

Protective Effects of Melatonin against Carcinogen-Induced Oxidative Damage in the Thyroid

1
Department of Endocrinology and Metabolic Diseases, Medical University of Lodz, Rzgowska St. 281/289, 93-338 Lodz, Poland
2
Polish Mother’s Memorial Hospital-Research Institute, Rzgowska St. 281/289, 93-338 Lodz, Poland
*
Author to whom correspondence should be addressed.
Cancers 2024, 16(9), 1646; https://doi.org/10.3390/cancers16091646
Submission received: 26 March 2024
/
Revised: 22 April 2024
/
Accepted: 23 April 2024
/
Published: 25 April 2024
(This article belongs to the Special Issue Melatonin and Cancer: Current Challenges and Future Perspectives)
Abstract
:Simple Summary
Melatonin is a hormone primarily produced in the pineal gland in response to darkness. Apart from regulating our body’s internal clock, it possesses strong antioxidant properties. Thanks to these characteristics, melatonin shows promise in preventing and treating various health issues, such as cancer. This review explores how melatonin can protect the thyroid gland from oxidative damage and, consequently, prevent the development of thyroid disease caused by documented or potential carcinogens. The article emphasizes the importance of melatonin in protecting the thyroid and how it can help to treat thyroid diseases, including cancer.
Abstract
Melatonin, primarily synthesized in the pineal gland, plays a crucial role in regulating circadian rhythms and possesses significant antioxidative properties. By neutralizing free radicals and reducing oxidative stress, melatonin emerges as a promising agent for the prevention and therapy of many different disorders, including cancer. This paper reviews the relationship between the thyroid gland and melatonin, presenting experimental evidence on the protective effects of this indoleamine against oxidative damage to macromolecules in thyroid tissue caused by documented carcinogens (as classified by the International Agency for Research on Cancer, IARC) or caused by potential carcinogens. Furthermore, the possible influence on cancer therapy in humans and the overall well-being of cancer patients are discussed. The article highlights melatonin’s essential role in maintaining thyroid health and its contribution to management strategies in patients with thyroid cancer and other thyroid diseases.
1. The Thyroid Gland as an Organ of Oxidative Nature
The thyroid gland is a crucial endocrine organ responsible for controlling fundamental physiological processes such as metabolism, tissue/organ growth, and development in all vertebrates [1]. It contains two types of hormone-producing cells: (1) the major type, called thyroid follicular cells (or thyrocytes), which produce mainly thyroxine (T4) and, to a lesser extent, triiodothyronine (T3), and (2) the other type, called parafollicular cells (or C cells), in which calcitonin is synthesized.
The primary secretory product of thyrocytes, T4, is also called a prohormone; it requires conversion to the active T3 in peripheral tissues. T3 exerts effects on virtually all human cells possessing a nucleus and plays a crucial role in increasing the overall metabolic rate. Both T3 and T4 are iodothyronines, i.e., products of iodine incorporation into tyrosine residues within thyroglobulin (Tg).
Although the process of thyroid hormone synthesis is crucial for the entire organism, it can pose risks to the thyroid gland itself under special circumstances. This is because it involves a complex and multistep series of redox reactions utilizing hydrogen peroxide (H2O2), as the primary oxidizing agent, which is the basic form of reactive oxygen species (ROS). H2O2 acts as an oxidative equivalent in the following reactions catalyzed by thyroid peroxidase (thyroperoxidase; TPO): iodide oxidation, iodination of tyrosyl residues in Tg, and the subsequent phenoxy ether bond formation between pairs of iodotyrosines to generate iodothyronines. Because H2O2 is indispensable in the process of thyroid hormone formation and, therefore, should be available in unlimited quantities, it is probably present in the thyroid in amounts exceeding those required for iodine incorporation into thyroid hormones [2]. The fact that normal thyroid function depends on ROS and, therefore, this organ is constantly exposed to them, places the thyroid gland at risk of high oxidative stress. This phenomenon may justify the statement that the thyroid gland is an organ of “oxidative nature” [3] [Figure 1].
H2O2 is produced in the thyroid gland by NADPH oxidase/dual oxidase (NOX/DUOX), which are family transmembrane proteins. The thyroidal NOX/DUOX group comprises three enzymes: i.e., dual oxidase 1 (DUOX1), dual oxidase 2 (DUOX2) and NADPH oxidase 4 (NOX4). Of importance, DUOX1 and DUOX2 are typically active only at the apical plasma membrane of thyroid follicular cells [4]. Whereas DUOX2 is the main NOX/DUOX isoform catalyzing H2O2 formation for thyroid hormone synthesis [5,6], DUOX1 is probably active only under certain conditions associated with DUOX2 unavailability; however, its role in thyroid physiology is still not clear [7].
Contrary to DUOX enzymes, NOX4 is distinguished by its intracellular localization and, notably, by its continuous uncontrolled H2O2 production, which results solely from the gene expression of this enzyme [8]. The physiological function of H2O2 produced by NOX4 in the normal thyroid gland is not fully known. However, it is implied that NOX4 plays an important role in the regulation of redox processes occurring close to the basal cellular membrane. Studies indicate that H2O2 produced by NOX4 is potentially involved in the redox-sensitive control of thyroid differentiation [9,10]. It has been shown that TSH represses and iodine excess increases NOX4 mRNA expression [10]. NOX4-derived H2O2 also mediates endoplasmic reticulum signaling [11]. Therefore, taking into account the large amounts of secretory proteins required for thyroid functioning synthesized by the endoplasmic reticulum, the function of NOX4 may be of crucial importance [12].
Uncontrolled H2O2 production by NOX4 gains special significance when we consider that H2O2, in addition to serving as a substrate in hormone synthesis and signal transduction, can become a significant source of free radicals and other ROS. Because it is an oxidizing agent, H2O2 can induce damage to biological macromolecules such as DNA, lipids, and proteins. Additionally, H2O2 is a compound with a relatively long half-life (milliseconds to seconds) [13] and as a non-polar molecule has the capacity to diffuse through biological membranes, creating a hazard in a place far from its origin. Although H2O2 itself has limited reactivity towards macromolecules, it can give rise to highly reactive and most harmful free radical, i.e., hydroxyl radical (•OH), in a process that is catalyzed by transition metal ions, typically ferrous ion (Fe2+), which is known as the Fenton reaction (Fe2+ + H2O2 → Fe3+ + •OH + OH−). This reaction is particularly important in the thyroid gland, where heme iron is a component of TPO, a typical heme-containing enzyme. Therefore, iron is necessary for TPO biological activity and, consequently, for thyroid hormone synthesis.
Both experimental and epidemiological studies have shown that iron deficiency can negatively impact thyroid function [14]. It is known that thyroid hormone concentrations are lower in patients with iron deficiency, especially in certain patient groups such as pregnant women [15]. While the level of iron in the thyroid gland is tightly regulated, the fact that activated TPO is located at the apical membrane, exposing its heme-linked catalytic site to the thyroid follicular lumen, may result in an excessive amount of iron, which could potentially increase the risk of adverse oxidative reactions.
H2O2, through various mechanisms, may contribute to the formation of thyroid cancer [16]. Importantly, H2O2 may be involved in the co-occurrence of thyroid cancer and congenital hypothyroidism [17].
It is worth mentioning that in numerous experimental studies, Fenton reaction substrates, i.e., Fe2+ and H2O2, are repeatedly confirmed to increase oxidative damage to macromolecules in different tissues [18,19,20], such as the thyroid gland [21,22,23].
Given that the substrates of Fenton reaction are essential for thyroid hormone synthesis, yet—when in excess—can also inflict notable cellular harm and disrupt regular function, it is likely that thyroid epithelial cells possess a robust defense mechanism to counteract potential damage induced by free radicals. These protective mechanisms comprise antioxidative enzymes, such as superoxide dismutase (SOD), glutathione peroxidase (GPx), glutathione reductase (GR), catalase (CAT), and recently discovered peroxiredoxins [24]. In agreement with such an assumption, we made the following observations. The basal level of lipid peroxidation (LPO)—the index of oxidative damage to membrane lipids—was not higher in the thyroid gland than in other tissues [25]. Next, Fenton reaction-induced lipid peroxidation in the thyroid was lower than that in non-endocrine tissues such as the liver, kidney, brain cortex, spleen, small intestine [20], and ovary [25]. However, under conditions associated with endogenous abnormalities or with exposure to exogenous pro-oxidative agents, antioxidative machinery may become inefficient, resulting in huge oxidative damage and, consequently, in the development of various diseases, including cancer [26]. Interestingly, our studies also indicate that under basal conditions, female thyroid cells are exposed to higher concentrations of H2O2, most likely due to the activity of NADPH oxidases, primarily NOX4, and to a lesser extent, DUOX1 and DUOX2 [27], which can explain—at least partially—the higher prevalence of thyroid diseases, such as cancer, in the female population.
It should also be stressed that iodine, the fundamental building component of thyroid hormones, serves as a potent oxidizing agent. It can act as both an antioxidant and a prooxidant, depending on its chemical form. On the one hand, iodide ion (I−) is a potent antioxidant, the reducing properties of which make it an important free radical scavenger [28]. On the other hand, hypoiodite anion (IO−), which is oxidized iodine, is a potent oxidant with strong bactericidal activity. Moreover, an excess of iodine can interfere with thyroid hormone synthesis and, therefore, can act as a potential endocrine-disrupting chemical (EDC, endocrine disruptor), and this action occurs—at least partially—via mechanisms involving oxidative stress (reviewed in detail in [29]).
2. Melatonin as an Antioxidant—The Short Overview
Melatonin (N-acetyl-5-methoxytryptamine) is a ubiquitous molecule with a broad presence in nature, synthesized by a multitude of living organisms. It is postulated to have emerged on Earth approximately 3.2 to 3.5 billion years ago in photosynthetic cyanobacteria. Its synthesis in these organisms served as an essential antioxidant mechanism to counteract the production of harmful ROS during the photosynthetic process [30,31]. In accordance with the endosymbiotic theory regarding the origin of eukaryotic organelles [32], these organisms became integral components of eukaryotic physiology, together with the melatonin synthesis machinery [33]. These attributes, having conferred notable advantages to the functionality of the emerging eukaryote, were subsequently conserved and, throughout the course of evolution, were adapted for a number of other critical functions [31]. Nevertheless, the original antioxidative function of melatonin was retained and persists in present-day mammals, including humans.
In mammals, the major site for melatonin synthesis is the pineal gland. In addition, melatonin is produced to a lesser extent by numerous peripheral organs, including the bone marrow, lymphocytes, eyes, gastrointestinal tract, and human and rodent skin. Melatonin synthesis and secretion are suppressed by light and enhanced by dark [34]. The synthesis of melatonin involves a multi-step process, commencing with tryptophan as the precursor, followed by successive enzymatic conversions leading to the formation of serotonin, and culminating in the conversion to melatonin [35]. It should be stressed that melatonin is a lipophilic compound that can easily penetrate biological membranes [36].
