
Evaluation and Optimization of the Anti-Melanogenic Activity of 1-(2-Cyclohexylmethoxy-6-hydroxy-phenyl)-3-(4-hydroxymethyl-phenyl)-propenone Derivatives


Abstract
:1. Introduction
2. Results and Discussion
2.1. Chemistry
2.2. Biological Evaluation
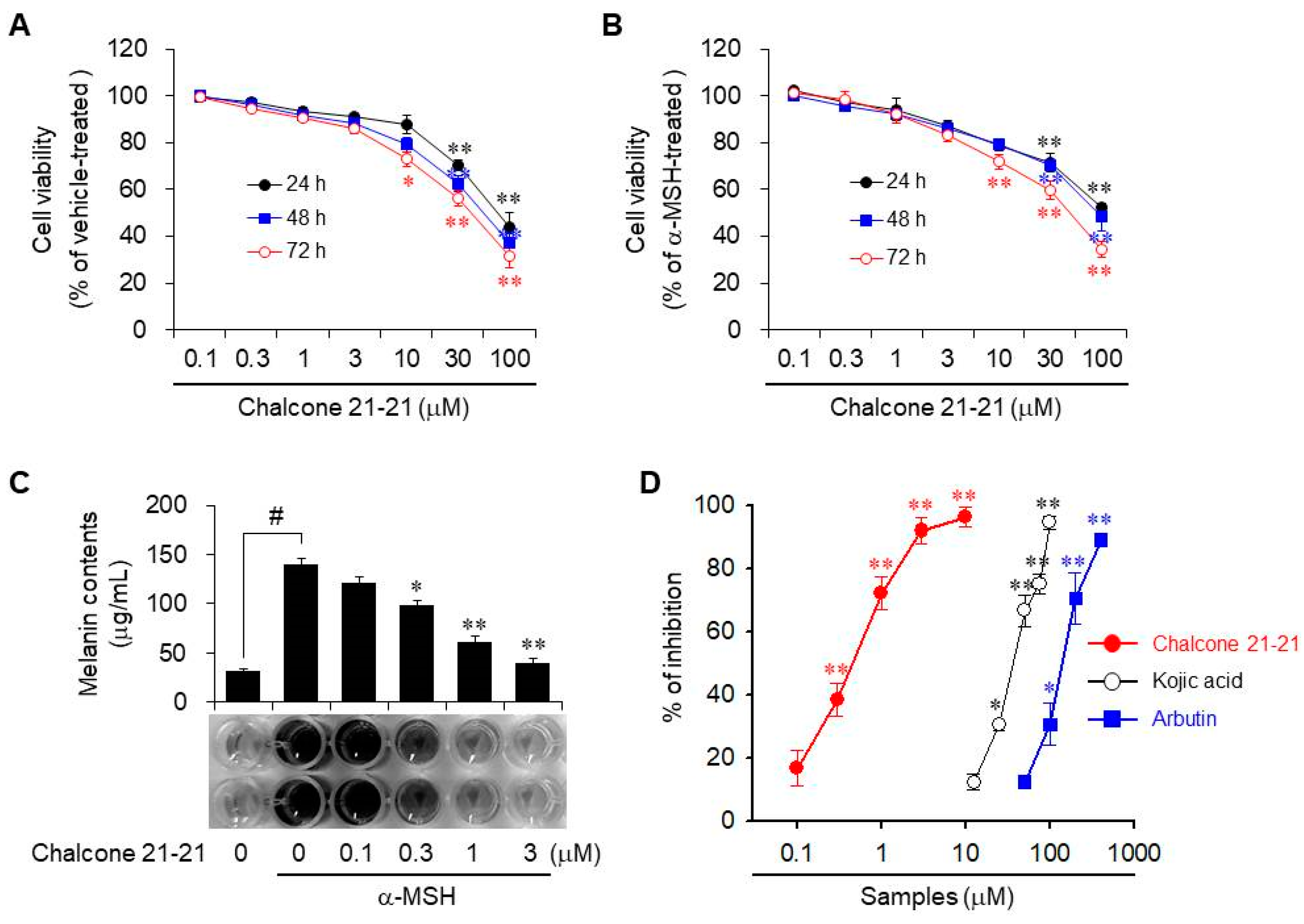
2.2.1. Cellular Melanin Production
2.2.2. Anti-Melanogenic Activity of Chalcone 21-21
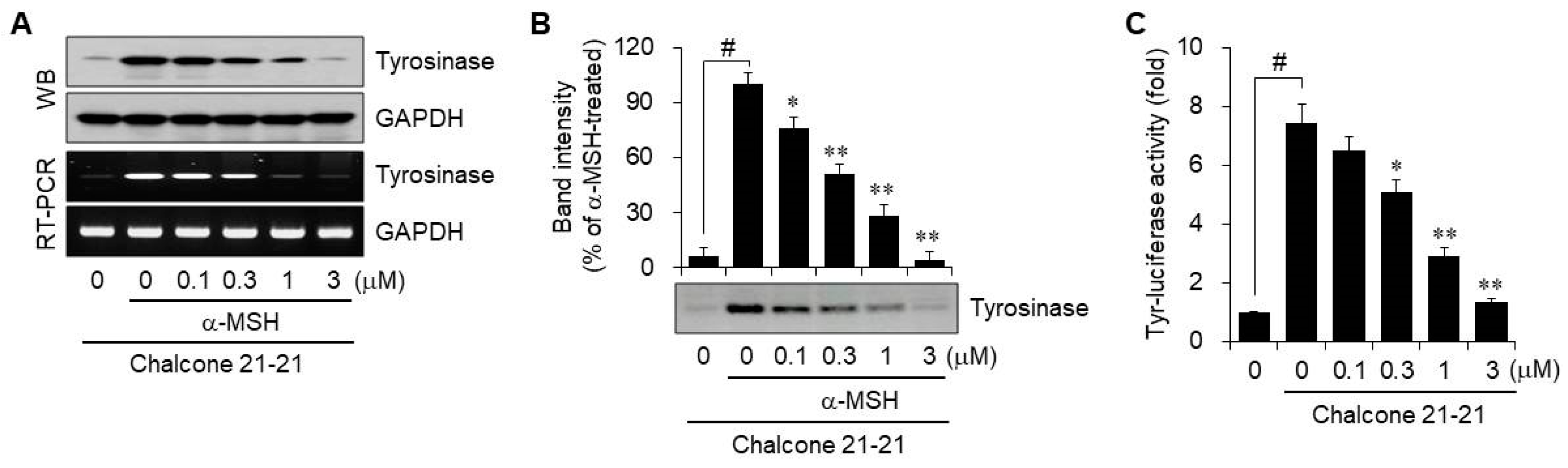
2.2.3. Inhibition of Tyrosinase Activity
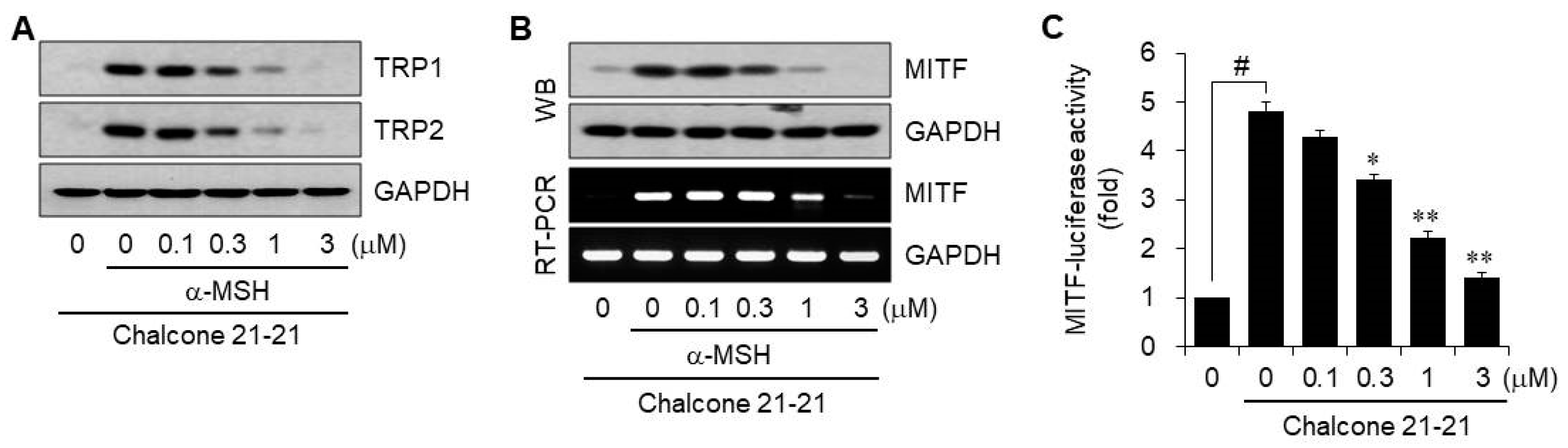
2.2.4. Inhibition of Melanogenesis-Related Proteins
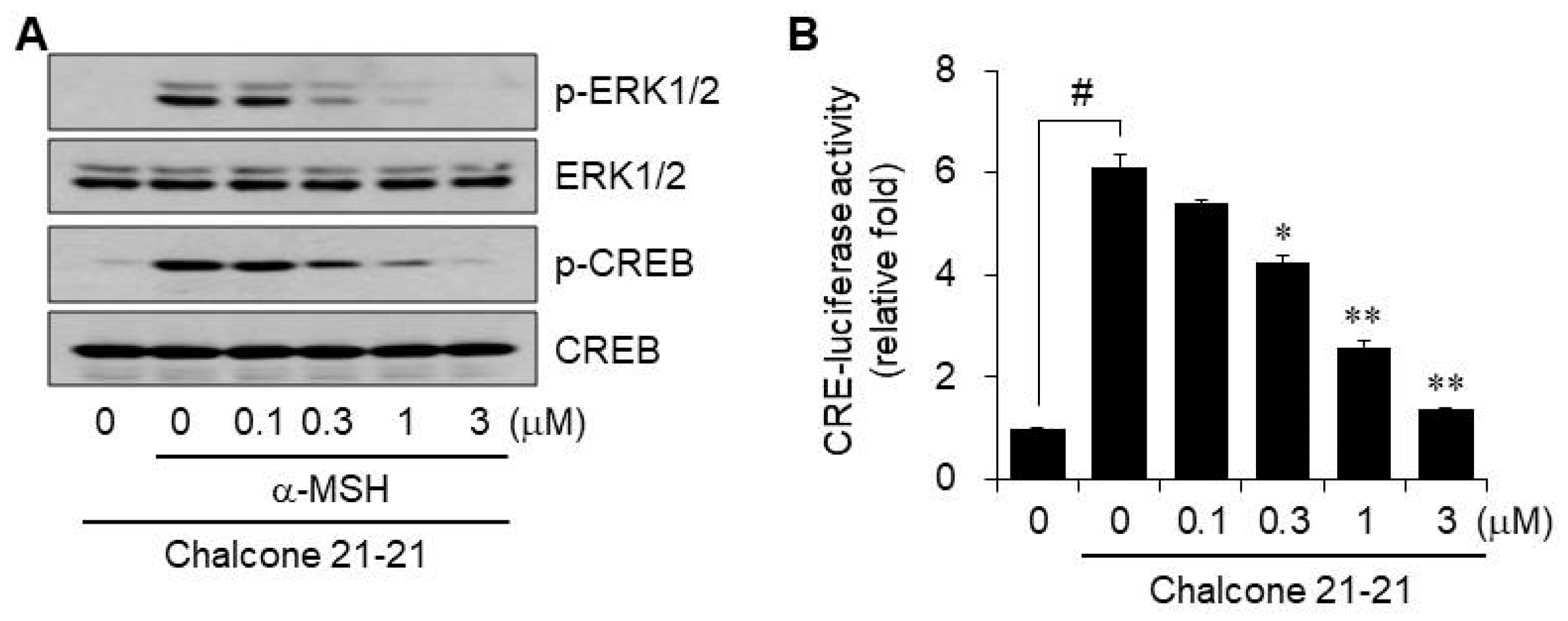
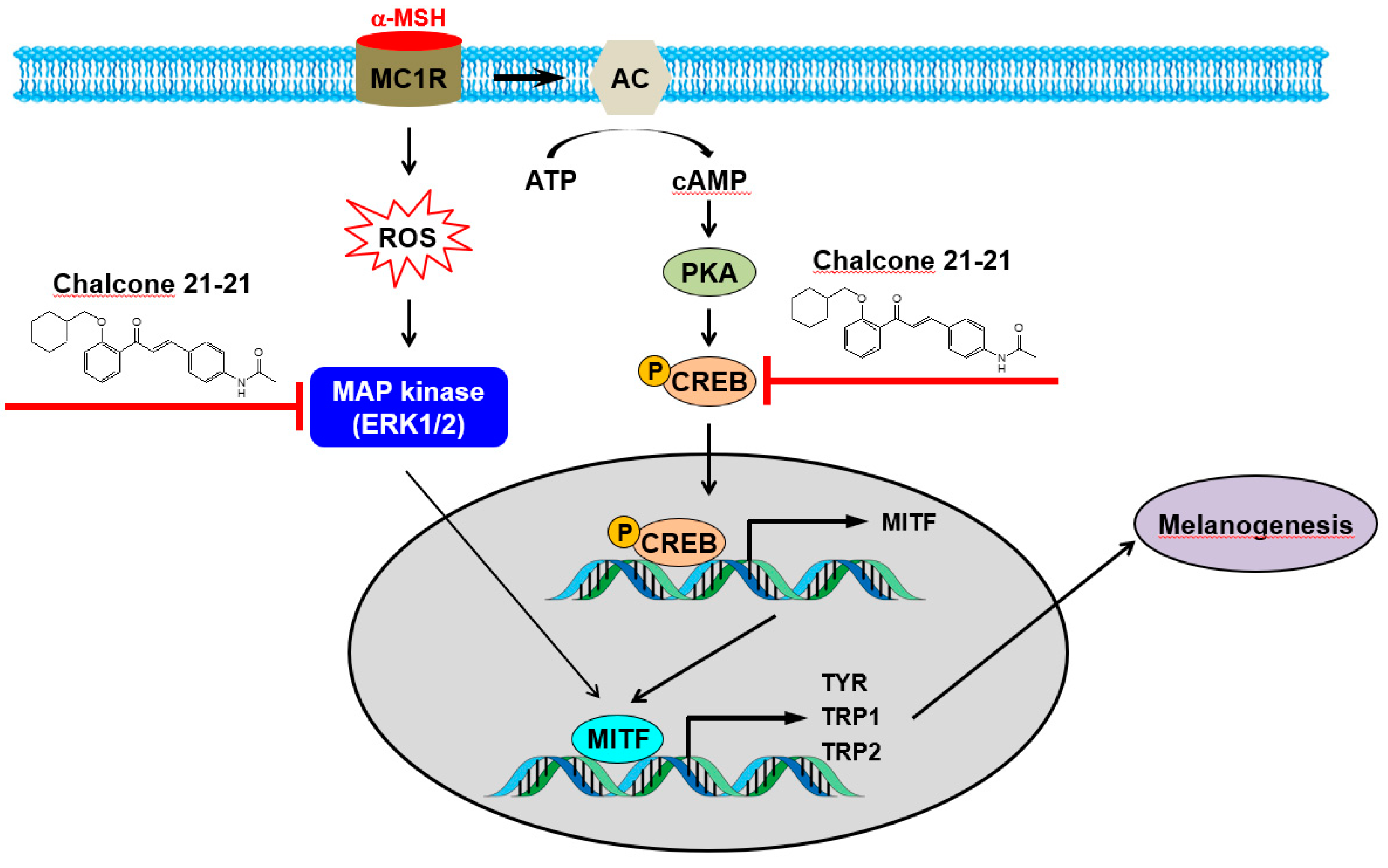
2.2.5. Inhibition of ERK and CREB Signaling
3. Materials and Methods
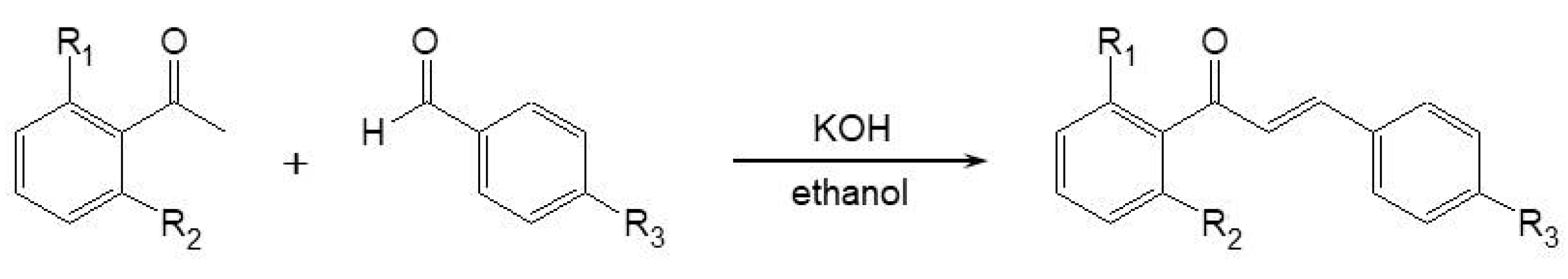
3.1. Chemical Synthesis
3.1.1. (E)-3-(4-Ethylphenyl)-1-(2-hydroxy-6-((4-(trifluoromethoxy)benzyl)oxy)phenyl)prop-2-en-1-one (21-1)
3.1.2. (E)-3-(4-Chlorophenyl)-1-(2-hydroxy-6-((4-(trifluoromethoxy)benzyl)oxy)phenyl)prop-2-en-1-one (21-2)
3.1.3. (E)-1-(2-Hydroxy-6-((4-(trifluoromethoxy)benzyl)oxy)phenyl)-3-(4-methoxyphenyl)prop-2-en-1-one (21-3)
3.1.4. (E)-4-(3-(2-Hydroxy-6-((4-(trifluoromethoxy)benzyl)oxy)phenyl)-3-oxoprop-1-en-1-yl)benzoic acid (21-4)
3.1.5. (E)-N-(4-(3-(2-Hydroxy-6-((4-(trifluoromethoxy)benzyl)oxy)phenyl)-3-oxoprop-1-en-1-yl)phenyl)acetamide (21-5)
3.1.6. (E)-1-(2-(Benzyloxy)-6-hydroxyphenyl)-3-(4-ethylphenyl)prop-2-en-1-one (21-6)
