1. Introduction
Langmuir monolayers at the air-water interface, demonstrate several 2-D phases and phase transitions, ranging from gas-like phase to more condensed phases, when compressed laterally [
1]. However, if the monolayer film is compressed beyond its stability limit, a transition from 2-D to 3-D structure occurs. This transition from 2-D to 3-D, referred to as monolayer collapse, occurs at a constant surface pressure. Monolayer collapse has been a subject of interest, especially for biological lung surfactant mixtures that are present at the air-water interface in alveoli of lungs and helps reduce the work of breathing by maintaining near zero surface tension during compression, while maintaining a stable film during monolayer collapse. There are several different mechanisms of collapse, including buckling, budding and vesiculation, as reviewed in detail by Lee [
2]. Further, during cyclic compression and expansion, monolayers can collapse either reversibly or irreversibly. Of particular interest, in lung surfactant films, is the buckling mechanism of collapse. Buckling of the film is characterized by formation of a 3-D structure called folds that forms invaginations into the subphase, retaining the molecules close to the surface. Folds “unfold” upon expansion, allowing the collapsed material to be re-incorporated into the monolayer, thus making the collapse reversible. These folds are formed perpendicular to the axis of compression (or parallel to the barriers used for compression), and are often formed across the entire width of the Langmuir trough. On the other hand, complex amphiphilic mixtures, when compressed, can also be prone to a rejection of molecules from the surface before reaching the limiting area of compression. Collapse following this path is deemed irreversible. In this case, a loss in the material at the interface is inevitable. Therefore, it is believed that proper lung surfactant function requires formation of folds that will allow reversible collapse.
Of particular interest to public health and safety is how inhaled particles, such as nanoparticles and pollutants in the air, effect lung surfactant function. The lung surfactants act as barrier against particles that are small enough to deposit inside the alveoli. Several factors including particle shape, density, the size of the inhaled material, as well as the health status and mode of breathing, play key roles in the fate of such particles inside the lungs [
3]. The size determines the physical mechanism by which the particles deposit in the lungs. Particles that are larger than 5 microns usually deposit through inertial impaction or gravitational sedimentation (1–8 microns), whereas, particles smaller than 500 nm deposit in the lungs through Brownian diffusion [
4]. Thus, depending on the size, the particles can end up either in the upper respiratory tract or even reach the alveoli. Larger particles, usually within 10 to 20 microns, mostly deposit in the upper respiratory tract, whereas, particles as big as 5 microns may deposit in the alveoli [
5]. In case of evaluating the toxicity of nanomaterials, the smaller range is of particular interest as the nanoparticles fall within this limit.
With the advancement in nanotechnology, engineered nanoparticles have been gaining significant grounds in different areas including biomedicine [
6,
7,
8,
9]. The small size of the particles as well as relative ease in surface-tunability make these nanoparticles suitable vehicles for targeted drug delivery amongst other biomedical applications [
10,
11]. Therefore, it is highly likely for nanoparticles to either intentionally or unintentionally enter human bodies, and it is necessary to evaluate the compatibility and the toxicity of the nanoparticles when interacting with various physiological components. In this regard, the respiratory tract is one common route for the entry of the nanoparticles. The small size allows the particles to reach the depths of the alveoli [
12]. Upon entering the alveoli, nanoparticles then interact with LS monolayer. Research has shown the impact of size, hydrophobicity, and concentration of different nanoparticles on the functioning of surfactants [
13,
14,
15,
16,
17,
18,
19,
20]. Many of these studies monitor changes in the surface pressure vs. area isotherms due to incorporation of nanoparticles, thus focusing on the thermodynamic aspects of the changes induced by nanoparticles on model membranes. More recently, some of these studies have also coupled studies focused on changes in the surface pressure with studies monitoring nanoparticle induced changes in the morphology of these films. Such studies have presented new information about the impact of nanoparticles on model lipid monolayers [
13,
14,
15,
16,
21]. For example, Tatur et. al., showed that even though the isotherms of different model surfactants are not affected by their interaction with hydrophobic gold nanoparticles, the morphology at the air-water interface is predominantly altered in the case of the particle exposure [
21]. Thus, in addition to the surface pressure-area isotherms, the surface morphology should be carefully studied while assessing the behavior of the nanoparticles. Similarly, previous work from our lab has shown that the actual composition of the lipid mixture used can also lead to differences in interactions with nanoparticles [
22]. However, the impact of nanoparticles on collapse mechanism in phospholipid monolayers is currently not well understood and has not been studied in detail. Understanding how nanoparticles impact the mechanisms of collapse in lung surfactant monolayers is an important issue that should be addressed and is the main focus of this paper. We are particularly interested in studying the impact of carbon-based nanoparticles, specifically engineered carbon nanodiamonds (ECNs), on the mechanisms of monolayer collapse in phospholipid monolayers.