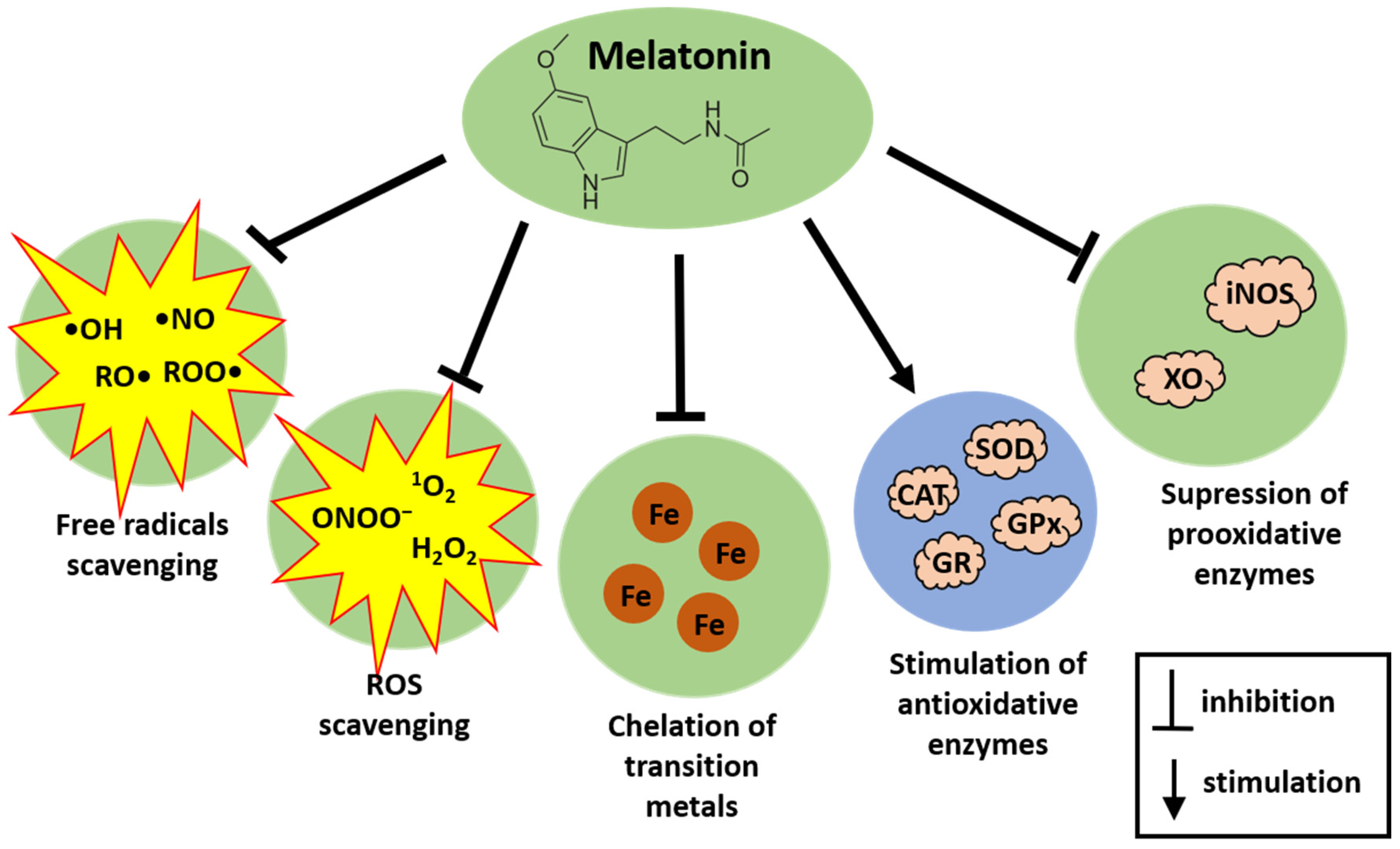
Melatonin achieves its antioxidative action via a variety of means. Melatonin effectively neutralizes a broad spectrum of free radicals, including the highly reactive and destructive •OH, alkoxy radicals (RO•), peroxy radicals (ROO•), and nitric oxide (•NO) [37]. In addition, melatonin also reportedly chelates transition metals, which are involved in the Fenton reaction; in doing so, melatonin reduces the formation of •OH [38]. Additionally, it targets various non-radical oxidants like H2O2, singlet oxygen (1O2), and peroxynitrite anion (ONOO−) [39]. As an electron-rich molecule, melatonin directly interacts with ROS, subsequently producing the following stable metabolites: cyclic 3-hydroxymelatonin, N1-acetyl-N2-formyl-5-methoxykynuramine, and N1-acetyl-5-methoxykynuramine [36,37]. Each of these metabolites also functions as a free radical scavenger. As a result, a single melatonin molecule along with its metabolites can neutralize numerous ROS [36,37]. Moreover, melatonin and its metabolites, contrary to antioxidants such as vitamin C, vitamin E, and glutathione (GSH), do not undergo redox cycling, and, thus, they do not promote secondary oxidation [36]. Also of importance, end-products of antioxidative action of melatonin are stable and excreted in the urine [40].
Besides directly neutralizing free radicals, melatonin can stimulate antioxidative enzymes such CAT, SOD, GPx, and GR [41,42,43]. At the same time, melatonin suppresses the activity of pro-oxidant enzymes, such as inducible nitric oxide synthase [44] or xanthine oxidase, the latter enzyme generating ROS produced after exposure to microwave radiation [45].
The physiological blood concentration of melatonin in humans depends on several factors, especially light and dark exposure. During the daytime, it is found to be 0–20 pg/mL, and at night it can reach levels as high as 40–200 pg/mL [46]. The nocturnal melatonin peak generally decreases with age, although there are notable individual variations. Some elderly individuals exhibit rather low nighttime melatonin levels that are nearly identical to their daytime levels, while others maintain a distinct melatonin rhythm with only modest reductions in nocturnal values [47]. The melatonin level in organisms can also be influenced by the consumption of melatonin-rich foods [48] or administration of aminoacids required for melatonin production [49]. Additionally, melatonin is increasingly being used as an over-the-counter dietary supplement mainly for the treatment of circadian rhythm sleep disorders [50]. Furthermore, it has gained recognition as a supplement that supports conventional drug therapies in various conditions, such as multiple sclerosis [51] and cancer [52]. Of importance, melatonin has a very good safety profile, which was also documented when this indoleamine was applied at very high doses. Some mild side effects, such as sleepiness, nausea, dizziness, and headache, have been observed exclusively during long-term treatment [53]. Usually recommended daily doses of melatonin range from 2 to 10 mg. In turn, as a high dose as 25 mg was used in clinical trials [54]. It is worth mentioning that intravenous administration of melatonin at a dose of 25 mg resulted in its blood concentration of approximately 7.52 × 105 pg/mL [54].
It is worth stressing that melatonin stands out among various hormones implicated in thyroid health due to its potent antioxidative effects, which are critical in the context of thyroid disorders. Unlike many other hormones that influence thyroid function through direct regulatory mechanisms, melatonin primarily contributes through its ability to modulate oxidative stress, a key factor in thyroid physiology and probably thyroid pathology.
Thyroid disorders, including both hypothyroidism and hyperthyroidism, are often accompanied by an increased oxidative stress burden due to abnormal metabolic rates associated with synthesis, degradation, and peripheral effects of thyroid hormones. The fact that melatonin is a highly effective free radical scavenger and is able to stimulate antioxidative enzymes but inhibit prooxidative enzymes means this indoleamine can be particularly beneficial in the context of thyroid diseases. It helps to protect thyroid cells from oxidative damage, which can lead to cellular dysfunction or death, and plays a role in preventing the onset and progression of thyroid diseases. Therefore, it is highly probable that melatonin affects the thyroid gland using mostly mechanisms different from endocrine mechanisms, which distinguishes this hormone from other hormones acting via binding to specific receptors. Summarizing this issue, the potent antioxidative properties of melatonin set it apart from other hormones that influence thyroid function, offering a distinctive therapeutic potential in managing thyroid disorders.
Antioxidative actions of melatonin are presented in Figure 2.
3. Relationship between the Thyroid and Melatonin
The interplay between melatonin, a hormone governing circadian rhythms and sleep-wake cycles, and the thyroid gland, a key component of the endocrine system regulating metabolism, reveals intricate physiological connections. Earlier studies clearly showed the profound impact of melatonin on the thyroid gland, indicating its direct inhibitory effects on both growth processes and thyroid hormone synthesis (reviewed in detail in [55]). It was also observed that melatonin is able to affect thyroid function and thyroid hormone concentrations in the blood, acting at the level of the hypothalamus, the pituitary gland, and/or the thyroid gland [56]. In photoperiodic species, namely animals that rely on day length or night length for biological and behavioral changes such as growth, migration, reproduction, and rest periods, melatonin coordinates the local control of thyroid hormone metabolism within the medio-basal hypothalamus, thereby initiating the photoperiodic response [57,58,59,60].
Melatonin can have also a direct effect on the activity of the thyroid gland. Interestingly, C cells but also thyroid follicular cells express mRNAs of the key enzymes in melatonin biosynthesis (i.e., aralkylamine N-acetyltransferase (AANAT) and acetyl serotonin methyltransferase (ASMT)), and they also possess melatonin receptor (MT1) [61]. It has also been observed that thyroid C cells synthesize melatonin under TSH control [62]. The significance of both phenomena remains to be elucidated.
Further investigations have found that daily administration of melatonin decreased T3 and T4 concentrations in dogs [63] and also suppressed thyroid hormone secretion in hyperthyroid rats [64]. Conversely, in the study on rats with propylthiouracil-induced hypothyroidism, a significant decrease in melatonin levels has been observed [65]. These findings underscore a complex and bidirectional interaction between melatonin and thyroid hormone homeostasis.
The relationship between the thyroid and melatonin documented in experimental studies is presented in Figure 3.
4. Potential Protective Effects of Melatonin against Carcinogenesis in Humans, Human Tissues, or Human-like Cell Lines
Due to such properties of melatonin as antioxidative, anti-inflammatory, immune-modulating, and endocrine effects, this indoleamine may also modulate the process of carcinogenesis. Indeed, it was found to exhibit anticarcinogenic action via antiproliferative, antiangiogenic, apoptotic, and metastasis-inhibitory pathways in experimental models. For example, antiproliferative effects of melatonin occurred via downregulation of oncogenic factors, such as cyclin D1 in MCF-7 breast cancer cells [66] or of histone deacetylase 9 (HDAC9) in non–small cell lung cancer [67]. The antiangiogenic properties of melatonin were demonstrated in human HepG2 liver cancer cells, where this indoleamine interferes with vascular endothelial growth factor (VEGF) [68]. Its ability to inhibit metastasis is evident in breast and lung cancers, where it disrupts key signaling pathways such as DJ-1/KLF17/ID-1 and epithelial-mesenchymal transition, respectively; the latter is mediated through the MT1 receptor and several downstream effectors including PLC, p38/ERK, and β-catenin [69,70]. In turn, in studies in humans with solid or liquid tumors, melatonin improved the sensitivity to conventional treatment [71,72].
Although there is a considerable amount of experimental research on the anticancer therapeutic properties of melatonin, studies involving humans are comparatively limited. Moreover, considering the wide spectrum of cancer types, anticancer therapies, and the range of melatonin actions, reaching a consensus about potential advantageous effects of melatonin applied in humans is challenging. The following examples of the therapeutic effects of melatonin in cancer patients are worth mentioning: a higher rate of tumor regression in patients with colorectal and gastric cancer [73], a higher survival rate in patients with solid neoplasms [74], and improvement of the chemotherapeutic effects of 5-fluorouracil [73], frequently employed in the treatment of various neoplasms. Another example of using melatonin as an anticancer adjuvant is its co-administration with platinum-based neoadjuvant chemotherapy for oral squamous cell carcinoma. This study demonstrated that administering 20 mg/daily of melatonin for seven days before and during therapy could reduce the expression of hypoxia-inducible factor (HIF-1α), which is a transcription factor contributing to cancer development during hypoxic conditions [75]. Melatonin co-administrated with chemotherapy has also reduced residual tumor percentage; however, it did not affect clinical response to neoadjuvant chemotherapy [75].
Another equally important aspect of melatonin’s influence on cancer therapy is its potential to have a broad impact on the adverse effects of anticancer medications and on the overall well-being of cancer patients. An example is the neuroprotective effects of melatonin counteracting adverse effects of adjuvant chemotherapy applied in breast cancer patients [76].
In our studies, we have observed that melatonin can completely prevent oxidative damage to membrane lipids caused by sorafenib or lenvatinib (multi-targeted tyrosine kinase inhibitors approved to treat advanced hepatocellular carcinoma, renal cell carcinoma, and radioiodine-refractory differentiated thyroid carcinoma) in noncancerous porcine tissues of the thyroid, liver, and kidney [77]. As both drugs can cause adverse effects resulting partially from ROS generation [78], melatonin can be considered to be applied in co-treatment with these tyrosine kinase inhibitors to prevent their undesirable toxic effects occurring via oxidative stress.
Although there has not been a dedicated study examining melatonin as a treatment for thyroid cancer in humans, melatonin shows some therapeutic effects in experimental studies.
As was mentioned above, melatonin minimizes thyroid cell damage due to irradiation [79,80]. On the other hand, some recent studies have proposed that melatonin can enhance the therapeutic effects of radiotherapy by acting as a radiosensitizer in thyroid cancer cells [81].
In another study with the use of a cell line from anaplastic thyroid cancer, melatonin exerted a dose-dependent cytotoxic effect. It significantly decreased cell viability and induced cell reproductive death at concentrations greater than 1 mM [82]. Such findings suggest that melatonin could be valuable as an adjuvant in anaplastic thyroid cancer therapy.
Research on a mouse model of thyroid cancer has demonstrated that melatonin treatment increased the production of the hydrogen sulfide (H2S) signaling molecule, which in turn increased the expression of pro-apoptotic tumor suppressor, resulting in the activation of cell apoptosis and suppression of cancer cell proliferation [83].
It is worth considering to what extent melatonin would be effective in particular types of thyroid cancer. The mechanisms behind the anticancer effects of melatonin are not sufficiently understood to allow for comparison across such diverse cancers as papillary and follicular cancer, both differentiated thyroid cancer formed from thyroid follicular cells and medullary thyroid cancer formed from C cells. Additionally, both types of thyroid cells possess the machinery for thyroid hormone synthesis as well as melatonin receptors. Therefore, it is currently challenging to conclusively determine if the effects of melatonin may vary specifically among these types of thyroid cancer due to the limited understanding of its underlying anticancer mechanisms.
The mechanisms of the potential protective effects of melatonin against the process of carcinogenesis are presented in Figure 4.
5. Evidence on Protective Effects of Melatonin against Oxidative Damage to Macromolecules Caused by Documented/Potential Carcinogens in the Thyroid
5.1. Melatonin and Oxidative Stress in the Thyroid
Thus summarizing results published so far on the protective effects of melatonin against oxidative damage caused by documented or potential carcinogens is justified. Whereas it is reasonable to consider such an action of melatonin in different tissues, the thyroid gland does constitute such an organ, which is characterized by relatively high levels of oxidative stress. Various in vitro and in vivo models were used to study the protective effects of melatonin against unfavorable oxidative processes in the thyroid. Regarding studies in humans, only a few publications on this issue are available in the literature.
Different aspects of oxidative stress were examined, including oxidative damage to macromolecules (such as membrane lipids, proteins, DNA, and other nucleic acids), activities of pro- and antioxidative enzymes, and levels of free radicals and other reactive oxygen species.
