1. Introduction
Actinomycetes representing the genus
Streptomyces are remarkably potent bacterial producers of antibiotics [
1]. A plethora of antimicrobial compounds has been isolated from
Streptomyces so far, including streptomycin, erythromycin, rapamycin, and neomycin [
2]. In this large group of structurally and functionally diverse molecules some substances have been investigated for decades, mostly in the context of biosynthetic origins, strain development, and bioprocess optimization. Many diverse strategies have been developed to improve the production of antibiotics, including the rewiring of regulatory networks through removal of repressor genes, overexpression of positive regulators, ribosome engineering (see the review of Xia et al. [
3] and references therein), introduction of drug resistance-producing mutations [
4], chemical and physical mutagenesis [
5], genome engineering [
6], and the optimization of bioprocess conditions and medium composition [
7]. Several recent studies focused on the production of oxytetracycline, a broad-spectrum antibiotic, by
Streptomyces rimosus [
8,
9,
10,
11]. This species is also known to secrete an antifungal substance rimocidin [
12,
13] and harbors the genetic basis for the desferrioxamine siderophore biosynthesis [
14]. In a recent study, Kuhl et al. [
15] demonstrated that inoculating actinobacteria into the liquid medium containing talc microparticles resulted in morphological and production-related alterations compared to the conventional cultures. Among other findings, it was shown that the microparticle-enhanced cultivation can be successfully used to improve oxytetracycline titers in the cultivation broths of
S. rimosus. In the present work, the co-inoculation of
S. rimosus with the spores of different species was evaluated as an alternative approach of influencing the growth and metabolic capabilities of
S. rimosus. The idea was to comparatively evaluate the morphological and biosynthetic outcomes of
S. rimosus co-cultivation with several different, morphologically diverse filamentous microorganisms. To broaden the perspective on the biosynthetic capabilities of
S. rimosus in co-cultures, the relative amounts of other detected secondary metabolites (the byproducts of oxytetracycline production) were also investigated.
The main goal of the present work was to evaluate the production of oxytetracycline by S. rimosus in submerged co-cultures with other filamentous microorganisms in relation to the observed morphological characteristics.
2. Results
In the present study,
S. rimosus was confronted with morphologically and biochemically diverse filamentous microorganisms in the two-species co-cultures performed under submerged conditions in shake flasks. The group of co-culture partners included three species of filamentous fungi classified as
Ascomycota, namely
Penicillium rubens,
Chaetomium globosum, and
Aspergillus niger, a fungus representing
Zygomycota, namely
Mucor racemosus, and
Streptomyces noursei, a filamentous microorganism belonging to
Actinobacteria. Most of the strains applied here were previously characterized by Kowalska et al. [