3.1.7. (E)-1-(2-(Benzyloxy)-6-hydroxyphenyl)-3-(4-chlorophenyl)prop-2-en-1-one (21-7)
3.1.8. (E)-1-(2-(Benzyloxy)-6-hydroxyphenyl)-3-(4-methoxyphenyl)prop-2-en-1-one (21-8)
3.1.9. (E)-4-(3-(2-(Benzyloxy)-6-hydroxyphenyl)-3-oxoprop-1-en-1-yl)benzoic acid (21-9)
3.1.10. (E)-1-(2-(Benzyloxy)phenyl)-3-(4-ethylphenyl)prop-2-en-1-one (21-10)
3.1.11. (E)-1-(2-(Benzyloxy)phenyl)-3-(4-chlorophenyl)prop-2-en-1-one (21-11)
3.1.12. (E)-1-(2-(Benzyloxy)phenyl)-3-(4-methoxyphenyl)prop-2-en-1-one (21-12)
3.1.13. (E)-4-(3-(2-(Benzyloxy)phenyl)-3-oxoprop-1-en-1-yl)benzoic acid (21-13)
3.1.14. (E)-N-(4-(3-(2-(Benzyloxy)phenyl)-3-oxoprop-1-en-1-yl)phenyl)acetamide (21-14)
3.1.15. (E)-3-(4-Ethylphenyl)-1-(2-hydroxyphenyl)prop-2-en-1-one (21-15)
3.1.16. (E)-1-(2-Hydroxyphenyl)-3-(4-methoxyphenyl)prop-2-en-1-one (21-16)
3.1.17. (E)-N-(4-(3-(2-Hydroxyphenyl)-3-oxoprop-1-en-1-yl)phenyl)acetamide (21-17)
3.1.18. (E)-1-(2-(Cyclohexylmethoxy)phenyl)-3-(4-ethylphenyl)prop-2-en-1-one (21-18)
3.1.19. (E)-3-(4-Chlorophenyl)-1-(2-(cyclohexylmethoxy)phenyl)prop-2-en-1-one (21-19)
3.1.20. (E)-4-(3-(2-(Cyclohexylmethoxy)phenyl)-3-oxoprop-1-en-1-yl)benzoic acid (21-20)
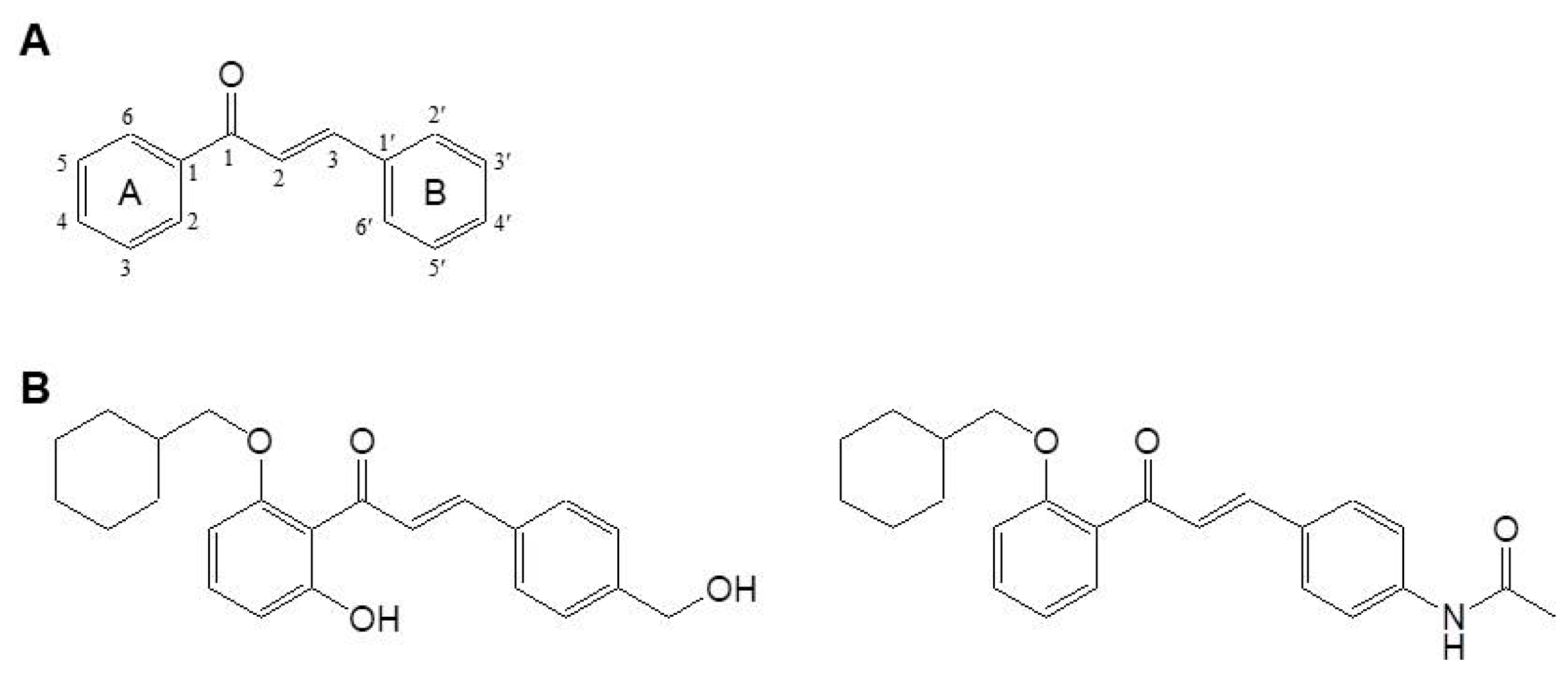
3.1.21. (E)-N-(4-(3-(2-(Cyclohexylmethoxy)phenyl)-3-oxoprop-1-en-1-yl)phenyl)acetamide (21-21)
3.2. Biological Assays
3.2.1. Reagents
3.2.2. Cell Line and Culture Conditions