Taking into account the “oxidative nature” of the thyroid and the strong antioxidative properties of melatonin, it is especially interesting to investigate the extent to which melatonin affects favorably oxidative damage in this organ. Foremost, several in vitro studies performed by our team have shown that melatonin alone did not reduce LPO levels below the physiological threshold in the thyroid gland [20,21,25,84,85]. No changes in LPO levels in response to melatonin exposure were also observed in other tissues such as the spleen, liver, brain cortex, kidney, or intestine both in vitro and in vivo [20,86,87]. This action may offer an additional advantage since oxidative processes occur with a certain intensity in biological structures and are essential for physiological processes. However, these results neither imply that melatonin is ineffective as an antioxidant nor exclude its potential to prevent oxidative changes resulting from the action of any pro-oxidant. In fact, in the case of oxidative damage caused by pro-oxidative agents, we have observed that melatonin is an effective protective antioxidant in the thyroid gland.
In most studies cited below, melatonin revealed protective effects when used in concentrations ranging from nM to mM under in vitro conditions or when used in doses from 1 to 10 mg/kg under in vivo conditions.
5.2. Protective Effects of Melatonin against Oxidative Damage Caused by Documented Carcinogens (Listed in the International Agency for Research on Cancer (IARC) Monographs)
5.2.1. Hexavalent Chromium/Potassium Dichromate
Hexavalent chromium (Cr(VI)) is a highly toxic form of chromium used in various industrial processes. Exposure to Cr(VI) is prevalent in industries such as stainless steel production, chrome plating, leather tanning, and chemical dye production. Exposure primarily occurs through inhalation of contaminated air or ingestion of water polluted with Cr(VI).
Chromium (VI) compounds are classified by the International Agency for Research on Cancer (IARC) as Group 1 carcinogens [88], indicating that they are carcinogenic to humans. The following cancer types are best documented to be associated with the exposure to Cr(VI): cancers of the lung, nasal cavity, and paranasal sinuses; however, Cr(VI) may also increase the risk of brain cancer, malignant lymphoma, and tumors of the endocrine glands [89]. Cr(VI) induces cellular damage and adverse health effects through various mechanisms such as oxidative stress, DNA damage, dysregulating gene expression, and cancer-related signal transduction [90]. Upon entry into cells, Cr(VI) is reduced by ascorbate and biological thiols such as GSH; this process generates H2O2 and other free radical species that may contribute to increased oxidative stress causing damage to cellular lipids, proteins, and DNA [91]. Exposure levels to Cr(VI) can range from background environmental levels of less than 1 microgram per cubic meter (µg/m3) to over 100 µg/m3 in occupational settings involving chrome plating, stainless steel welding, and manufacturing processes.
As demonstrated in studies on adult male albino rats, the administration of potassium dichromate (K2Cr2O7) (a hexavalent chromium compound), Cr(VI)) in the dose of 25 mg/kg/day for 2 months leads, among other effects, to follicular hyperplasia and follicular enlargement in the thyroid [92]. In this study, the simultaneous administration of melatonin in the dose of 10 mg/kg/day demonstrated a strong protective effect, as thyroid follicular cells from animals treated with K2Cr2O7 plus melatonin appeared ultrastructurally similar to those of the control group. The above effect surpassed that of vitamin C, highlighting melatonin’s potential as a mitigating factor against the adverse impacts of Cr(VI) exposure on thyroid function [92].
5.2.2. Radiotherapy/Ionizing Radiation
Radiotherapy is a type of cancer treatment that uses high-energy electromagnetic radiation (such as gamma or X-rays) or radiation of particles (such as protons, neutrons, and high-speed electrons). Passing through the cells, this ionizing radiation can cause the displacement of electrons from atoms, which, in turn, produces biological effects by breaking chemical bonds and forming highly reactive free radicals. This process can induce DNA damage either by direct means (DNA damage directly caused by ionization) or indirectly (through the production of free radicals), consequently leading to cell death.
According to the IARC classification, all types of ionizing radiation belong to Group 1 carcinogens (i.e., carcinogenic to humans) [93]. Since radiotherapy is commonly employed in patients with head and neck cancer or breast cancers, and also lymphomas and central nervous system tumors, the radiation in these regions often affects the thyroid gland. It is worth mentioning that the most common complication that can occur after radiation therapy for breast or head and neck cancers is hypothyroidism [94], but in rare cases, new systemic and local cancers, including thyroid cancer, can appear [95]. Radiation therapy doses, measured in Gray (Gy), for treating various cancers range from 10 to 70 Gy, depending on cancer type, stage, treatment goal, and area treated.
Studies conducted on rats have shown that radiation of the neck region with a total dose of 16–18 Gy led to increased inflammation, vacuolization, degradation, swelling, and necrosis in the thyroid gland [79]. Additionally, there were elevated levels of tumor necrosis factor-α (TNF-α), interleukin-1β (IL-1β), thiobarbituric acid reactive substances (TBARS), and nitric oxide (NO) [80]. Melatonin at doses of 10–50 mg/kg administered 10–15 min before exposure to radiation caused a significant reduction of these unfavorable changes in histopathological and biochemical parameters [79,80]. Moreover, GSH values were higher in the melatonin plus radiotherapy group compared to radiotherapy alone [80].
It is worth mentioning that the antioxidative effects of melatonin in protection against cellular damage caused by ionizing radiation were summarized over 20 years ago by one of the authors of the present review [96]; however, the thyroid tissue had not been examined in these earlier studies.
5.2.3. Radioiodinetherapy/Iodine-131
Radioisotopes of iodine are called radioiodine or radioactive iodine (RAI). Iodine-131 (I-131) is a radioisotope of iodine that primarily emits β rays and—to a much lesser extent—γ rays. With a relatively short half-life of just over 8 days, it remains the radionuclide of choice for both therapeutic and diagnostic applications. I-131 therapy is indicated for the management of hyperthyroidism and thyroid cancer. Its action is dependent on the uptake of iodine by thyroid tissue. Once absorbed by thyroid follicular cells, I-131 emits β rays, a form of ionizing radiation that induces local damage to thyroid follicular cells.
Radioiodines, including I-131, have been classified by the IARC as Group 1 carcinogens, indicating their carcinogenicity to humans [97]. In the case of RAI therapy for hyperthyroidism, the most recent meta-analysis revealed no significant elevation in risk for specific types of cancer, with the exception of thyroid cancer incidence, for which the risk was statistically higher [98]. Regarding RAI therapy for differentiated thyroid cancer, it has been found that this treatment in childhood and young adulthood was associated with an increased risk of solid cancer (by 23%) and leukemia (by 92%), particularly more than 20 years after exposure [99].
Due to these undesirable effects of radioiodine, it is important to determine possible protective factors. In animal studies conducted on rats, melatonin in a dose of 12 mg/kg/day administered 2 days before and for 1 week after oral RAI prevented adverse effects of a single dose of 111 MBq (3 mCi) RAI on spermatozoa quality [100] and on the oxidative stress parameters in the liver [101]. A capsule containing 300 mg of melatonin powder given 1 h before the administration of iodine-131 (370 to 740 MBq (10 to 20 mCi)) slightly reduced (without statistical significance) the rate of I-131-induced chromosomal damage in lymphocytes from hyperthyroid patients [102].
Regarding isotope I-123, which emits mostly γ radiation and is used almost exclusively for diagnostic imaging, no results are found in the literature on its damaging effects on macromolecules.
5.2.4. Night-Shift Work
Night-shift work is defined as work occurring during the regular sleeping hours of the general population. According to the IARC classification, night-shift work belongs to Group 2A (probably carcinogenic to humans) [103]. There is sufficient evidence in experimental animals for the carcinogenic effects of alterations in the light–dark schedule. However, evidence of the carcinogenic effects of night-shift work in humans is limited. The hypothesized contributing factors are directly related to disruptions in the circadian rhythm and decreased melatonin production, both considered oncogenic in nature [104]. The circadian rhythm is a fundamental internal biological process, which regulates crucial mechanisms such as metabolism, DNA repair, and immune system function, all integral to cancer pathogenesis. The circadian rhythm enables human bodies to function optimally in a 24-h cycle.
The IARC Working Group based its decision to classify night-shift work as probably carcinogenic to humans (Group 2A) on evidence primarily derived from studies on breast cancer risk and, to a lesser extent, on prostate and colorectal cancer risk [103]. Studies examining the correlation between night-shift work and other types of cancer are limited. Regarding the correlation between night-shift work and thyroid cancer risk, a study among nurses found no direct link [105]. However, the research did suggest a potential association between persistent sleep difficulties among night-shift workers and a higher risk of thyroid cancer. Notably, nurses with over a decade of night-shift experience, coupled with frequent sleep difficulties, showed a modestly increased risk of thyroid cancer [105]. Additionally, in a large US cohort study, it was found that exposure to light at night (LAN) was positively associated with thyroid cancer risk [106]; this relationship may be directly related to the fact that exposure to artificial LAN inhibits nighttime secretion of melatonin and subsequently may cause circadian disruption [107].
Despite the well-established role of melatonin in circadian rhythm regulation, there has not been a dedicated study examining melatonin as a preventive treatment for night-shift workers. This underscores the need for further research to explore the potential protective effects of melatonin in mitigating the adverse health impacts associated with night-shift work, particularly in the context of cancer risk.
5.2.5. Nitrobenzene
Nitrobenzene is a synthetic chemical not found naturally in the environment. It is widely used in industry, primarily in the synthesis of aniline—a commodity chemical produced on a very large scale and used as a precursor of polyurethane, dyes, and other industrial chemicals. Given its extensive industrial usage, nitrobenzene is ubiquitously present in the environment, increasing the likelihood of hazardous exposure.
Nitrobenzene is classified by the IARC as a Group 2B compound (possibly carcinogenic to humans) [108]. It has been demonstrated that chronic inhalation exposure of experimental animals to nitrobenzene is associated with an increased incidence of tumors at multiple sites, including follicular thyroid tumors [109]. The U.S. Environmental Protection Agency, based on studies in rats and mice, determined that the Reference Dose (RfD) for nitrobenzene is 0.0005 mg/kg/day [110]. This RfD represents an estimate of a daily human exposure (including sensitive subpopulations) that is likely to be without a significant risk of adverse effects over a lifetime. The observed toxic effects of nitrobenzene include an increased incidence of thyroid follicular adenomas in rats and mice. Additionally, endometrial tumors were reported in the same species [111].
In the research conducted by our team, nitrobenzene in concentrations of 7.5 and 10.0 mM increased LPO levels in porcine thyroid homogenates [112]. In this study, we have shown that melatonin in concentrations as low as 0.0001 mM effectively prevented LPO damage caused by nitrobenzene (7.5 mM) [112]. It should be stressed that such low concentrations of melatonin (0.0001 mM) are only two orders of magnitude higher than physiological melatonin blood concentration in humans i.e., 0.0001 mM vs. 0.000001 mM [46].
5.2.6. Bromium/Potassium Bromate
Potassium bromate (KBrO3) has been classified by the IARC as a compound belonging to Group 2B carcinogens (a possible human carcinogen) [113]. KBrO3 is a strong oxidizing agent. It was commonly used in the past as a food additive in the bakery industry, but in some countries industrial use of KBrO3 has persisted.
Oxidative properties of KBrO3 play a fundamental role in its carcinogenic action. This compound increases the formation of free radicals and reactive oxygen species (ROS), such as peroxynitrite anion (ONOO−) or nitric oxide (NO•), and—simultaneously—it decreases the activity of antioxidative enzymes, such as GPX [114]. KBrO3 was found to cause oxidative DNA damage, as measured by the increased level of 8-hydroxy-2′-deoxyguanosine (8-OHdG), and to induce mutations (GC to TA transversions) in the rat kidney [115].
In studies conducted by our team, we investigated the effects of melatonin and another indole substance possessing a chemical structure similar to that of melatonin, indole-3-propionic acid (IPA), on LPO induced by KBrO3. We found that neither melatonin (at concentrations ranging from 0.01 mM to 7.5 mM) nor IPA (at concentrations ranging from 0.01 mM to 10 mM) reduced LPO induced by 5 mM KBrO3 in porcine thyroid homogenates [116]. However, when melatonin or IPA was used in vivo in rats as a pretreatment, it decreased LPO induced by a single injection of KBrO3 in the dose of 110 mg/kg in the thyroid gland [116]. This protective effect was achieved by administering melatonin or IPA at a dose of 0.0645 mmol/kg (i.e., 15 mg/kg) b.w. twice daily for 10 days. It is worth mentioning that not only in vivo but also in vitro damaging effects of KBrO3 were prevented by propylthiouracil, a well-known antithyroid medication that possesses certain antioxidative properties [116]. However, taking into account clinical applications of the above results [116], only melatonin or IPA can be considered in preventive actions, as propylthiouracil would cause undesired side effects such as hypothyroidism.
5.2.7. Mercury
Mercury (Hg) is one of the top ten chemicals of major public health concerns as stated by the World Health Organization (WHO) [117]. This chemical is a well-known toxic metal, causing occupational but also accidental exposures and, consequently, damage in human and animal organs. Although several observations link Hg exposure to cancer, the scientific evidence regarding the potential role of Hg in carcinogenesis is not clear [118].
According to the IARC classification, inorganic Hg species belong to Group 3 agents (agents with inadequate evidence for carcinogenicity in humans and experimental animals) [119]. In contrast, methylmercury (MeHg)—a compound formed from inorganic mercury by the action of microbes that live in aquatic systems—is categorized by the IARC as a Group 2B compound (potentially carcinogenic to humans) [119]. Although the RfD for elemental mercury was not assessed, it was determined to be 0.1 µg/kg/day for methylmercury [120].