Studies have shown that the chemistry of the surfactant mixtures (phospholipid combinations and presence of LS proteins or their synthetic analogs) enable fold formation during compression and reincorporation of material during expansion cycles [
23,
24,
25]. On the other hand, our previous work, focused on ECN induced changes in lipid domain packing in the LE-LC regions has shown that differences in lipid-ECN interactions are modulated by other lipid headgroup charge and tail saturation [
22]. Specifically, we observed that at lower surface pressures, the anionic ECNs behave as line active species when interacting with zwitterionic phospholipids. But, in the presence of anionic phospholipids, electrostatic repulsion plays a greater role. We hypothesize that ECN induced changes in mechanisms of monolayer collapse should also depend on the composition of the lipid mixtures used. Therefore, in this work, we focus on ECN induced changes in the reversible and irreversible collapse of model lipid membranes using four different lipid mixtures that reflect the lipid headgroups and tail saturations commonly seen in lung surfactants.
While native surfactants are made up of 90% by weight lipids and 10% by weight proteins, in this work, we only focus on the interactions between the phospholipids and the ECN, in the absence of proteins [
26,
27]. Zwitterionic, disaturated dipalmitoylphosphatidylcholine (DPPC) is the most abundant phospholipid present in biological lung surfactants and is often used either as the major component or on its own to evaluate the efficacy of novel synthetic lung surfactants [
28]. Therefore, in this study DPPC is used as the major lipid component (making up 70% by weight of the lipid mixture). Additionally, unsaturated phosphatidylcholine as well as negatively charged, saturated and unsaturated phosphatidylglycerol are found in native LS mixtures, and are often used in synthetic LS mixtures. Therefore, in this study lipid mixtures containing saturated and unsaturated phosphatidylglycerol were also used at 30 wt.% to study the interaction of ECNs with LS mixtures having a net negative charge reflecting the overall charge of LS monolayers. Lipid mixtures containing DPPC with 30 wt.% unsaturated phosphatidylcholine as well as saturated phosphatidyl-etholamine were used as zwitterionic lipid mixtures. Analysis of cyclic compression expansion surface-pressure isotherms, combined with fluorescence images of the surface of the monolayer during monolayer collapse are together used to arrive at conclusions regarding the impact of ECNs on the mechanisms of monolayer collapse, and the role of lipid headgroup charge and tail saturation on ECN-induced changes in monolayer collapse. Our results together also present ECN-induced changes in the ability of different LS model mixtures to reincorporate material during multiple compression-expansion cycles.
3. Discussion
In this section we discuss the major implications of our results presented above. In the present work, the impact of nanoparticle-phospholipid interactions on multiple compression/expansion cycles were studied, along with direct visualization of the phospholipid monolayer morphology at monolayer collapse, to understand the impact of nanoparticles on different mechanisms of monolayer collapse. Further, we also explored how variations in monolayer composition demonstrate differences in the response to nanoparticle interactions. Multiple compression/expansion cycles are relevant for lung surfactants because a loss in the material is encountered after each cycle of breathing. A greater loss in the material with the addition of nanomaterial suggests a detrimental impact of the particles on the proper functioning of the surfactants. Kodama et al., used multiple compression/expansion cycles to discuss how particle size, ranging from 20 nm to 1.0 μm, affect the phase behavior of surfactant monolayers [
30]. While several previous studies have focused on the effect of nanomaterial on the surface activity and surface morphology of native as well as model lung surfactant monolayers [
13,
16,
31], to the best of our knowledge, the impact of nanoparticles on collapse mechanisms in lipid systems containing anionic, zwitterionic or mixed anionic/zwitterionic lipids is currently unknown.