3.2.3. Cellular Melanin Contents
3.2.4. Cell Viability Assay
3.2.5. Zymography
3.2.6. Western Blot Analysis
3.2.7. Semi-Quantitative RT-PCR
3.2.8. Transfection and Promoter Assay
3.2.9. Statistics
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Costin, G.E.; Hearing, V.J. Human skin pigmentation: melanocytes modulate skin color in response to stress. FASEB J. 2007, 21, 976–994. [Google Scholar] [CrossRef]
- ElObeid, A.S.; Kamal-Eldin, A.; Abdelhalim, M.A.K.; Haseeb, A.M. Pharmacological Properties of Melanin and its Function in Health. Basic Clin. Pharmacol. Toxicol. 2017, 120, 515–522. [Google Scholar] [CrossRef]
- Davis, E.C.; Callender, V.D. Postinflammatory hyperpigmentation: a review of the epidemiology, clinical features, and treatment options in skin of color. J. Clin. Aesthet. Dermatol. 2010, 3, 20–31. [Google Scholar]
- Niu, C.; Aisa, H.A. Upregulation of Melanogenesis and Tyrosinase Activity: Potential Agents for Vitiligo. Molecules 2017, 22, 1303. [Google Scholar] [CrossRef] [PubMed]
- Pillaiyar, T.; Namasivayam, V.; Manickam, M.; Jung, S.H. Inhibitors of Melanogenesis: An Updated Review. J. Med. Chem. 2018, 61, 7395–7418. [Google Scholar] [CrossRef] [PubMed]
- D’Mello, S.A.; Finlay, G.J.; Baguley, B.C.; Askarian-Amiri, M.E. Signaling Pathways in Melanogenesis. Int. J. Mol. Sci. 2016, 17, 1144. [Google Scholar] [CrossRef] [PubMed]
