence of drug resistance in Mycobacterium tuberculosis. Cell 2003, 113, 183–193. [Google Scholar] [CrossRef]
- Humeniuk, R.; Menon, L.G.; Mishra, P.J.; Gorlick, R.; Sowers, R.; Rode, W.; Pizzorno, G.; Cheng, Y.C.; Kemeny, N.; Bertino, J.R.; et al. Decreased levels of UMP kinase as a mechanism of fluoropyrimidine resistance. Mol. Cancer Ther. 2009, 8, 1037–1044. [Google Scholar] [CrossRef]
- Santi, D.V.; McHenry, C.S.; Sommer, H. Mechanism of interaction of thymidylate synthetase with 5-fluorodeoxyuridylate. Biochemistry 1974, 13, 471–481. [Google Scholar] [CrossRef]
- Curtin, N.J.; Harris, A.L.; Aherne, G.W. Mechanism of cell death following thymidylate synthase inhibition: 2′-deoxyuridine-5′-triphosphate accumulation, DNA damage, and growth inhibition following exposure to CB3717 and dipyridamole. Cancer Res. 1991, 51, 2346–2352. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate | Strain/ Subspecies | MIC 5-FU (µg/mL) | MBC 5-FU (µg/mL) | MIC AMX (µg/mL) | Temperature (°C) | Media a |
|---|---|---|---|---|---|---|
| Actinobacillus pleuropneumoniae | CVCC 265 | 32 | >128 | 37 | TSB Broth + NAD | |
| Escherichia coli | O157:H7 | 32 | >128 | 37 | LB Broth | |
| Glaesserella parasuis | [20] | 0.625 | 5 | 37 | TSB Broth + serum + NAD | |
| GBS | [19] | 10 | 40 | 37 | TSB Broth | |
| Salmonella Typhimurium | SAT52 | 20 | 40 | 37 | LB Broth | |
| Staphylococcus aureus | ATCC25923 | 0.625 | 10 | 37 | TSB Broth | |
| Streptococcus suis | HA9801 | 5 | 10 | 1.25 | 37 | TSB Broth |
| Streptococcus suis | HA9801 | 40 | 160 | 1.25 | 37 | TSB Broth + Thymine |
| Streptococcus suis | HA9801 ΔthyA | 80 | 320 | 1.25 | 37 | TSB Broth + Thymine |
| Streptococcus suis | SS-1 | 5 | 10 | 2.5 | 37 | TSB Broth |
| Streptococcus suis | SS-2 | 5 | 10 | 2.5 | 37 | TSB Broth |
| Streptococcus suis | SS-1 | 20 | 80 | 2.5 | 37 | TSB Broth + Thymine |
| Streptococcus suis | SS-2 | 40 | 160 | 2.5 | 37 | TSB Broth + Thymine |
| Plasmids and Primers | Relevant Characteristics a or Sequence (5′-3′) b | Size (bp) | References or Target Gene |
|---|---|---|---|
| Plasmids | |||
| pSET4s | Thermosensitive suicide vector; SpcR | [22] | |
| pSET4s-ΔthyA | Knockout vector for thyA deletion; SpcR | This study | |
| Primers | |||
| gdh-F | GTTGAGCCTGAGCGTATCATC | 425 | An internal region of gdh |
| gdh-R | CCAGTCAAGACACCTGCATC | ||
| cps2-F | ATTGGTAGGCACTGTCGTTGGTC | 191 | An internal region of cps2 |
| cps2-R | AGAACTTAGCATTGTTGCGGTGG | ||
| thyA-A | CCGCGTCGACCATCACTCGTCTTGAAATAATCGTT | 792 | The left arm of thyA |
| thyA-B | GGTCTTAGTATAGCAAATTCTAGCA | ||
| thyA-C | TGCTAGAATTTGCTATACTAAGACCCTGGATTAGCAGTGAAGAACTTCGT | 760 | The right arm of thyA |
| thyA-D | ACGCGGATCCCATACGGGTTTTTCTCGTCTTTTGG | ||
| thyA-ORF-S | GTCTAAGGGCGAGTTTCC | 554 | An internal region of thyA |
| thyA-ORF-A | GACGGAGTAATTCTTCAGC | ||
| thyA-XY-S | AGTCCTCAATCCTGCCTACATCGTT | 1636 | A fragment containing thyA |
| thyA-XY-A | TCGCCATTAAATTGACC | ||
| Q-16S rRNA-F | GTTGCGAACGGGTGAGTAA | An internal region of 16S rRNA | |
| Q-16S rRNA-R | TCTCAGGTCGGCTATGTATC G | ||
| Q-recA-F | TCCCCTGAGTCTGGCTGTG | An internal region of recA | |
| Q-recA-R | ATGGTGGTATTGCTGCCTTTA | ||
| Q-radA-F | AACATAGTCGGCGTCACCA | An internal region of radA | |
| Q-radA-R | AAAATCGCTTCGGCTCCAC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zuo, J.; Quan, Y.; Li, J.; Li, Y.; Song, D.; Li, X.; Wang, Y.; Yi, L.; Wang, Y. Tackling Antibiotic Resistance: Exploring 5-Fluorouracil as a Promising Antimicrobial Strategy for the Treatment of Streptococcus suis Infection. Animals 2024, 14, 1286. https://doi.org/10.3390/ani14091286
Zuo J, Quan Y, Li J, Li Y, Song D, Li X, Wang Y, Yi L, Wang Y. Tackling Antibiotic Resistance: Exploring 5-Fluorouracil as a Promising Antimicrobial Strategy for the Treatment of Streptococcus suis Infection. Animals. 2024; 14(9):1286. https://doi.org/10.3390/ani14091286
Chicago/Turabian StyleZuo, Jing, Yingying Quan, Jinpeng Li, Yue Li, Dong Song, Xingping Li, Yuxin Wang, Li Yi, and Yang Wang. 2024. "Tackling Antibiotic Resistance: Exploring 5-Fluorouracil as a Promising Antimicrobial Strategy for the Treatment of Streptococcus suis Infection" Animals 14, no. 9: 1286. https://doi.org/10.3390/ani14091286