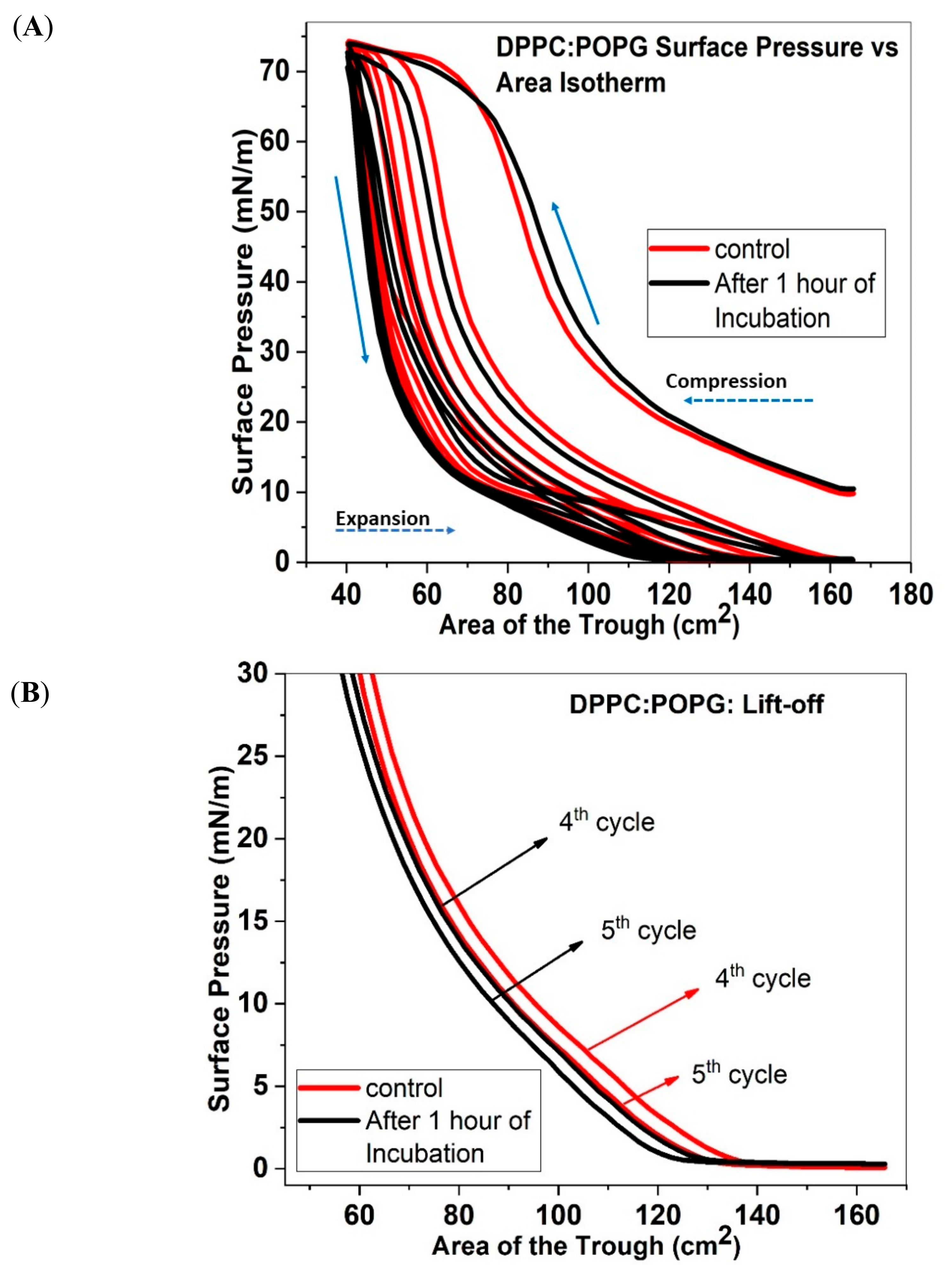
Therefore, in this work, we use multiple compression/expansion cycles to understand how the same nanoparticle influences the mechanisms of collapse in four different lipid systems that demonstrate different collapse mechanisms due to differences in headgroup charge and tail saturations. Surface pressure vs. area isotherms were used to calculate the total area under the curve as well as a shift in the lift-off area in compression isotherms. An increase in the total area under the curve indicates an increase in material retention at the air-water interface, while a shift in the lift-off area to higher areas suggest more material at the interface. Additionally, fluorescence images provide direct visual evidence of the mechanisms of collapse (formation of collapse cracks vs. vesicles).