- Pillaiyar, T.; Manickam, M.; Jung, S.H. Recent development of signaling pathways inhibitors of melanogenesis. Cell Signal. 2017, 40, 99–115. [Google Scholar] [CrossRef] [PubMed]
- Bukhari, S.N.; Jasamai, M.; Jantan, I. Synthesis and biological evaluation of chalcone derivatives (mini review). Mini Rev. Med. Chem. 2012, 12, 1394–1403. [Google Scholar] [PubMed]
- Matos, M.J.; Vazquez-Rodriguez, S.; Uriarte, E.; Santana, L. Potential pharmacological uses of chalcones: A patent review (from June 2011–2014). Expert Opin. Ther. Pat. 2015, 25, 351–366. [Google Scholar] [CrossRef] [PubMed]
- Karthikeyan, C.; Moorthy, N.S.; Ramasamy, S.; Vanam, U.; Manivannan, E.; Karunagaran, D.; Trivedi, P. Advances in chalcones with anticancer activities. Recent Pat Anticancer Drug Discov. 2015, 10, 97–115. [Google Scholar] [PubMed]
- Rozmer, Z.; Perjési, P. Naturally occurring chalcones and their biological activities. Phytochem. Rev. 2016, 15, 87–120. [Google Scholar] [CrossRef]
- Zhuang, C.; Zhang, W.; Sheng, C.; Zhang, W.; Xing, C.; Miao, Z. Chalcone: A Privileged Structure in Medicinal Chemistry. Chem. Rev. 2017, 117, 7762–7810. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.H.; Park, K.C.; Park, J.H.; Lee, C.G.; Ye, S.K.; Park, J.Y. Inhibition of tyrosinase activity and melanin production by the chalcone derivative 1-(2-cyclohexylmethoxy-6-hydroxy-phenyl)-3-(4-hydroxymethyl-phenyl)-propenone. Biochem. Biophys. Res. Commun. 2016, 480, 648–654. [Google Scholar] [CrossRef] [PubMed]