Due to their toxic properties, mercury and its derivatives can induce oxidative damage, genotoxicity, and autoimmune reactions. Such adverse phenomena have also been observed in the thyroid gland [121,122]. For instance, it has been shown that occupational exposure to mercury is associated with RNA oxidative damage, as measured by the increased level of 8-oxo-7,8-dihydroguanosine (8-oxoGuo) [123]. These findings suggest a potential role of mercury and its derivatives in the pathogenesis of thyroid cancer, autoimmune thyroiditis, and hypothyroidism. A recently published meta-analysis has revealed an association between mercury exposure and thyroid cancer risk, implying a possible predisposing factor; however, further research is necessary to define the clinical relevance of this relationship [124].
In studies on adult male albino rats of the Wistar strain, it has been shown that the administration of mercury chloride (inorganic mercury compound) in the dose of 2–4 mg/kg b.w. caused a significant increase in oxidative stress in the thyroid gland imposed by a significant decline in levels of antioxidative enzymes such as SOD, CAT, GPX, GR, and non-enzymatic antioxidant GSH, followed by an elevated level of LPO [125]. Co-administration of melatonin in the dose of 5 mg/kg b.w. partially protected against mercury-induced changes [125].
5.3. Protective Effects of Melatonin against Oxidative Damage Caused by Potential Carcinogens (Not Listed in the International Agency for Research on Cancer Monographs)
5.3.1. Potassium Iodate
Iodine is an essential element for the synthesis of thyroid hormones. Iodine deficiency can lead to hypothyroidism and, consequently, to metabolic and developmental disorders, and it is associated with a higher risk of follicular thyroid cancer. Therefore, many countries have implemented iodine prophylaxis programs to eliminate iodine deficiency. Although excessive iodine intake may induce thyroid dysfunction and increase the relative risk of papillary thyroid cancer, the benefits outweigh the risks [126].
In most programs of iodine prophylaxis, iodized salt is used with either potassium iodide (KI) or potassium iodate (KIO3), as iodine carriers. It is known that these two iodine compounds possess distinct pro- and antioxidative properties. Unlike KI, KIO3 acts as an oxidant and can readily react with oxidizable substances [29].
Despite the chemical similarity of KIO3 to the previously described KBrO3, studies on the oxidative properties of iodate have shown that it has low, if any, genotoxic potential [127]. For this reason, KIO3 is not included in the IARC carcinogen list. However, in our study, we found that KIO3 can induce oxidative damage to membrane lipids in homogenates of several tissues, including thyroid tissue.
We have observed that KIO3 at concentrations of 20, 15, 10, 7.5, and 5 mM increased LPO in various porcine tissues, including the thyroid, ovary, liver, kidney, brain, spleen, and small intestine. Notably, the damaging effect of KIO3 at 10 and 7.5 mM was lower in the thyroid than in other tissues, and the lowest concentration of 5 mM did not exhibit any damaging effect in the thyroid. It is worth mentioning that these concentrations of KIO3 are close to the physiological iodine levels in the thyroid (approx. 9.0 mM). Whereas melatonin (5 mM) reduced LPO induced by 10, 7.5, and 5 mM of KIO3 in all tissues, in the thyroid it was additionally protective against as high a concentration of KIO3 as 15 mM, indicating that melatonin has a particularly beneficial role in the thyroid gland. Furthermore, LPO levels in the thyroid (vs. other tissues) were found to be lower when exposed to KIO3 in conjunction with melatonin. This disparity in LPO levels may suggest that the damaging effect of KIO3 is weaker in the thyroid than in other tissues, highlighting the tissue-specific protective role of melatonin against oxidative stress [25,84,85]. Moreover, cumulative protective effects of melatonin (5 mM) and IPA (10 mM) in the thyroid homogenates were stronger than those revealed by each of these two antioxidants used separately [84]. Interestingly, we have also observed that potassium iodide (KI), another iodine compound used in salt iodization programs, did not induce LPO in the thyroid when used in concentrations similar to those, in which KIO3 revealed damaging effects; moreover, it even reduced oxidative damage induced by the Fenton reaction substrates [128]. Our observations suggest that melatonin could be an effective agent in preventing oxidative damage and potentially reducing the risk of thyroid cancer formation caused by iodine compounds used in iodine prophylaxis.
It is worth mentioning that iodine when in excess is considered to be not only a potential carcinogen but also a potential endocrine disruptor [29].
5.3.2. Iron-Induced Oxidative Damage
Iron, a crucial micronutrient, acts as a cofactor in fundamental cellular processes, including oxygen transport, cell proliferation, and energy metabolism. However, an imbalance leading to iron overload can induce heightened oxidative stress, potentially resulting in adverse effects. Although iron is not considered by the IARC as a human carcinogen, numerous animal models unequivocally indicate that an excess of iron can contribute to carcinogenesis. This is further substantiated by a range of human epidemiological data concerning cancer risk and prognosis [129].
Our first study on the potential protective effects of melatonin against experimentally induced oxidative damage in the thyroid was performed with the use of a classic experimental model. In the in vitro study, we applied Fenton reaction substrates, i.e., ferrous ions (Fe2+), in the concentration of 40 µM and hydrogen peroxide (H2O2) in the concentrations of 0.5 mM to induce LPO in porcine thyroid homogenates [21]. Expectedly, Fenton reaction substrates increased the level of LPO, and this damaging effect was prevented by melatonin in a concentration-dependent manner, with complete reduction to a control level when melatonin was added in the concentration of 5 mM. Interestingly, in our more recent work, we demonstrated that melatonin in the concentration of 5 mM is also highly effective in protecting against oxidative damage induced by extremely high concentrations of iron, i.e., as high as 4.8 mM. The protective effects of melatonin were observed not only in porcine thyroid but also in various non-endocrine porcine tissues (liver, kidney, brain cortex, spleen, and small intestine) [20]. The high iron concentrations used in our study [20] correspond to those found in patients with iron overload caused by congenital disturbances of iron metabolism (hemochromatosis) and by secondary hemochromatosis resulting from repeated blood transfusions or overconsumption of iron. Local iron overload can also be caused by chronic hepatitis C, ovarian endometriosis, or asbestos exposure [130]. This is particularly important when we take into account the fact that the carcinogenicity of iron has been clearly shown both in animal models and human studies [131]. In fact, hemochromatosis, chronic hepatitis C, or ovarian endometriosis are associated with an increased risk of cancer [130].
5.3.3. Endocrine Disruptors
Endocrine disruptors are exogenous chemicals or mixtures of chemicals that interfere with any aspect of hormone action [132]. Compounds such as ammonium thiocyanate (NH4SCN), sodium fluoride (NaF), potassium selenocyanate (KSeCN), sodium chlorate (NaClO3), potassium perchlorate (KClO4), potassium nitrate (KNO3), and bisphenol A (BPA) pose significant risks to thyroid health. They can disrupt iodine uptake through inhibition of the sodium/iodide symporter (NIS) activity, which is crucial for active iodine transport into the thyroid; consequently, they inhibit thyroid hormone synthesis and metabolism [133]. They are shortly called NIS inhibitors. Although these endocrine disruptors are not specifically listed as carcinogens by the IARC, it has been observed that they can act through the induction of oxidative stress, and they are generally considered potential carcinogens. Therefore, in our recent study, we examined the damaging effects of these NIS inhibitors and the potential protective effects caused by melatonin and IPA [134].
Ammonium thiocyanate (NH4SCN) is utilized across a wide range of industries and chemical processes. It is commonly employed in the production of herbicides and thiourea and as a stabilizing agent in textile and dyeing processes. Its metabolic transformation in organisms may lead to the formation of reactive species, such as thiocyanate ions (SCN−), which can participate in reactions generating ROS [135]. In our in vitro study, NH4SCN used in concentrations of 250–500 mM increased LPO levels in thyroid homogenates, while both melatonin and IPA, at concentrations of 5 mM, completely prevented these changes [134].
Sodium fluoride (NaF) is a chemical compound recognized for its role in promoting dental health, being widely used in toothpaste and drinking water to prevent tooth decay by strengthening tooth enamel. It also serves various industrial purposes, including acting as a flux in the manufacture of aluminum and as an insecticide. The toxicity caused by fluoride is attributed to increased oxidative stress and altered antioxidant defense mechanisms [136,137]. In the in vitro study performed by our team, NaF in concentrations of 25–100 mM induced LPO in thyroid homogenates. As expected, both melatonin (5 mM) and IPA (5 mM) completely prevented NaF-induced oxidative damage to membrane lipids [134].
Selenium (Se) is an essential trace element that plays a vital role in various biological processes. Its main biological role is related to its incorporation into selenoproteins, which, among other functions, participate in redox homeostasis and the metabolism of thyroid hormones. On the other hand, Se in higher concentrations can exert strong toxicity, in part by contributing to the formation of ROS [138]. Generally, it is shown that selenium compounds such as KSeCN can serve as antioxidants, impacting redox homeostasis and cellular health [139]. Although KSeCN is mostly known as an antioxidant and it has not been discussed until now in the literature as a potential carcinogen, we decided to check if this NIS inhibitor is able to induce oxidative damage in the thyroid. In our study, incubation of thyroid homogenates in the presence of KSeCN increased LPO in a concentration-dependent manner (statistically significant for 500 mM). Melatonin (5 mM) and IPA (5 mM) completely prevented KSeCN-induced LPO in the thyroid [134].
Sodium chlorate (NaClO3), a powerful oxidizing agent, is commonly used in the production of herbicides and explosives and in the bleaching process of paper pulp. Due to its properties, it can cause oxidative stress in organisms by generating ROS. It has been shown that the reduction of chlorate (ClO3−) to chloride ions (Cl−) results in the formation of ROS and free radicals, which can damage macromolecules [140]. In our in vitro study, NaClO3 (in concentrations of 0.5–10 mM) increased LPO levels in the thyroid; however, neither melatonin nor IPA prevented these oxidative damages [134].
Two NIS inhibitors applied in our study, i.e., potassium perchlorate (KClO4) and potassium nitrate (KNO3), did not change LPO in thyroid homogenates [134]. Thus, their damaging effects in the thyroid can be checked in other experimental models.
Bisphenol A is a synthetic chemical compound extensively used in the production of polycarbonate plastics and epoxy resins, found in a wide range of consumer goods such as water bottles, food containers, and the linings of metal cans [141]. It is a well-known endocrine disruptor. At the same time, potential carcinogenic effects of bisphenol A are suggested in the literature [142]. Bisphenol A has been demonstrated to increase H2O2 production in thyrocytes in both in vivo (40 mg/kg b.w. daily for 15 days) and in vitro (1 nM) settings [143], but neither melatonin nor other antioxidants were applied in this model for checking their potential protection. In our in vitro study, bisphenol A did not affect oxidative damage to membrane lipids in porcine thyroid homogenates [134]; therefore, the potential protective effects of melatonin against LPO induced by bisphenol A are still not known. However, other in vitro models should be applied to check if bisphenol A is able to induce oxidative damage to macromolecules in the thyroid gland.
The protective effects of melatonin against oxidative damage caused by potential carcinogens (not listed in the International Agency for Research on Cancer (IARC) Monographs) are summarized in Table 2 and presented in Figure 5.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 2.
Protective effects of melatonin against oxidative damage caused by potential carcinogens (not listed in the International Agency for Research on Cancer (IARC) Monographs). MDA + 4-HDA, malondialdehyde+4-hydroxyalkenals; ↑, increase.
Table 2.
Protective effects of melatonin against oxidative damage caused by potential carcinogens (not listed in the International Agency for Research on Cancer (IARC) Monographs). MDA + 4-HDA, malondialdehyde+4-hydroxyalkenals; ↑, increase.
| Agent/Dose | Species/Organ/Tissue/ Cellular Compartment | Effect of Agent | Dose of Melatonin, Which Reduced or Prevented the Effect of Agent | Refs. |
|---|---|---|---|---|
| Potassium iodate (KIO3) | Porcine thyroid homogenates | ↑MDA + 4-HDA | 5 mM | [25,84,85] |
| Ferrous ion (Fe2+)/ferrous sulfate (FeSO4) 40 µM or 37.5 µM–4.8 mM | Porcine thyroid homogenates | ↑MDA + 4-HDA | 5 mM | [20,21] |
| Ammonium thiocyanate (NH4SCN) 250–500 mM | Porcine thyroid homogenates | ↑MDA + 4-HDA | 5 mM | [134] |
| Sodium fluoride (NaF) 25–100 mM | Porcine thyroid homogenates | ↑MDA + 4-HDA | 5 mM | [134] |
| Potassium selenocyanate (KSeCN) 500 mM | Porcine thyroid homogenates | ↑MDA + 4-HDA | 5 mM | [134] |
| Sodium chlorate (NaClO3) 0.5–10 mM | Porcine thyroid homogenates | ↑MDA + 4-HDA | No protection | [134] |
6. Conclusions and Future Directions
This review has underscored the potent antioxidative effects of melatonin on the thyroid gland, highlighting its protective role against carcinogen-induced oxidative stress and its potential implications in thyroid health and disease. The evidence from various experimental models supports melatonin’s capacity to mitigate oxidative damage, the process being a critical factor in the pathogenesis of thyroid disorders, cancer included. However, the scarcity of human studies marks a significant gap in our understanding and application of these findings in clinical settings.