Overall, our data suggest that the impact of ECN on the reversibility of phospholipid monolayers is dependent on both the charge of the monolayer as well as the lipid tail saturation. Fluorescence imaging provides clear evidence that addition of small amounts of these negatively charged nanoparticles did not change the actual mechanism of collapse for all the different systems studied. However, direct visual imaging fails to reveal the subtle changes observed when ECN interacts with phospholipid mixtures. These ECN induced subtle changes in monolayer collapse could only be captured by analyzing multiple cycles of compressions and expansions.
3.1. Membrane Packing Influences ECN’s Ability to Modulate Monolayer Collapse in Mixed Lipid Systems
We start by analyzing our results for the DDPC:POPG and DPPC:DPPG mixtures. Both of these mixtures are often used as model LS mixtures and are therefore biologically relevant. It is important to note that while both lipid systems have an overall negative charge due to the PG headgroup, the difference lies in the tail saturation. DPPC:DPPG lipid mixture is more packed than DPPC:POPG, due to the unsaturation in the POPG tails. A comparison of our fluorescence images for these two systems with and without the nanoparticles show that both these mixtures undergo reversible monolayer collapse. However, an analysis of the multiple compression/expansion studies show that when nanoparticles are added to lipid mixtures, an increase in material retention is measured between the compression/expansion cycles containing saturated lipids only. On the other hand, for the mixtures containing unsaturated POPG lipids, a loss in material retention is measured. Further, for the saturated DPPC/DPPG lipid mixture, we find that the trough area where “lift off” occurs is shifted to higher areas, while the “lift off” area is shifted to lower areas for the mixtures containing unsaturated POPG. Similarly, our compressibility data shows that the trough area where monolayer collapse occurs is also shifted to higher area for lipid films containing only saturated lipids, while films containing a mixture of saturated and unsaturated lipids have to be compressed further before they reach collapse, when ECN is added to them suggesting there is less reincorporation of material than the control system. These results together suggest that saturated lipids better enable incorporation of the nanoparticles into the monolayers, and thus have a positive impact on the overall performance. On the other hand, the presence of unsaturated POPG lipids seem to cause a detrimental effect on the monolayers’ ability to collapse reversibly and retain material at the interface. Based on this comparison we conclude that interactions of the nanoparticles with unsaturated lipid mixtures cause increased material loss during monolayer collapse.
This conclusion is further validated by our results for the DPPC:POPC system. Addition of ECN to this monolayer system shows the highest loss of material among all the systems studied here. One possible explanation for this behavior is the possible changes to the line energy of the lipid monolayer induced by the nanoparticles due to their interactions with the lipid headgroups. We have previously shown that in case of lipid monolayers containing POPG, the positively charged nanoparticles avoid the negatively charged POPG lipids that are present in the more fluid liquid-expanded (LE) region and cause a lowering of the line tension. Similarly, for the DPPC:POPC system, we have previously shown that ECNs induce a lowering of the line tension between domain boundaries in this system, as evidenced by a transition in the domain morphology from the signature kidney-bean domains seen in DPPC systems, to domains with arms. On the other hand, in case of the saturated DPPC:DPPG lipid system, addition of ECN caused an increase in the line energy of the system by partitioning into the more fluid LE phase. These results together suggest that ECN-induced lowering of the line tension of monolayers containing mismatched lipid tails adversely impact their ability to collapse reversibly.
3.2. Lipid Headgroup Charge Influences the Ability of ECN to Alter Monolayer Collapse
To explore if ECN induced changes in monolayer collapse depends on the presence of anionic lipid headgroups, we compare our results for lipids with zwitterionic vs. anionic lipids, both for saturated and unsaturated lipid systems.