- Nakabo, D.; Okano, Y.; Kandori, N.; Satahira, T.; Kataoka, N.; Akamatsu, J.; Okada, Y. Convenient Synthesis and Physiological Activities of Flavonoids in Coreopsis lanceolata L. Petals and Their Related Compounds. Molecules. 2018, 23, 1671. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.Y.; Fisher, D.E. Melanocyte biology and skin pigmentation. Nature 2007, 445, 843–850. [Google Scholar] [CrossRef] [PubMed]
- Wolf Horrell, E.M.; Boulanger, M.C.; D’Orazio, J.A. Melanocortin 1 Receptor: Structure, Function, and Regulation. Front. Genet. 2016, 7, 95. [Google Scholar] [CrossRef]
- Solano, F.; Briganti, S.; Picardo, M.; Ghanem, G. Hypopigmenting agents: an updated review on biological, chemical and clinical aspects. Pigment Cell Res. 2006, 19, 550–571. [Google Scholar] [CrossRef]
- Gunia-Krzyżak, A.; Popiol, J.; Marona, H. Melanogenesis Inhibitors: Strategies for Searching for and Evaluation of Active Compounds. Curr. Med. Chem. 2016, 23, 3548–3574. [Google Scholar] [CrossRef]
- Park, H.Y.; Kosmadaki, M.; Yaar, M.; Gilchrest, B.A. Cellular mechanisms regulating human melanogenesis. Cell. Mol. Life Sci. 2009, 66, 1493–1506. [Google Scholar] [CrossRef]
- Videira, I.F.; Moura, D.F.; Magina, S. Mechanisms regulating melanogenesis. An. Bras. Dermatol. 2013, 88, 76–83. [Google Scholar] [CrossRef]