16,
17] in terms of their morphological development under submerged conditions. The co-cultures were characterized with respect to antibiotic production (
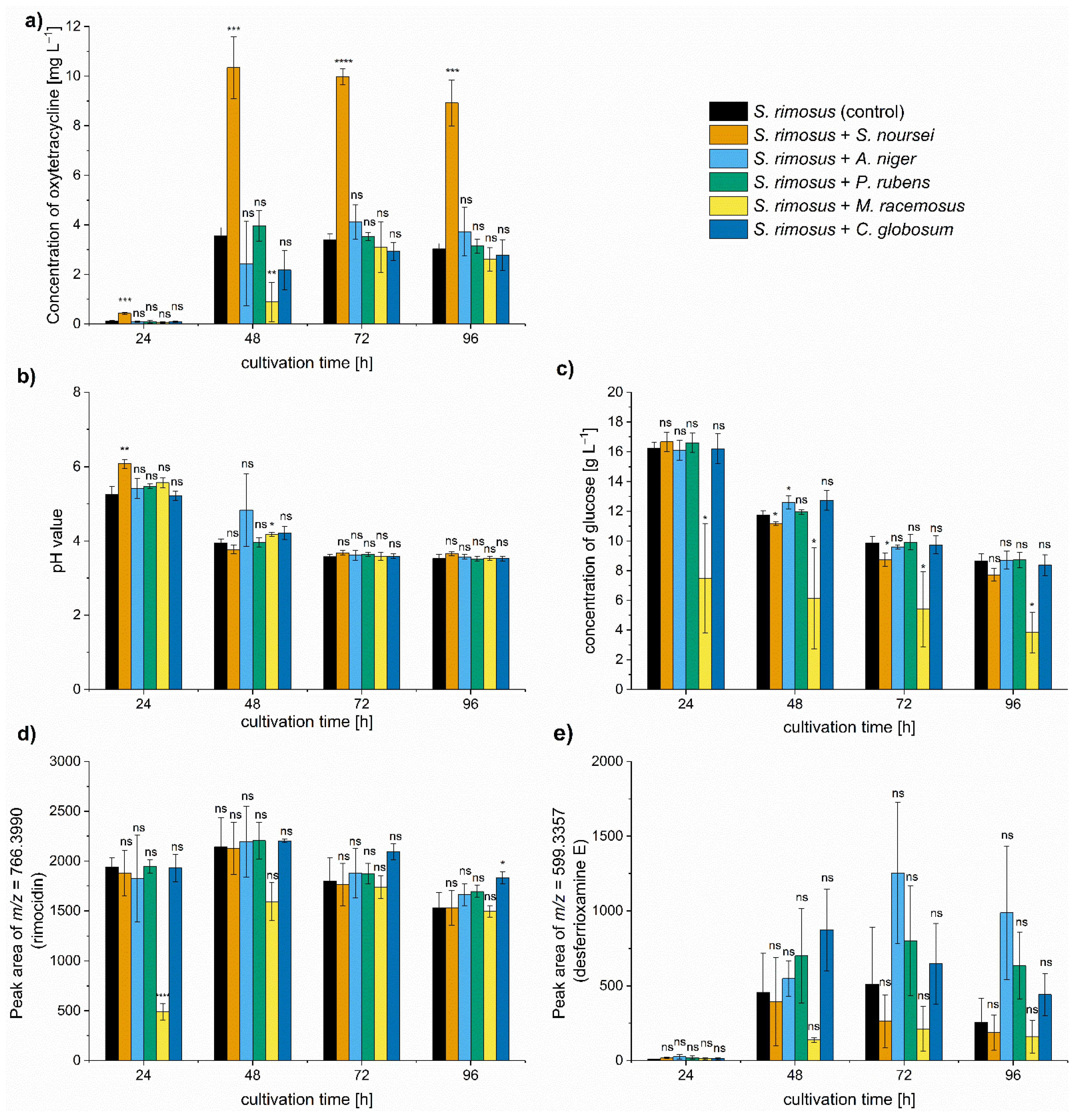
Figure 1a), pH levels (
Figure 1b), and the utilization of carbon source (glucose) (
Figure 1c). Additional experimental steps involved the semiquantitative analysis of the byproducts of oxytetracycline production, namely rimocidin (
Figure 1d) and desferrioxamine E (
Figure 1e), identified by considering their respective
m/
z values in agreement with literature data. The morphological parameters (projected area, elongation, roughness, and morphology number) were determined based on microscopic observations conducted during the cultivation process (
Figure S1).
The most important observation concerned the improvement of oxytetracycline production in the co-cultivation variant with
S. noursei. The mean concentration of this antibiotic in the monoculture control reached 3.5 mg L
−1 in 48 h of the run, whereas in the “
S. rimosus +
S. noursei” co-culture the value of 10.3 mg L
−1 was recorded (
Figure 1a). The remaining co-cultures also exhibited changes with respect to oxytetracycline titers, but the differences relative to the
S. rimosus monoculture were not found to be significant (
Figure 1a). It was also noted that the biosynthesis of oxytetracycline in “
S. rimosus +
M. racemosus” co-culture was visibly delayed compared to other tested variants (
Figure 1a). The outcomes of oxytetracycline production were confronted with the measurements of pH (
Figure 1b) and glucose levels in the broth (
Figure 1c). After 24 h, the pH was below the value of 6 in all tested cultivations except the “
S. rimosus +
S. noursei” co-culture. However, starting from 72 h of the run, the pH ultimately stabilized at the level between 3.5 and 3.7 in all tested cases (
Figure 1b). As far as the concentration of carbon source was concerned, it proceeded without striking differences among the variants, except for the “
S. rimosus +
M. racemosus” co-culture, in which glucose consumption was stimulated (
Figure 1c). The next step was to investigate whether stimulating oxytetracycline biosynthesis by
S. noursei was accompanied by enhancing the production of other secondary metabolites. The peaks at
m/
z = 766.3990 and
m/
z = 599.3357 agreed with the theoretical [M-H]
− ion values of rimocidin (Δ
m/
z = −0.0024) and desferrioxamine E (Δ
m/
z = −0.0048), respectively, and their relative amounts were assessed by determining the respective peak areas. No significant stimulation of rimocidin biosynthesis was revealed in “
S. rimosus +
S. noursei” nor in any other co-culture (
Figure 1d). Its production was delayed in the presence of
M. racemosus, which agreed with the observations made for oxytetracycline (
Figure 1a). Although in the case of desferrioxamine E the variability of peak area values recorded among the independent experiments was rather high, there was no doubt that the oxytetracycline-favoring co-cultivation of
S. rimosus with
S. noursei did not elevate the levels of desferrioxamine E (
Figure 1e).