Future research should prioritize clinical trials to establish the therapeutic efficacy and safety of melatonin in human individuals with thyroid disorders. It is imperative to explore the molecular mechanisms underlying melatonin’s protective effects in greater detail, to identify potential targets for intervention. Moreover, longitudinal studies assessing the long-term outcomes of melatonin supplementation in individuals at risk of thyroid diseases could provide invaluable insights. The integration of melatonin into therapeutic regimens for thyroid conditions necessitates a multidisciplinary approach, combining insights from endocrinology, oncology, and pharmacology, to harness its full potential as a noninvasive, natural antioxidant in the prevention and treatment of thyroid diseases.
Author Contributions
Conceptualization, M.K.-L. and J.S.; methodology, M.K.-L.; software, J.S.; validation, M.K.-L. and J.S.; investigation, J.S.; data curation, J.S.; writing—original draft preparation, J.S. and M.K.-L.; writing—review and editing, M.K.-L.; visualization, J.S.; supervision, M.K.-L. All authors have read and agreed to the published version of the manuscript.
Funding
This study was funded by statutory funds from the Medical University of Lodz (503/1-107-03/503-11-001).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Maenhaut, C.; Christophe, D.; Vassart, G.; Dumont, J.; Roger, P.P.; Opitz, R. Ontogeny, Anatomy, Metabolism and Physiology of the Thyroid. In Endotext; Feingold, K.R., Anawalt, B., Blackman, M.R., Boyce, A., Chrousos, G., Corpas, E., de Herder, W.W., Dhatariya, K., Dungan, K., Hofland, J., et al., Eds.; MDText.com, Inc.: South Dartmouth, MA, USA, 2015. [Google Scholar]
- Szanto, I.; Pusztaszeri, M.; Mavromati, M. H2O2 Metabolism in Normal Thyroid Cells and in Thyroid Tumorigenesis: Focus on NADPH Oxidases. Antioxidants 2019, 8, 126. [Google Scholar] [CrossRef] [PubMed]
- Karbownik-Lewińska, M.; Kokoszko-Bilska, A. Oxidative damage to macromolecules in the thyroid–experimental evidence. Thyroid. Res. 2012, 5, 25. [Google Scholar] [CrossRef] [PubMed]
- Faria, C.C.; Fortunato, R.S. The role of dual oxidases in physiology and cancer. Genet. Mol. Biol. 2020, 43 (Suppl. S1), e20190096. [Google Scholar] [CrossRef] [PubMed]
- Grasberger, H. Defects of thyroidal hydrogen peroxide generation in congenital hypothyroidism. Mol. Cell. Endocrinol. 2010, 322, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Liu, W.; Zhang, L.; Wang, F.; Wang, F.; Gu, M.; Wang, X.; Liu, S. Identification and analyzes of DUOX2 mutations in two familial congenital hypothyroidism cases. Endocrine 2021, 72, 147–156. [Google Scholar] [CrossRef]
- Hulur, I.; Hermanns, P.; Nestoris, C.; Heger, S.; Refetoff, S.; Pohlenz, J.; Grasberger, H. A single copy of the recently identified dual oxidase maturation factor (DUOXA) 1 gene produces only mild transient hypothyroidism in a patient with a novel biallelic DUOXA2 mutation and monoallelic DUOXA1 deletion. J. Clin. Endocrinol. Metab. 2011, 96, E841–E845. [Google Scholar] [CrossRef] [PubMed]
- Szanto, I. NADPH Oxidase 4 (NOX4) in Cancer: Linking Redox Signals to Oncogenic Metabolic Adaptation. Int. J. Mol. Sci. 2022, 23, 2702. [Google Scholar] [CrossRef]
- Azouzi, N.; Cailloux, J.; Cazarin, J.M.; Knauf, J.A.; Cracchiolo, J.; Al Ghuzlan, A.; Hartl, D.; Polak, M.; Carré, A.; El Mzibri, M.; et al. NADPH Oxidase NOX4 Is a Critical Mediator of BRAFV600E-Induced Downregulation of the Sodium/Iodide Symporter in Papillary Thyroid Carcinomas. Antioxid. Redox Signal 2017, 26, 864–877. [Google Scholar] [CrossRef]
- Oglio, R.; Salvarredi, L.; Rossich, L.; Copelli, S.; Pisarev, M.; Juvenal, G.; Thomasz, L. Participation of NADPH 4 oxidase in thyroid regulation. Mol. Cell. Endocrinol. 2019, 480, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Wu, R.F.; Ma, Z.; Liu, Z.; Terada, L.S. Nox4-derived H2O2 mediates endoplasmic reticulum signaling through local Ras activation. Mol. Cell. Biol. 2010, 30, 3553–3568. [Google Scholar] [CrossRef]
- Malhotra, J.D.; Kaufman, R.J. Endoplasmic reticulum stress and oxidative stress: A vicious cycle or a double-edged sword? Antioxid. Redox Signal. 2007, 9, 2277–2293. [Google Scholar] [CrossRef] [PubMed]
- Andrés, C.M.C.; Pérez de la Lastra, J.M.; Juan, C.A.; Plou, F.J.; Pérez-Lebeña, E. Chemistry of Hydrogen Peroxide Formation and Elimination in Mammalian Cells, and Its Role in Various Pathologies. Stresses 2022, 2, 256–274. [Google Scholar] [CrossRef]
- Garofalo, V.; Condorelli, R.A.; Cannarella, R.; Aversa, A.; Calogero, A.E.; La Vignera, S. Relationship between Iron Deficiency and Thyroid Function: A Systematic Review and Meta-Analysis. Nutrients 2023, 15, 4790. [Google Scholar] [CrossRef] [PubMed]
- Abbas, W.; Elmugabil, A.; Hamdan, H.Z.; Rayis, D.A.; Adam, I. Iron deficiency and thyroid dysfunction among sudanese women in first trimester of pregnancy: A cross-sectional study. BMC Endocr. Disord. 2023, 23, 223. [Google Scholar] [CrossRef] [PubMed]
- Krohn, K.; Maier, J.; Paschke, R. Mechanisms of disease: Hydrogen peroxide, DNA damage and mutagenesis in the development of thyroid tumors. Nat. Clin. Pract. Endocrinol. Metab. 2007, 3, 713–720. [Google Scholar] [CrossRef] [PubMed]
- Penna, G.; Rubio, I.G.S.; Brust, E.S.; Cazarin, J.; Hecht, F.; Alkmim, N.R.; Rajão, K.M.A.B.; Ramos, H.E. Congenital hypothyroidism and thyroid cancer. Endocr. Relat. Cancer 2021, 28, R217–R230. [Google Scholar] [CrossRef] [PubMed]
- Rynkowska, A.; Stępniak, J.; Karbownik-Lewińska, M. Fenton Reaction-Induced Oxidative Damage to Membrane Lipids and Protective Effects of 17β-Estradiol in Porcine Ovary and Thyroid Homogenates. Int. J. Environ. Res. Public Health 2020, 17, 6841. [Google Scholar] [CrossRef] [PubMed]
- Rynkowska, A.; Stępniak, J.; Karbownik-Lewińska, M. Melatonin and Indole-3-Propionic Acid Reduce Oxidative Damage to Membrane Lipids Induced by High Iron Concentrations in Porcine Skin. Membranes 2021, 11, 571. [Google Scholar] [CrossRef]
- Stępniak, J.; Rynkowska, A.; Karbownik-Lewińska, M. Membrane Lipids in the Thyroid Comparing to Those in Non-Endocrine Tissues Are Less Sensitive to Pro-Oxidative Effects of Fenton Reaction Substrates. Front. Mol. Biosci. 2022, 9, 901062. [Google Scholar] [CrossRef]
- Karbownik, M.; Lewiński, A. Melatonin reduces Fenton reaction-induced lipid peroxidation in porcine thyroid tissue. J. Cell. Biochem. 2003, 90, 806–811. [Google Scholar] [CrossRef]
- Stępniak, J.; Lewiński, A.; Karbownik-Lewińska, M. Membrane lipids and nuclear DNA are differently susceptive to Fenton reaction substrates in porcine thyroid. Toxicol. Vitr. 2013, 27, 71–78. [Google Scholar] [CrossRef]
- Stepniak, J.; Lewinski, A.; Karbownik-Lewinska, M. Oxidative damage to membrane lipids in the thyroid–no differences between sexes. Drug Chem. Toxicol. 2021, 44, 655–660. [Google Scholar] [CrossRef] [PubMed]
- Kovacic, P.; Edwards, C. Integrated approach to the mechanisms of thyroid toxins: Electron transfer, reactive oxygen species, oxidative stress, cell signaling, receptors, and antioxidants. J. Recept. Signal Transduct. Res. 2010, 30, 133–142. [Google Scholar] [CrossRef] [PubMed]
- Iwan, P.; Stepniak, J.; Karbownik-Lewinska, M. Pro-Oxidative Effect of KIO3 and Protective Effect of Melatonin in the Thyroid-Comparison to Other Tissues. Life 2021, 11, 592, Erratum in Life 2022, 12, 592. [Google Scholar] [CrossRef] [PubMed]
- Cheung, E.C.; Vousden, K.H. The role of ROS in tumour development and progression. Nat. Rev. Cancer 2022, 22, 280–297. [Google Scholar] [CrossRef] [PubMed]
- Stepniak, J.; Lewinski, A.; Karbownik-Lewinska, M. Sexual Dimorphism of NADPH Oxidase/H2O2 System in Rat Thyroid Cells; Effect of Exogenous 17β-Estradiol. Int. J. Mol. Sci. 2018, 19, 4063. [Google Scholar] [CrossRef] [PubMed]
- Sorrenti, S.; Baldini, E.; Pironi, D.; Lauro, A.; D’Orazi, V.; Tartaglia, F.; Tripodi, D.; Lori, E.; Gagliardi, F.; Praticò, M.; et al. Iodine: Its Role in Thyroid Hormone Biosynthesis and Beyond. Nutrients 2021, 13, 4469. [Google Scholar] [CrossRef] [PubMed]
- Karbownik-Lewińska, M.; Stępniak, J.; Iwan, P.; Lewiński, A. Iodine as a potential endocrine disruptor-a role of oxidative stress. Endocrine 2022, 78, 219–240. [Google Scholar] [CrossRef] [PubMed]
- Byeon, Y.; Lee, K.; Park, Y.I.; Park, S.; Back, K. Molecular cloning and functional analysis of serotonin N-acetyltransferase from the cyanobacterium Synechocystis sp. PCC 6803. J. Pineal Res. 2013, 55, 371–376. [Google Scholar] [CrossRef]
- Manchester, L.C.; Coto-Montes, A.; Boga, J.A.; Andersen, L.P.; Zhou, Z.; Galano, A.; Vriend, J.; Tan, D.X.; Reiter, R.J. Melatonin: An ancient molecule that makes oxygen metabolically tolerable. J. Pineal Res. 2015, 59, 403–419. [Google Scholar] [CrossRef]
- Margulis, L. Symbiotic theory of the origin of eukaryotic organelles; criteria for proof. Symp. Soc. Exp. Biol. 1975, 29, 21–38. [Google Scholar]
- Tan, D.X.; Hardeland, R.; Manchester, L.C.; Paredes, S.D.; Korkmaz, A.; Sainz, R.M.; Mayo, J.C.; Fuentes-Broto, L.; Reiter, R.J. The changing biological roles of melatonin during evolution: From an antioxidant to signals of darkness, sexual selection and fitness. Biol. Rev. Camb. Philos. Soc. 2010, 85, 607–623. [Google Scholar] [CrossRef] [PubMed]
- Tan, D.X.; Reiter, R.J.; Zimmerman, S.; Hardeland, R. Melatonin: Both a Messenger of Darkness and a Participant in the Cellular Actions of Non-Visible Solar Radiation of Near Infrared Light. Biology 2023, 12, 89. [Google Scholar] [CrossRef]
- Zhao, D.; Yu, Y.; Shen, Y.; Liu, Q.; Zhao, Z.; Sharma, R.; Reiter, R.J. Melatonin Synthesis and Function: Evolutionary History in Animals and Plants. Front. Endocrinol. 2019, 10, 249. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Mayo, J.C.; Tan, D.X.; Sainz, R.M.; Alatorre-Jimenez, M.; Qin, L. Melatonin as an antioxidant: Under promises but over delivers. J. Pineal Res. 2016, 61, 253–278. [Google Scholar] [CrossRef]
- Tan, D.X.; Hardeland, R.; Manchester, L.C.; Galano, A.; Reiter, R.J. Cyclic-3-hydroxymelatonin (C3HOM), a potent antioxidant, scavenges free radicals and suppresses oxidative reactions. Curr. Med. Chem. 2014, 21, 1557–1565. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Tang, Q.; Zeng, Y. Melatonin: Potential avenue for treating iron overload disorders. Ageing Res. Rev. 2022, 81, 101717. [Google Scholar] [CrossRef]