- Ito, S.; Wakamatsu, K. Chemistry of mixed melanogenesis—Pivotal roles of dopaquinone. Photochem. Photobiol. 2008, 84, 582–592. [Google Scholar] [CrossRef]
- Hearing, V.J. Determination of melanin synthetic pathways. J. Invest Dermatol. 2011, 131, E8–E11. [Google Scholar] [CrossRef]
- Schiaffino, M.V. Signaling pathways in melanosome biogenesis and pathology. Int. J. Biochem. Cell. Biol. 2010, 42, 1094–1104. [Google Scholar] [CrossRef]
- Bonaventure, J.; Domingues, M.J.; Larue, L. Cellular and molecular mechanisms controlling the migration of melanocytes and melanoma cells. Pigment Cell Melanoma Res. 2013, 26, 316–325. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds chalcone 21 and its 21 derivatives are available from the authors. |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Substitutions | Inhibition (%) at 3 μM | Viability (%) at 3 μM | IC50 (μM) | ||
|---|---|---|---|---|---|---|
| R1 | R2 | R3 | ||||
| Chalcone 21 | cyclohexylmethoxy | OH | CH2OH | 41.7 ± 3.3 | 95.1 ± 3.9 | >3.0 |
| 21-1 | 4-trifluoromethoxybenzyloxy | OH | CH2CH3 | 26.3 ± 6.5 | 79.7 ± 3.9 | >3.0 |
| 21-2 | 4-trifluoromethoxybenzyloxy | OH | Cl | 73.9 ± 6.8 | 60.6 ± 3.2 | 1.2 |
| 21-3 | 4-trifluoromethoxybenzyloxy | OH | OCH3 | 47.2 ± 7.8 | 78.6 ± 2.7 | >3.0 |