A comparison between the DPPC:POPG and DPPC:POPC systems show that addition of ECN induces a higher loss in material between multiple cycles for the zwitterionic DPPC:POPC system, when compared with DPPC:POPG monolayers containing a net negative charge. Since both of these systems have the same combination of tail saturation, we can attribute the difference in ECN’s influence on monolayer collapse to a difference in the lipid headgroup.
The negative charge on the ECN causes it to avoid the PG headgroup and therefore interact less in the monolayer. This lack of interaction may cause less loss of material between multiple compression/expansion cycles. A comparison of DPPC:DPPG and DPPC:DPPE monolayers show that the increase in material retention induced by ECNs is also more pronounced in the lipid monolayer containing anionic headgroup (note that there is no mismatch in the lipid tails in these two systems). Together, these results suggest that negatively charged nanoparticles produce a stronger change in monolayer collapse for lipid monolayers containing anionic lipids.
3.3. Fluorescence Imaging of the Monolayer Shows that the Mechanism of Monolayer Collapse is not Altered by ECN
Gopal et al. have shown that DPPC:POPG monolayers, laterally compressed under conditions, which are similar to our study, collapses via reversible folding mechanism [
32]. These reversible folds appear as bright streaks perpendicular to the direction of compression, and range from 100 μm to about 1 mm in length. These reversible structures unfold when the monolayer is expanded, and material reincorporates into the monolayer without notably altering the morphology. Our study shows the occurrence of similar collapse features in the case of DPPC:POPG, DPPC:DPPG and DPPC:DPPE. With the addition of ECN, bright streaks were again seen spanning across the fluorescence micrograph, suggesting that the monolayer retains this collapse feature in all three lipid systems. To prove that this behavior can be reproduced regardless of the mechanism of collapse, we imaged DPPC:POPC monolayers in the absence and presence of ECNs. DPPC:POPC monolayers collapses with the formation of vesicles, which appears as bright specks on the monolayer. These bright specks are thought to be globular vesicles that usually detach from the monolayer [
32]. Large vesicles usually end up detaching from the monolayer making the collapse irreversible. In the case of DPPC:POPC control, the formation of the vesicles suggests irreversibility of the monolayer which was also confirmed by the loss of area using compression/expansion isotherms. When ECN was added to the DPPC:POPC mixture, more vesicles appeared on the surface at monolayer collapse. This confirms an increase in the loss of material from the surface when they are compressed beyond the collapse pressure. Again, this loss of material confirms what we inferred, based on our analysis of the isotherm data. Therefore, through fluorescence images, we can conclude that ECN doesn’t alter the mechanism of collapse regardless of the phospholipid it encounters. However, thorough analysis of the compression/expansion cycles over many cycles presents subtle differences in ECN–induced changes to monolayer collapse.
4. Materials and Methods
4.1. Materials
The phospholipids, 1,2-dipalmitoyl-sn-glycero-3-phosphocholine (DPPC), 1-palmitoyl-2-oleoyl-sn-glycero-3-phospho-(1’-rac-glycerol) (sodium salt) (POPG), 1-palmitoyl-2-oleoyl-glycero-3-phosphocholine (POPC), 1,2-dipalmitoyl-sn-glycero-3-phospho-(1′-rac-glycerol) (sodium salt) (DPPG), and 1,2-dipalmitoyl-sn-glycero-3-phosphoethanolamine [DPPE] were obtained from Avanti Polar Lipids (Alabaster, AL, USA). The phospholipid mixtures were purchased in chloroform mixtures at concentrations of 5 or 25 mg/mL. The phospholipid dye that was used in our study, Texas red 1,2-dihexadecanoyl-sn-glycero-3-phosphoethanolamine, triethylammonium salt (TXR-DHPE), was obtained from Life Technologies (Invitrogen, Grand Island, NY, USA). The engineered carbon nanodiamonds were procured from Microdiamant (Lengwil, Switzerland). Details about the physical properties of these ECNs have been published before. Briefly, the size of the ECN is 240 nm in the organic mixture, and the accompanying polydispersity is 0.35. Furthermore, the ECN used in this study is negatively charged with a zeta potential of −28 mV.