- Galano, A.; Tan, D.X.; Reiter, R.J. Melatonin: A Versatile Protector against Oxidative DNA Damage. Molecules 2018, 23, 530. [Google Scholar] [CrossRef] [PubMed]
- Tan, D.X.; Manchester, L.C.; Reiter, R.J.; Qi, W.B.; Karbownik, M.; Calvo, J.R. Significance of melatonin in antioxidative defense system: Reactions and products. Biol. Signals Recept. 2000, 9, 137–159. [Google Scholar] [CrossRef]
- Barlow-Walden, L.R.; Reiter, R.J.; Abe, M.; Pablos, M.; Menendez-Pelaez, A.; Chen, L.D.; Poeggeler, B. Melatonin stimulates brain glutathione peroxidase activity. Neurochem. Int. 1995, 26, 497–502. [Google Scholar] [CrossRef]
- Abo Taleb, H.A.; Alghamdi, B.S. Neuroprotective Effects of Melatonin during Demyelination and Remyelination Stages in a Mouse Model of Multiple Sclerosis. J. Mol. Neurosci. 2020, 70, 386–402. [Google Scholar] [CrossRef] [PubMed]
- Fischer, T.W.; Kleszczyński, K.; Hardkop, L.H.; Kruse, N.; Zillikens, D. Melatonin enhances antioxidative enzyme gene expression (CAT, GPx, SOD), prevents their UVR-induced depletion, and protects against the formation of DNA damage (8-hydroxy-2’-deoxyguanosine) in ex vivo human skin. J. Pineal Res. 2013, 54, 303–312. [Google Scholar] [CrossRef] [PubMed]
- Laothong, U.; Pinlaor, P.; Hiraku, Y.; Boonsiri, P.; Prakobwong, S.; Khoontawad, J.; Pinlaor, S. Protective effect of melatonin against Opisthorchis viverrini-induced oxidative and nitrosative DNA damage and liver injury in hamsters. J. Pineal Res. 2010, 49, 271–282. [Google Scholar] [CrossRef] [PubMed]
- Sokolovic, D.; Djordjevic, B.; Kocic, G.; Stoimenov, T.J.; Stanojkovic, Z.; Sokolovic, D.M.; Veljkovic, A.; Ristic, G.; Despotovic, M.; Milisavljevic, D.; et al. The Effects of Melatonin on Oxidative Stress Parameters and DNA Fragmentation in Testicular Tissue of Rats Exposed to Microwave Radiation. Adv. Clin. Exp. Med. 2015, 24, 429–436. [Google Scholar] [CrossRef] [PubMed]
- Waldhauser, F.; Weiszenbacher, G.; Frisch, H.; Zeitlhuber, U.; Waldhauser, M.; Wurtman, R.J. Fall in nocturnal serum of melatonin during puberty and pubescence. Lancet 1984, 1, 362–365. [Google Scholar] [CrossRef]
- Sack, R.L.; Lewy, A.J.; Erb, D.L.; Vollmer, W.M.; Singer, C.M. Human melatonin production decreases with age. J. Pineal Res. 1986, 3, 379–388. [Google Scholar] [CrossRef] [PubMed]
- Sae-Teaw, M.; Johns, J.; Johns, N.P.; Subongkot, S. Serum melatonin levels and antioxidant capacities after consumption of pineapple, orange, or banana by healthy male volunteers. J. Pineal Res. 2013, 55, 58–64. [Google Scholar] [CrossRef] [PubMed]
- Esteban, S.; Nicolaus, C.; Garmundi, A.; Rial, R.V.; Rodríguez, A.B.; Ortega, E.; Ibars, C.B. Effect of orally administered l-tryptophan on serotonin, melatonin, and the innate immune response in the rat. Mol. Cell. Biochem. 2004, 267, 39–46. [Google Scholar] [CrossRef]
- Li, J.; Somers, V.K.; Xu, H.; Lopez-Jimenez, F.; Covassin, N. Trends in Use of Melatonin Supplements Among US Adults, 1999–2018. JAMA 2022, 327, 483–485. [Google Scholar] [CrossRef]
- Morsali, S.; Sabahi, Z.; Kakaei, J.; Hakimzadeh, Z.; Hamidi, S.; Gholipour-Khalili, E.; Sanaie, S.; Talebi, M.; Naseri, A. Clinical efficacy and safety of melatonin supplementation in multiple sclerosis: A systematic review. Inflammopharmacology 2023, 31, 2213–2220. [Google Scholar] [CrossRef]
- Minich, D.M.; Henning, M.; Darley, C.; Fahoum, M.; Schuler, C.B.; Frame, J. Is Melatonin the “Next Vitamin D”?: A Review of Emerging Science, Clinical Uses, Safety, and Dietary Supplements. Nutrients 2022, 14, 3934. [Google Scholar] [CrossRef] [PubMed]
- Andersen, L.P.; Gögenur, I.; Rosenberg, J.; Reiter, R.J. The Safety of Melatonin in Humans. Clin. Drug Investig. 2016, 36, 169–175. [Google Scholar] [CrossRef] [PubMed]
- Zetner, D.; Andersen, L.P.K.; Alder, R.; Jessen, M.L.; Tolstrup, A.; Rosenberg, J. Pharmacokinetics and Safety of Intravenous, Intravesical, Rectal, Transdermal, and Vaginal Melatonin in Healthy Female Volunteers: A Cross-Over Study. Pharmacology 2021, 106, 169–176. [Google Scholar] [CrossRef]
- Lewinski, A.; Karbownik, M. REVIEW. Melatonin and the thyroid gland. Neuro Endocrinol. Lett. 2002, 23 (Suppl. S1), 73–78. [Google Scholar] [PubMed]
- Wood, S.; Loudon, A. Clocks for all seasons: Unwinding the roles and mechanisms of circadian and interval timers in the hypothalamus and pituitary. J. Endocrinol. 2014, 222, R39–R59. [Google Scholar] [CrossRef] [PubMed]
- Yasuo, S.; Nakao, N.; Ohkura, S.; Iigo, M.; Hagiwara, S.; Goto, A.; Ando, H.; Yamamura, T.; Watanabe, M.; Watanabe, T.; et al. Long-day suppressed expression of type 2 deiodinase gene in the mediobasal hypothalamus of the Saanen goat, a short-day breeder: Implication for seasonal window of thyroid hormone action on reproductive neuroendocrine axis. Endocrinology 2006, 147, 432–440. [Google Scholar] [CrossRef] [PubMed]
- Ertek, S. Molecular economy of nature with two thyrotropins from different parts of the pituitary: Pars tuberalis thyroid-stimulating hormone and pars distalis thyroid-stimulating hormone. Arch. Med. Sci. 2021, 17, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Freeman, D.A.; Teubner, B.J.; Smith, C.D.; Prendergast, B.J. Exogenous T3 mimics long day lengths in Siberian hamsters. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2007, 292, R2368–R2372. [Google Scholar] [CrossRef] [PubMed]
- Hanon, E.A.; Lincoln, G.A.; Fustin, J.M.; Dardente, H.; Masson-Pévet, M.; Morgan, P.J.; Hazlerigg, D.G. Ancestral TSH mechanism signals summer in a photoperiodic mammal. Curr. Biol. 2008, 18, 1147–1152. [Google Scholar] [CrossRef]
- Garcia-Marin, R.; de Miguel, M.; Fernández-Santos, J.M.; Carrillo-Vico, A.; Utrilla, J.C.; Morillo-Bernal, J.; Díaz-Parrado, E.; Rodríguez-Prieto, I.; Guerrero, J.M.; Martín-Lacave, I. Melatonin-synthesizing enzymes and melatonin receptor in rat thyroid cells. Histol. Histopathol. 2012, 27, 1429–1438. [Google Scholar] [CrossRef]
- Garcia-Marin, R.; Fernandez-Santos, J.M.; Morillo-Bernal, J.; Gordillo-Martinez, F.; Vazquez-Roman, V.; Utrilla, J.C.; Carrillo-Vico, A.; Guerrero, J.M.; Martin-Lacave, I. Melatonin in the thyroid gland: Regulation by thyroid-stimulating hormone and role in thyroglobulin gene expression. J. Physiol. Pharmacol. 2015, 66, 643–652. [Google Scholar]
- Taheri, P.; Mogheiseh, A.; Shojaee Tabrizi, A.; Nazifi, S.; Salavati, S.; Koohi, F. Changes in thyroid hormones, leptin, ghrelin and, galanin following oral melatonin administration in intact and castrated dogs: A preliminary study. BMC Vet. Res. 2019, 15, 145. [Google Scholar] [CrossRef] [PubMed]
- Mogulkoc, R.; Baltaci, A.K. The effect of intraperitoneal melatonin supplementation on the release of thyroid hormones and testosterone in rats with hyperthyroid. Neuro Endocrinol. Lett. 2003, 24, 345–347. [Google Scholar] [PubMed]
- Belviranli, M.; Baltaci, A.K. The relation between reduced serum melatonin levels and zinc in rats with induced hypothyroidism. Cell Biochem. Funct. 2008, 26, 19–23. [Google Scholar] [CrossRef] [PubMed]
- Cini, G.; Neri, B.; Pacini, A.; Cesati, V.; Sassoli, C.; Quattrone, S.; D’Apolito, M.; Fazio, A.; Scapagnini, G.; Provenzani, A.; et al. Antiproliferative activity of melatonin by transcriptional inhibition of cyclin D1 expression: A molecular basis for melatonin-induced oncostatic effects. J. Pineal Res. 2005, 39, 12–20. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Liu, D.; Di, S.; Zhang, Z.; Li, W.; Zhang, J.; Xu, L.; Guo, K.; Zhu, Y.; Li, X.; et al. Histone deacetylase 9 downregulation decreases tumor growth and promotes apoptosis in non-small cell lung cancer after melatonin treatment. J. Pineal Res. 2019, 67, e12587. [Google Scholar] [CrossRef] [PubMed]
- Carbajo-Pescador, S.; Ordoñez, R.; Benet, M.; Jover, R.; García-Palomo, A.; Mauriz, J.L.; González-Gallego, J. Inhibition of VEGF expression through blockade of Hif1α and STAT3 signalling mediates the anti-angiogenic effect of melatonin in HepG2 liver cancer cells. Br. J. Cancer 2013, 109, 83–91. [Google Scholar] [CrossRef] [PubMed]
- El-Sokkary, G.H.; Ismail, I.A.; Saber, S.H. Melatonin inhibits breast cancer cell invasion through modulating DJ-1/KLF17/ID-1 signaling pathway. J. Cell. Biochem. 2019, 120, 3945–3957. [Google Scholar] [CrossRef] [PubMed]
- Chao, C.C.; Chen, P.C.; Chiou, P.C.; Hsu, C.J.; Liu, P.I.; Yang, Y.C.; Reiter, R.J.; Yang, S.F.; Tang, C.H. Melatonin suppresses lung cancer metastasis by inhibition of epithelial-mesenchymal transition through targeting to Twist. Clin. Sci. 2019, 133, 709–722. [Google Scholar] [CrossRef]
- Gurunathan, S.; Qasim, M.; Kang, M.H.; Kim, J.H. Role and Therapeutic Potential of Melatonin in Various Type of Cancers. OncoTargets Ther. 2021, 14, 2019–2052. [Google Scholar] [CrossRef]
- Reiter, R.J.; Rosales-Corral, S.A.; Tan, D.X.; Acuna-Castroviejo, D.; Qin, L.; Yang, S.F.; Xu, K. Melatonin, a Full Service Anti-Cancer Agent: Inhibition of Initiation, Progression and Metastasis. Int. J. Mol. Sci. 2017, 18, 843. [Google Scholar] [CrossRef] [PubMed]
- Mafi, A.; Rezaee, M.; Hedayati, N.; Hogan, S.D.; Reiter, R.J.; Aarabi, M.H.; Asemi, Z. Melatonin and 5-fluorouracil combination chemotherapy: Opportunities and efficacy in cancer therapy. Cell Commun. Signal. 2023, 21, 33. [Google Scholar] [CrossRef]
- Lissoni, P. Biochemotherapy with standard chemotherapies plus the pineal hormone melatonin in the treatment of advanced solid neoplasms. Pathol. Biol. 2007, 55, 201–204. [Google Scholar] [CrossRef] [PubMed]
- Kartini, D.; Taher, A.; Panigoro, S.S.; Setiabudy, R.; Jusman, S.W.; Haryana, S.M.; Murdani, A.; Rustamadji, P.; Karisyah, A.; Rasyid, S.H. Melatonin effect on hypoxia inducible factor-1α and clinical response in patients with oral squamous cell carcinoma receiving neoadjuvant chemotherapy: A randomized controlled trial. J. Carcinog. 2021, 20, 5. [Google Scholar] [CrossRef] [PubMed]
- Palmer, A.C.S.; Zortea, M.; Souza, A.; Santos, V.; Biazús, J.V.; Torres, I.L.S.; Fregni, F.; Caumo, W. Clinical impact of melatonin on breast cancer patients undergoing chemotherapy; effects on cognition, sleep and depressive symptoms: A randomized, double-blind, placebo-controlled trial. PLoS ONE 2020, 15, e0231379. [Google Scholar] [CrossRef] [PubMed]
- Stępniak, J.; Krawczyk-Lipiec, J.; Lewiński, A.; Karbownik-Lewińska, M. Sorafenib versus Lenvatinib Causes Stronger Oxidative Damage to Membrane Lipids in Noncancerous Tissues of the Thyroid, Liver, and Kidney: Effective Protection by Melatonin and Indole-3-Propionic Acid. Biomedicines 2022, 10, 2890. [Google Scholar] [CrossRef] [PubMed]