| 21-4 | 4-trifluoromethoxybenzyloxy | OH | COOH | 38.8 ± 6.2 | 78.9 ± 2.1 | >3.0 |
| 21-5 | 4-trifluoromethoxybenzyloxy | OH | NHCOCH3 | 89.4 ± 8.4 | 76.5 ± 3.1 | 0.88 |
| 21-6 | benzyloxy | OH | CH2CH3 | 42.8 ± 7.1 | 85.0 ± 4.6 | >3.0 |
| 21-7 | benzyloxy | OH | Cl | 65.0 ± 4.9 | 59.8 ± 3.9 | 2.3 |
| 21-8 | benzyloxy | OH | OCH3 | 35.7 ± 4.6 | 74.4 ± 3.1 | >3.0 |
| 21-9 | benzyloxy | OH | COOH | 55.8 ± 6.2 | 72.9 ± 2.2 | 2.6 |
| 21-10 | benzyloxy | H | CH2CH3 | 36.6 ± 5.6 | 77.0 ± 2.6 | >3.0 |
| 21-11 | benzyloxy | H | Cl | 72.0 ± 7.0 | 67.8 ± 3.5 | 2.0 |
| 21-12 | benzyloxy | H | OCH3 | 35.7 ± 5.8 | 81.6 ± 3.5 | >3.0 |
| 21-13 | benzyloxy | H | COOH | 91.8 ± 6.4 | 71.2 ± 2.8 | 1.6 |
| 21-14 | benzyloxy | H | NHCOCH3 | 47.1 ± 6.4 | 74.5 ± 3.8 | >3.0 |
| 21-15 | OH | H | CH2CH3 | 23.8 ± 4.9 | 78.3 ± 2.9 | >3.0 |
| 21-16 | OH | H | OCH3 | 21.6 ± 6.1 | 80.4 ± 3.0 | >3.0 |
| 21-17 | OH | H | NHCOCH3 | 9.4 ± 5.4 | 81.8 ± 4.8 | >3.0 |
| 21-18 | cyclohexylmethoxy | H | CH2CH3 | 29.1 ± 4.9 | 87.6 ± 4.2 | >3.0 |
| 21-19 | cyclohexylmethoxy | H | Cl | 49.7 ± 7.7 | 76.4 ± 3.6 | >3.0 |
| 21-20 | cyclohexylmethoxy | H | COOH | 19.8 ± 8.5 | 90.2 ± 3.2 | >3.0 |
| 21-21 | cyclohexylmethoxy | H | NHCOCH3 | 92.0 ± 5.5 | 86.3 ± 2.5 | 0.54 |
| Gene | Primer | Sequence |
|---|---|---|
| Tyrosinase | Forward | 5′-GAC GGT CAC TGC AGA CTT TG-3′ |
| Reverse | 5′-GCC ATG ACC AGG ATG AC-3′ | |