The phospholipids as well as the dye were diluted to 1 mg/mL in high-performance liquid chromatography (HPLC)-grade chloroform before using them in our studies. ECN suspensions were also prepared in the chloroform:methanol solutions. Chloroform, methanol, acetone, and isopropanol used in this study for preparing samples and cleaning equipment, were purchased from Thermo Fisher Scientific Inc. (Pittsburgh, PA, USA). The water, used as the cleaning agent and sub-phase, had a resistivity of 18.2 MΩ/cm), which was prepared in a Millipore gradient system (Billerica, MA, USA).
4.2. Methods
4.2.1. Sample Preparation
Table 1 shows the phospholipid-ECN solutions used in our study. Solutions of DPPC:POPG, DPPC:POPC, DPPG mixtures (7:3 by weight) were prepared in chloroform. 1 weight % TXR-DHPE dye (dissolved in 4:1 chloroform:methanol mixture) was added to the lipid samples. Carbon nanodiamonds suspensions were sonicated for 2 h, and immediately afterward, stoichiometric volumes were added to the lipid mixtures for the experiments involved in this study.
4.2.2. Langmuir Studies
Nanodiamonds were mixed with the lipid samples at a concentration of 1 wt.%. While in a previous study varying concentrations were used, in this work we chose to use 1 wt.% based on our previous work [
25]. The ECN were sonicated for 2 h, added to the lipid mixtures and added dropwise on the surface of ultrapure water contained in a Langmuir Ribbon Trough, purchased from Biolin Scientific Inc. (Phoenix, AZ, USA). The trough consists of movable ribbon that can compress/expand such that the molecules on the surface can go through different phases. The multiple compression and expansion cycles serve as a model that mimics the decrease and increase in the alveolar area with exhalation/inhalation. The maximum area of the trough is 166 cm
2, and the minimum area is 46 cm
2, which provided the samples with enough area to reach high surface pressure values upon compression. Furthermore, the ribbon is computer controlled to move at a uniform rate with the help of the software supplied by Biolin Scientific Inc. Material was added to the surface in such a way that the starting surface pressure was around 10 mN/m. Although surface pressure vs. area studies are often started at an initial surface pressure of 0 mN/m, in this work a higher starting surface pressure was used to ensure that collapse pressure was reached for all five compression cycles (see
Figure S5). After spreading the sample solution on the water surface and before starting the compression/expansion cycles, the chloroform was allowed to evaporate for 20 min. This waiting period also provides sufficient time for the monolayer to spread uniformly on the surface. After the 20-min period, the ribbon was moved at the rate of 125 mm/min for the compression/expansion isotherms. However, in the case of fluorescence imaging, a slower rate of 7.0 mm/min was used for clarity.
4.2.3. Fluorescence Imaging
The trough is also coupled with an Eclipse fluorescence microscope (Nikon, Japan) for visualizing the surface morphology of the monolayer. The microscope is equipped with a 40×–long working distance objective lens along with motorized-focusing capabilities that allow us to monitor the surface of the monolayer continuously. A dichroic mirror/barrier filter assembly is used in this setup to direct the excitation light perpendicular to the monolayer, whereupon, the emitted light is filtered and captured by the microscope coupled with a fast CCD camera (Andor Luca, Twin Cities, MN, USA). For our purposes, we recorded images in sequences of five to observe the morphology.
4.3. Theoretical Analysis
4.3.1. Analysis of Material Loss
Area under the Curve and Percentage Recovery
Surface pressure versus area isotherms can provide evidence of interaction between molecules at the interface. With the progression of compression/expansion cycles, the isotherm for most Langmuir monolayers is expected to shift to lower areas, which is indicative of the loss in material once the film is compressed beyond its collapse pressure. Further, it is important to notice that a Langmuir film compressed beyond its collapse pressure shows a difference in its Π-A isotherm between the compression and expansion cycle. This difference in the two Π-A isotherms between the compression and expansion cycle is often referred to as the hysteresis in the surface tension. Almost all lung surfactant mixtures demonstrate this hysteresis [
29]. Therefore, we focused on analyzing the effect of addition of nanoparticles to the hysteresis of the different lipid mixtures. Hysteresis corresponds to the area enclosed within the two curves, which forms an envelope. It can be measured from the P-A isotherms, by calculating the polygon area (integral area under the curve) function in Origin 2017. A lowering of such area shows the extent of material loss, which is one of the primary focuses of our study.