- Ma, W.; Liu, M.; Liang, F.; Zhao, L.; Gao, C.; Jiang, X.; Zhang, X.; Zhan, H.; Hu, H.; Zhao, Z. Cardiotoxicity of sorafenib is mediated through elevation of ROS level and CaMKII activity and dysregulation of calcium homoeostasis. Basic. Clin. Pharmacol. Toxicol. 2020, 126, 166–180. [Google Scholar] [CrossRef] [PubMed]
- Aras, S.; Tanzer, I.H.O.; Karaçavuş, S.; Sayir, N.; Erdem, E.; Hacımustafaoğlu, F.; Erdoğan, C.E.; Sapmaz, T.; İkizceli, T.; Pençe, H.H.; et al. Effect of melatonin on low and high dose radiotherapy induced thyroid injury. Biotech. Histochem. 2023, 98, 346–352. [Google Scholar] [CrossRef] [PubMed]
- Arıcıgil, M.; Dündar, M.A.; Yücel, A.; Eryılmaz, M.A.; Aktan, M.; Alan, M.A.; Fındık, S.; Kılınç, İ. Melatonin prevents possible radiotherapy-induced thyroid injury. Int. J. Radiat. Biol. 2017, 93, 1350–1356. [Google Scholar] [CrossRef]
- Zou, Z.W.; Liu, T.; Li, Y.; Chen, P.; Peng, X.; Ma, C.; Zhang, W.J.; Li, P.D. Melatonin suppresses thyroid cancer growth and overcomes radioresistance via inhibition of p65 phosphorylation and induction of ROS. Redox Biol. 2018, 16, 226–236. [Google Scholar] [CrossRef]
- Ghorbani-Anarkooli, M.; Dabirian, S.; Zendedel, A.; Moladoust, H.; Bahadori, M.H. Effects of melatonin on the toxicity and proliferation of human anaplastic thyroid cancer cell line. Acta Histochem. 2021, 123, 151700. [Google Scholar] [CrossRef] [PubMed]
- Jia, H.; Sun, W.; Li, X.; Xu, W. Melatonin promotes apoptosis of thyroid cancer cells via regulating the signaling of microRNA-21 (miR-21) and microRNA-30e (miR-30e). Bioengineered 2022, 13, 9588–9601. [Google Scholar] [CrossRef] [PubMed]
- Iwan, P.; Stepniak, J.; Karbownik-Lewinska, M. Cumulative Protective Effect of Melatonin and Indole-3-Propionic Acid against KIO3-Induced Lipid Peroxidation in Porcine Thyroid. Toxics 2021, 9, 89. [Google Scholar] [CrossRef] [PubMed]
- Iwan, P.; Stepniak, J.; Karbownik-Lewinska, M. Melatonin reduces high levels of lipid peroxidation induced by potassium iodate in porcine thyroid. Int. J. Vitam. Nutr. Res. 2021, 91, 271–277. [Google Scholar] [CrossRef] [PubMed]
- Stasiak, M.; Zasada, K.; Lewinski, A.; Karbownik-Lewinska, M. Melatonin restores the basal level of lipid peroxidation in rat tissues exposed to potassium bromate in vitro. Neuro Endocrinol. Lett. 2010, 31, 363–369. [Google Scholar] [PubMed]
- Karbownik, M.; Stasiak, M.; Zygmunt, A.; Zasada, K.; Lewiński, A. Protective effects of melatonin and indole-3-propionic acid against lipid peroxidation, caused by potassium bromate in the rat kidney. Cell Biochem. Funct. 2006, 24, 483–489. [Google Scholar] [CrossRef] [PubMed]
- IARC. Arsenic, Metals, Fibres, and Dusts. In Volume 100C: A Review of Human Carcinogens; IARC: Lyon, France, 2012. [Google Scholar]
- Deng, Y.; Wang, M.; Tian, T.; Lin, S.; Xu, P.; Zhou, L.; Dai, C.; Hao, Q.; Wu, Y.; Zhai, Z.; et al. The Effect of Hexavalent Chromium on the Incidence and Mortality of Human Cancers: A Meta-Analysis Based on Published Epidemiological Cohort Studies. Front. Oncol. 2019, 9, 24. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Islam, R.; Wang, Y.; Zhang, X.; Liu, L.Z. Epigenetic Regulation in Chromium-, Nickel- and Cadmium-Induced Carcinogenesis. Cancers 2022, 14, 5768. [Google Scholar] [CrossRef] [PubMed]
- DesMarais, T.L.; Costa, M. Mechanisms of Chromium-Induced Toxicity. Curr. Opin. Toxicol. 2019, 14, 1–7. [Google Scholar] [CrossRef]
- Saeed, E.; El-Mansy, A.A.; Mazroa, S.A.; Moustafa, A.M. The possible protective role of vitamin C versus melatonin on potassium dichromate induced changes in thyroid gland: Light and electron microscopic study. Ultrastruct. Pathol. 2023, 47, 73–89. [Google Scholar] [CrossRef]
- IARC. Ionizing Radiation. In Part 1: X- and Gamma (γ)-Radiation, and Neutrons Volume 75: IARC Monographs on the Evaluation of Carcinogenic Risks to Humans; IARC: Lyon, France, 2000. [Google Scholar]
- Rooney, M.K.; Andring, L.M.; Corrigan, K.L.; Bernard, V.; Williamson, T.D.; Fuller, C.D.; Garden, A.S.; Gunn, B.; Lee, A.; Moreno, A.C.; et al. Hypothyroidism following Radiotherapy for Head and Neck Cancer: A Systematic Review of the Literature and Opportunities to Improve the Therapeutic Ratio. Cancers 2023, 15, 4321. [Google Scholar] [CrossRef]
- Brook, I. Late side effects of radiation treatment for head and neck cancer. Radiat. Oncol. J. 2020, 38, 84–92. [Google Scholar] [CrossRef]
- Karbownik, M.; Reiter, R.J. Antioxidative effects of melatonin in protection against cellular damage caused by ionizing radiation. Proc. Soc. Exp. Biol. Med. 2000, 225, 9–22. [Google Scholar] [CrossRef]
- IARC. Ionizing Radiation. In Part 2: Some Internally Deposited Radionuclides Volume 78: IARC Monographs on the Evaluation of Carcinogenic Risks to Humans; IARC: Lyon, France, 2001. [Google Scholar]
- Shim, S.R.; Kitahara, C.M.; Cha, E.S.; Kim, S.J.; Bang, Y.J.; Lee, W.J. Cancer Risk After Radioactive Iodine Treatment for Hyperthyroidism: A Systematic Review and Meta-analysis. JAMA Netw. Open 2021, 4, e2125072. [Google Scholar] [CrossRef]
- Pasqual, E.; Schonfeld, S.; Morton, L.M.; Villoing, D.; Lee, C.; Berrington de Gonzalez, A.; Kitahara, C.M. Association Between Radioactive Iodine Treatment for Pediatric and Young Adulthood Differentiated Thyroid Cancer and Risk of Second Primary Malignancies. J. Clin. Oncol. 2022, 40, 1439–1449. [Google Scholar] [CrossRef]
- Cebi Sen, C.; Yumusak, N.; Atilgan, H.I.; Sadic, M.; Koca, G.; Korkmaz, M. The protective effect of melatonin on sperm quality in rat after radioiodine treatment. Andrologia 2018, 50, e12962. [Google Scholar] [CrossRef]
- Barlas, A.M.; Sadic, M.; Atilgan, H.I.; Bag, Y.M.; Onalan, A.K.; Yumusak, N.; Senes, M.; Fidanci, V.; Pekcici, M.R.; Korkmaz, M.; et al. Melatonin: A hepatoprotective agent against radioiodine toxicity in rats. Bratisl. Med. J. 2017, 118, 95–100. [Google Scholar] [CrossRef]
- Alidadi, S.; Shabestani Monfared, A.; Amiri, M.; Zabihi, E.; Assadollahi, E.; Gholami, A.; Moazezi, Z.; Abedian, Z. The efficacy of melatonin against radiotoxicity of iodine-131 and its response to treatment in hyperthyroid patients: A randomized controlled trial. Nucl. Med. Rev. Cent. East. Eur. 2022, 25, 31–36. [Google Scholar] [CrossRef]
- IARC Monographs Vol 124 Group. Carcinogenicity of night shift work. Lancet Oncol. 2019, 20, 1058–1059. [Google Scholar] [CrossRef]
- Erren, T.C.; Morfeld, P.; Groß, J.V.; Wild, U.; Lewis, P. IARC 2019: “Night shift work” is probably carcinogenic: What about disturbed chronobiology in all walks of life? J. Occup. Med. Toxicol. 2019, 14, 29. [Google Scholar] [CrossRef]
- Papantoniou, K.; Konrad, P.; Haghayegh, S.; Strohmaier, S.; Eliassen, A.H.; Schernhammer, E. Rotating Night Shift Work, Sleep, and Thyroid Cancer Risk in the Nurses’ Health Study 2. Cancers 2023, 15, 5673. [Google Scholar] [CrossRef]
- Zhang, D.; Jones, R.R.; James, P.; Kitahara, C.M.; Xiao, Q. Associations between artificial light at night and risk for thyroid cancer: A large US cohort study. Cancer 2021, 127, 1448–1458. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J.; Tan, D.X.; Korkmaz, A.; Erren, T.C.; Piekarski, C.; Tamura, H.; Manchester, L.C. Light at night, chronodisruption, melatonin suppression, and cancer risk: A review. Crit. Rev. Oncog. 2007, 13, 303–328. [Google Scholar] [CrossRef]
- IARC Monographs Vol 123 Group. Carcinogenicity of some nitrobenzenes and other industrial chemicals. Lancet Oncol. 2018, 19, e681–e682. [Google Scholar] [CrossRef]
- Hsu, C.H.; Stedeford, T.; Okochi-Takada, E.; Ushijima, T.; Noguchi, H.; Muro-Cacho, C.; Holder, J.W.; Banasik, M. Framework analysis for the carcinogenic mode of action of nitrobenzene. J. Environ. Sci. Health C Environ. Carcinog. Ecotoxicol. Rev. 2007, 25, 155–184. [Google Scholar] [CrossRef]
- U.S. Environmental Protection Agency. Integrated Risk Information System (IRIS) on Nitrobenzene; National Center for Environmental Assessment, Office of Research and Development: Washington, DC, USA, 1999.
- Cattley, R.C.; Everitt, J.I.; Gross, E.A.; Moss, O.R.; Hamm, T.E., Jr.; Popp, J.A. Carcinogenicity and toxicity of inhaled nitrobenzene in B6C3F1 mice and F344 and CD rats. Fundam. Appl. Toxicol. 1994, 22, 328–340. [Google Scholar] [CrossRef]
- Zasada, K.; Karbownik-Lewinska, M. Comparison of potential protective effects of melatonin and propylthiouracil against lipid peroxidation caused by nitrobenzene in the thyroid gland. Toxicol. Ind. Health 2015, 31, 1195–1201. [Google Scholar] [CrossRef]
- IARC. Some Chemicals that Cause Tumours of the Kidney or Urinary Bladder in Rodents and Some Other Substances. In Volume 73: IARC Monographs on the Evaluation of Carcinogenic Risks to Humans; IARC: Lyon, France, 1999. [Google Scholar]
- Watanabe, S.; Togashi, S.; Fukui, T. Contribution of nitric oxide to potassium bromate-induced elevation of methaemoglobin concentration in mouse blood. Biol. Pharm. Bull. 2002, 25, 1315–1319. [Google Scholar] [CrossRef]
- Kakehashi, A.; Wei, M.; Fukushima, S.; Wanibuchi, H. Oxidative stress in the carcinogenicity of chemical carcinogens. Cancers 2013, 5, 1332–1354. [Google Scholar] [CrossRef]
- Karbownik, M.; Stasiak, M.; Zasada, K.; Zygmunt, A.; Lewinski, A. Comparison of potential protective effects of melatonin, indole-3-propionic acid, and propylthiouracil against lipid peroxidation caused by potassium bromate in the thyroid gland. J. Cell. Biochem. 2005, 95, 131–138. [Google Scholar] [CrossRef]
- World Health Organization. Preventing Disease through Healthy Environments: Exposure to Mercury: A Major Public Health Concern, 2nd ed. Available online: https://www.who.int/publications/i/item/9789240023567 (accessed on 10 January 2024).
- Skalny, A.V.; Aschner, M.; Sekacheva, M.I.; Santamaria, A.; Barbosa, F.; Ferrer, B.; Aaseth, J.; Paoliello, M.M.B.; Rocha, J.B.T.; Tinkov, A.A. Mercury and cancer: Where are we now after two decades of research? Food Chem. Toxicol. 2022, 164, 113001. [Google Scholar] [CrossRef]
- IARC. Beryllium, Cadmium, Mercury, and Exposures in the Glass Manufacturing Industry. In Volume 58: IARC Monographs on the Evaluation of Carcinogenic Risks to Humans; IARC: Lyon, France, 1993. [Google Scholar]
- U.S. Environmental Protection Agency. Integrated Risk Information System (IRIS) on Mercury; National Center for Environmental Assessment, Office of Research and Development: Washington, DC, USA, 2000.