| MITF | Forward | 5′-GAC GGT CAC TGC AGA CTT TG-3′ |
| Reverse | 5′-GCC ATG ACC AGG ATG AC-3′ | |
| GAPDH | Forward | 5′-TGA AGG TCG GTG TGA ACG GAT TTC GC-3′ |
| Reverse | 5′-CAT GTA GGC CAT GAG GTC CAC CAC-3′ |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, B.-H.; Hong, S.-N.; Ye, S.-K.; Park, J.-Y. Evaluation and Optimization of the Anti-Melanogenic Activity of 1-(2-Cyclohexylmethoxy-6-hydroxy-phenyl)-3-(4-hydroxymethyl-phenyl)-propenone Derivatives. Molecules 2019, 24, 1372. https://doi.org/10.3390/molecules24071372
Kim B-H, Hong S-N, Ye S-K, Park J-Y. Evaluation and Optimization of the Anti-Melanogenic Activity of 1-(2-Cyclohexylmethoxy-6-hydroxy-phenyl)-3-(4-hydroxymethyl-phenyl)-propenone Derivatives. Molecules. 2019; 24(7):1372. https://doi.org/10.3390/molecules24071372
Chicago/Turabian StyleKim, Byung-Hak, Soo-Nam Hong, Sang-Kyu Ye, and Jung-Youl Park. 2019. "Evaluation and Optimization of the Anti-Melanogenic Activity of 1-(2-Cyclohexylmethoxy-6-hydroxy-phenyl)-3-(4-hydroxymethyl-phenyl)-propenone Derivatives" Molecules 24, no. 7: 1372. https://doi.org/10.3390/molecules24071372
APA StyleKim, B. -H., Hong, S. -N., Ye, S. -K., & Park, J. -Y. (2019). Evaluation and Optimization of the Anti-Melanogenic Activity of 1-(2-Cyclohexylmethoxy-6-hydroxy-phenyl)-3-(4-hydroxymethyl-phenyl)-propenone Derivatives. Molecules, 24(7), 1372. https://doi.org/10.3390/molecules24071372