4.3.2. Model Based on Volmer’s Equation of State for the Prediction of the Π-A Isotherm at the Region of 2-Dimensional Coexistence Phase
Equations of state [EOS] at the air-water interface can be used to predict the two-dimensional phase coexistence region of the Π-A isotherms of amphiphilic monolayers. Fainerman and Volhardt have described such EOS for insoluble Langmuir monolayers, which is capable of predicting the Π-A isotherms at the gaseous region as well as the 2D phase transition for single amphiphile [
33]. Another theoretical model published by the same group describes the liquid expanded region of the Π-A isotherms of different amphiphilic molecules [
34]. Recently, Ghazvini et al. used the theoretical model proposed by Feinerman and Volhardt to understand the impact of pH on the packing of phospholipid membranes [
35]. Furthermore, these equations are also capable of predicting the material loss. Kodama used a modified form of this equation to calculate the material loss in a lipid mixture due to exposure to nanoparticles of different sizes. Here, we represent the key equations that have been used in the present work to quantify the material loss at the interface.
Mathematically, Volmer’s equation is expressed as follows (Equation (3)):
where π represents surface pressure of the monolayer, k is the Boltzmann constant, T is the temperature, ω is the average effective molecular area of the insoluble species, ω
o is the molecular area per water molecule, A is the available surface area per insoluble molecule, and π
coh is the cohesion pressure. However, the available surface area, A, requires the knowledge of the number of molecules, n, at the surface. A can be related to the trough area, A
T, as follows:
Using the definition for A from Equation (4), in Equation (3), Kuo et. al. characterized the material loss from the surface [
27] using Equation (4) below:
Here, ω
eff is an effective total molecular area, and is given by Equation (6):
Equation (5) can be directly fitted to the π-A isotherms. Additionally, ωeff helps us identify the extent of material loss.
However, it should be noted that equation 4 works only in the LE region. Since lipid mixtures with saturated lipids only do not often have a large LE phase at room temperature, we used equation 4 to calculate material loss in lipid mixtures containing unsaturated lipids.
5. Conclusions
In conclusion, our results, discussed above, demonstrate that the negatively charged ECN has varying impact on collapse of phospholipid monolayers.
While our fluorescence images after the first compression cycle shows that ECNs do not impact the mechanism of monolayer collapse, a detailed analysis of the surface pressure area (Π-A) isotherms over five consecutive compression/expansion cycles show that lipid mixtures containing ECN show a difference in material loss and material re-incorporation between the different lipid systems. Specifically, we find that when interacting with phospholipid mixtures containing saturated lipids only, ECN causes a net positive effect on monolayer collapse by improving the adsorption of material from the subphase. On the other hand, ECN causes more material loss between cycles for systems containing unsaturated lipids. Further, we find that for both saturated and unsaturated lipid mixtures, the presence of negative charge enables more retention/re-incorporation of material into the monolayer interface, when compared with the neutral lipids. Finally, ECN has the most negative impact on neutral unsaturated DPPC:POPC lipid mixture and an improvement in the compression/expansion isotherms for negative saturated DPPC:DPPG mixture. Since several of these lipid mixtures are used as model lung surfactant mixture, we conclude that it is crucial to study the impact of nanoparticles on multiple compression/expansion cycle to gain knowledge on the toxicity of nanomaterial on lung surfactants. Further, since lung surfactants contain unsaturated anionic lipids, our results suggest that the monolayer collapse mechanism of lung surfactants may not be impacted significantly due to the presence of nanoparticles. However, it is important to note here that one must be careful when drawing conclusions from model systems such as used here, since we have shown that the composition of the lipid mixtures play an important role when interacting with the nanoparticles. Further, the rate of compression may also play a role. The rate of compression used in these experiments does not reflect the dynamic nature of compression cycles that lung surfactants undergo during breathing and may impact the results. Therefore, the impact of nanoparticles on lipid mixtures during quasi-static vs. dynamic compressions should also be considered. In closing, our results clearly establish that when studying interactions between nanoparticles and model lipid mixtures, differences due to the headgroup charge and tail saturation of the lipid mixtures should also be accounted for, before drawing generalized conclusions about nanotoxicity of a material.















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}