- Zhu, X.; Kusaka, Y.; Sato, K.; Zhang, Q. The endocrine disruptive effects of mercury. Environ. Health Prev. Med. 2000, 4, 174–183. [Google Scholar] [CrossRef]
- Pamphlett, R.; Doble, P.A.; Bishop, D.P. Mercury in the human thyroid gland: Potential implications for thyroid cancer, autoimmune thyroiditis, and hypothyroidism. PLoS ONE 2021, 16, e0246748. [Google Scholar] [CrossRef]
- Buonaurio, F.; Astolfi, M.L.; Pigini, D.; Tranfo, G.; Canepari, S.; Pietroiusti, A.; D’Alessandro, I.; Sisto, R. Oxidative Stress Biomarkers in Urine of Metal Carpentry Workers Can Be Diagnostic for Occupational Exposure to Low Level of Welding Fumes from Associated Metals. Cancers 2021, 13, 3167. [Google Scholar] [CrossRef]
- Webster, A.M.; Pinion, D.; Pineda, E.; Aboueisha, H.; Hussein, M.H.; Fawzy, M.S.; Toraih, E.A.; Kandil, E. Elucidating the link between thyroid cancer and mercury exposure: A review and meta-analysis. Environ. Sci. Pollut. Res. Int. 2024, 31, 12841–12855. [Google Scholar] [CrossRef]
- Rao, M.V.; Chhunchha, B. Protective role of melatonin against the mercury induced oxidative stress in the rat thyroid. Food Chem. Toxicol. 2010, 48, 7–10. [Google Scholar] [CrossRef]
- Lisco, G.; De Tullio, A.; Triggiani, D.; Zupo, R.; Giagulli, V.A.; De Pergola, G.; Piazzolla, G.; Guastamacchia, E.; Sabbà, C.; Triggiani, V. Iodine Deficiency and Iodine Prophylaxis: An Overview and Update. Nutrients 2023, 15, 1004. [Google Scholar] [CrossRef]
- Poul, J.M.; Huet, S.; Godard, T.; Sanders, P. Lack of genotoxicity of potassium iodate in the alkaline comet assay and in the cytokinesis-block micronucleus test. Comparison to potassium bromate. Food Chem. Toxicol. 2004, 42, 203–209. [Google Scholar] [CrossRef]
- Milczarek, M.; Stępniak, J.; Lewiński, A.; Karbownik-Lewińska, M. Potassium iodide, but not potassium iodate, as a potential protective agent against oxidative damage to membrane lipids in porcine thyroid. Thyroid. Res. 2013, 6, 10. [Google Scholar] [CrossRef]
- Toyokuni, S.; Kong, Y.; Cheng, Z.; Sato, K.; Hayashi, S.; Ito, F.; Jiang, L.; Yanatori, I.; Okazaki, Y.; Akatsuka, S. Carcinogenesis as Side Effects of Iron and Oxygen Utilization: From the Unveiled Truth toward Ultimate Bioengineering. Cancers 2020, 12, 3320. [Google Scholar] [CrossRef]
- Toyokuni, S. Role of iron in carcinogenesis: Cancer as a ferrotoxic disease. Cancer Sci. 2009, 100, 9–16. [Google Scholar] [CrossRef]
- Fonseca-Nunes, A.; Jakszyn, P.; Agudo, A. Iron and cancer risk--a systematic review and meta-analysis of the epidemiological evidence. Cancer Epidemiol. Biomark. Prev. 2014, 23, 12–31. [Google Scholar] [CrossRef]
- Zoeller, R.T.; Brown, T.R.; Doan, L.L.; Gore, A.C.; Skakkebaek, N.E.; Soto, A.M.; Woodruff, T.J.; Vom Saal, F.S. Endocrine-disrupting chemicals and public health protection: A statement of principles from The Endocrine Society. Endocrinology 2012, 153, 4097–4110. [Google Scholar] [CrossRef]
- Calsolaro, V.; Pasqualetti, G.; Niccolai, F.; Caraccio, N.; Monzani, F. Thyroid Disrupting Chemicals. Int. J. Mol. Sci. 2017, 18, 2583. [Google Scholar] [CrossRef]
- Gładysz, A.K.; Stępniak, J.; Karbownik-Lewińska, M. Exogenous Melatonin Protects against Oxidative Damage to Membrane Lipids Caused by Some Sodium/Iodide Symporter Inhibitors in the Thyroid. Antioxidants 2023, 12, 1688. [Google Scholar] [CrossRef]
- Sarkar, D.; Chandra, A.; Chattopadyay, S.; Biswas, M.; Das, S.; Singh, L.; Ray, I. Possible mechanism of bamboo shoots (Bambusa balcooa) induced thyroid disruption—An in vitro study. Hum. Exp. Toxicol. 2021, 40, 483–496. [Google Scholar] [CrossRef]
- Adedara, I.A.; Ojuade, T.J.D.; Olabiyi, B.F.; Idris, U.F.; Onibiyo, E.M.; Ajeigbe, O.F.; Farombi, E.O. Taurine Ameliorates Renal Oxidative Damage and Thyroid Dysfunction in Rats Chronically Exposed to Fluoride. Biol. Trace Elem. Res. 2017, 175, 388–395. [Google Scholar] [CrossRef]
- Akinrinde, A.S.; Tijani, M.; Awodele, O.A.; Oyagbemi, A.A. Fluoride-induced hepatotoxicity is prevented by L-Arginine supplementation via suppression of oxidative stress and stimulation of nitric oxide production in rats. Toxicol. Environ. Health Sci. 2021, 13, 57–64. [Google Scholar] [CrossRef]
- Kitahara, J.; Seko, Y.; Imura, N. Possible involvement of active oxygen species in selenite toxicity in isolated rat hepatocytes. Arch. Toxicol. 1993, 67, 497–501. [Google Scholar] [CrossRef]
- Hariharan, S.; Dharmaraj, S. Selenium and selenoproteins: It’s role in regulation of inflammation. Inflammopharmacology 2020, 28, 667–695. [Google Scholar] [CrossRef]
- Ali, S.N.; Arif, A.; Ansari, F.A.; Mahmood, R. Cytoprotective effect of taurine against sodium chlorate-induced oxidative damage in human red blood cells: An ex vivo study. Amino Acids 2022, 54, 33–46. [Google Scholar] [CrossRef] [PubMed]
- Sirasanagandla, S.R.; Al-Huseini, I.; Sakr, H.; Moqadass, M.; Das, S.; Juliana, N.; Abu, I.F. Natural Products in Mitigation of Bisphenol A Toxicity: Future Therapeutic Use. Molecules 2022, 27, 5384. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.G.; Correia, J.; Adiga, D.; Rai, P.S.; Dsouza, H.S.; Chakrabarty, S.; Kabekkodu, S.P. A comprehensive review on the carcinogenic potential of bisphenol A: Clues and evidence. Environ. Sci. Pollut. Res. Int. 2021, 28, 19643–19663. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.M.D.; Xavier, L.L.F.; Gonçalves, C.F.L.; Santos-Silva, A.P.; Paiva-Melo, F.D.; Freitas, M.L.; Fortunato, R.S.; Alves, L.M.; Ferreira, A.C.F. Bisphenol A increases hydrogen peroxide generation by thyrocytes both in vivo and in vitro. Endocr. Connect. 2018, 7, 1196–1207. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Oxidative nature of the thyroid follicular cell (thyrocyte). Only documented reactive oxygen species (ROS) or free radicals, present inside the thyroid follicular cell, are marked in the scheme. Fe2+, ferrous ion; Fe3+, ferric ion; H2O2, hydrogen peroxide; O2, molecular oxygen; O2•−, superoxide anion radical; •OH, hydroxyl radical; OH−, hydroxide ion; TPO, thyroid peroxidase; DUOX2, dual oxidase 2; NOX4, NADPH oxidase 4; IO−, hypoiodite anion; I−, iodide ion; NIS, sodium/iodide symporter; Na+, sodium ion.
Figure 1.
Oxidative nature of the thyroid follicular cell (thyrocyte). Only documented reactive oxygen species (ROS) or free radicals, present inside the thyroid follicular cell, are marked in the scheme. Fe2+, ferrous ion; Fe3+, ferric ion; H2O2, hydrogen peroxide; O2, molecular oxygen; O2•−, superoxide anion radical; •OH, hydroxyl radical; OH−, hydroxide ion; TPO, thyroid peroxidase; DUOX2, dual oxidase 2; NOX4, NADPH oxidase 4; IO−, hypoiodite anion; I−, iodide ion; NIS, sodium/iodide symporter; Na+, sodium ion.

Figure 2.
Antioxidative actions of melatonin. •OH, hydroxyl radical; RO•, alkoxy radical; ROO•, peroxy radical; •NO, nitric oxide radical; H2O2, hydrogen peroxide; 1O2, singlet oxygen; ONOO−, peroxynitrite anion; SOD, superoxide dismutase; GPx, glutathione peroxidase; GR, glutathione reductase; CAT, catalase; XO, xanthine oxidase; iNOS, inducible nitric oxide synthase; Fe, iron.
Figure 2.
Antioxidative actions of melatonin. •OH, hydroxyl radical; RO•, alkoxy radical; ROO•, peroxy radical; •NO, nitric oxide radical; H2O2, hydrogen peroxide; 1O2, singlet oxygen; ONOO−, peroxynitrite anion; SOD, superoxide dismutase; GPx, glutathione peroxidase; GR, glutathione reductase; CAT, catalase; XO, xanthine oxidase; iNOS, inducible nitric oxide synthase; Fe, iron.

Figure 3.
The relationship between the thyroid and melatonin documented in experimental studies. T4, thyroxine; T3, triiodothyronine.
Figure 3.
The relationship between the thyroid and melatonin documented in experimental studies. T4, thyroxine; T3, triiodothyronine.

Figure 4.
The mechanisms of the potential protective effects of melatonin against the process of carcinogenesis.
Figure 4.
The mechanisms of the potential protective effects of melatonin against the process of carcinogenesis.

Figure 5.
Protective effects of melatonin against documented/potential carcinogens in the thyroid. LPO, lipid peroxidation. Actions of particular documented/potential carcinogens (as well as adequate references) are presented in Table 1 and Table 2.

Table 1.
Protective effects of melatonin against oxidative damage caused by documented carcinogens (listed in the International Agency for Research on Cancer (IARC) Monographs). MDA, malondialdehyde; MDA + 4-HDA, malondialdehyde + 4-hydroxyalkenals; SOD, superoxide dismutase; GPx, glutathione peroxidase; GR, glutathione reductase; CAT, catalase; GSH, glutathione; ip, intraperitoneally; ↑, increase; ↓, decrease.
Table 1.
Protective effects of melatonin against oxidative damage caused by documented carcinogens (listed in the International Agency for Research on Cancer (IARC) Monographs). MDA, malondialdehyde; MDA + 4-HDA, malondialdehyde + 4-hydroxyalkenals; SOD, superoxide dismutase; GPx, glutathione peroxidase; GR, glutathione reductase; CAT, catalase; GSH, glutathione; ip, intraperitoneally; ↑, increase; ↓, decrease.
| IARC Group | Agent/Dose | Species/Organ/ Tissue/Cellular Compartment | Effect of Carcinogen | Dose of Melatonin, Which Reduced or Prevented the Effect of Carcinogen | Refs. |
|---|---|---|---|---|---|
| 1 | Hexavalent chromium/potassium dichromate 25 mg/kg/day for 2 months | Adult male albino rats of Wistar strain | Follicular hyperplasia, follicular enlargement | 10 mg/kg/day for 2 months | [92] |
| 1 | Radiotherapy/ionizing radiation total dose of 16–18 Gy | Adult female rats | Increased inflammation, vacuolization, degradation, swelling, and necrosis in the thyroid gland | 10–50 mg/kg 10–15 min before exposure | [79,80] |
| 1 | Radioiodinetherapy /iodine-131 111 MBq | Adult rats | Oxidative stress parameters in the liver | 12 mg/kg/day | [101] |
| Radioiodinetherapy /iodine-131 370 to 740 MBq | Hyperthyroid patients | Chromosomal damage in lymphocyte | 300 mg | [102] | |
| 2A | Night-shift work | Humans | Indirect evidence on carcinogenesis | Not documented in the literature | - |
| 2B | Nitrobenzene 7.5–10.0 mM | Porcine thyroid homogenates | ↑MDA + 4-HDA in the thyroid | 0.0001 mM | [112] |
| 2B | Bromium/potassium bromate 110 mg/kg | Adult rats | ↑MDA + 4-HDA in the thyroid | 15 mg/kg twice daily for 10 days | [116] |
| 3 | Mercury/mercury chloride 2 and 4 mg/kg, orally | Adult male albino rats of Wistar strain | ↑MDA ↓SOD, ↓CAT, ↓GPx, ↓GR, ↓GSH | 5 mg/kg, ip | [125] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Stępniak, J.; Karbownik-Lewińska, M. Protective Effects of Melatonin against Carcinogen-Induced Oxidative Damage in the Thyroid. Cancers 2024, 16, 1646. https://doi.org/10.3390/cancers16091646
AMA Style
Stępniak J, Karbownik-Lewińska M. Protective Effects of Melatonin against Carcinogen-Induced Oxidative Damage in the Thyroid. Cancers. 2024; 16(9):1646. https://doi.org/10.3390/cancers16091646
Chicago/Turabian StyleStępniak, Jan, and Małgorzata Karbownik-Lewińska. 2024. "Protective Effects of Melatonin against Carcinogen-Induced Oxidative Damage in the Thyroid" Cancers 16, no. 9: 1646. https://doi.org/10.3390/cancers16091646
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.