With regard to the morphological characteristics of
S. rimosus, the qualitative differences among the tested variants were observed in microscopic images (
Figure S1a). The variability of projected area (
Figure S1b), roughness (
Figure S1c), elongation (
Figure S1d), and morphology number (
Figure S1e) values was rather high among the tested variants, as indicated by the standard deviation bars depicted in
Figure S1, but in the digital image analysis of filamentous morphologies this was not a surprising behavior [
16,
17]. In most cases, no significant differences of morphological parameter values were recorded among the variants. Importantly, considering the results of the quantitative morphological analysis, the “
S. rimosus +
S. noursei” co-culture was not found to be exceptional compared to other investigated co-cultures (
Figure S1). The changes of the so-called morphology number, a formula that combines several relevant morphological parameters [
18], did not differ significantly among the tested variants (
Figure S1e).
As an additional element of the study, the total ion chromatograms (TICs) were compared to determine if the co-cultivation led to the awakening of silent biosynthetic gene clusters in the investigated strains. However, no triggering of cryptic production pathways was found. Instead, an observation regarding the dominant role of
S. rimosus was made when the TICs from all cultures were aligned (
Figure S2). The chemical profiles of
S. rimosus-involving co-cultures resembled the one displayed by
S. rimosus itself (
Figure S2a), while being distinct from the monocultures of its microbial partners considered in the study (
Figure S2b). The chemical similarity between the cultivation broths containing
S. rimosus was interpreted as a display of the dominance of this actinobacterium over the accompanying species. This was also confirmed by the microscopic observations of monocultures (
Figure S3). If
S. rimosus was present in the co-culture, it prevented the microbial partner from achieving its usually observed mature (in terms of size and shape) morphological form. There was, however, one exception in this respect, namely the co-cultures with
M. racemosus. In this case, the fully developed morphological forms of
M. racemosus (both the yeast-like and branching cells) were easily noticeable next to
S. rimosus in the cultivation broth (
Figure S1a).
Arguably, the most frequently used approach of initiating the
Streptomyces-involving co-cultures is to inoculate the production medium with the use of precultures (e.g., see the works reviewed in [
19]). In the present study, the spores of participating species were applied instead. As the method of confronting the microorganisms in co-culture is one of the factors determining the productivity of the process, it was questioned whether the stimulatory effect of the “
S. rimosus +
S. noursei” co-culture would still be observed under different inoculation conditions. To address this, the spores of
S. rimosus were used to co-inoculate the medium either with the spores or the 24 h preculture of
S. noursei and the 60 h cultivation process was performed. In parallel, an alternative approach was tested that involved the confrontation of the 24 h preculture of
S. rimosus with the spores or the 24 h preculture of
S. noursei. The types of data gathered for various inoculation scenarios (“spores versus spores”, “spores versus preculture”, or “preculture versus preculture”) were the same as in the multispecies experiment described above and involved the oxytetracycline concentration (
Figure 2a), pH values (
Figure 2b), glucose utilization (
Figure 2c), levels of rimocidin (
Figure 2d), and desferrioxamine E (
Figure 2e), as well as the morphological characterization (
Figure 3).
The use of
S. rimosus preculture for inoculation resulted in relatively low titers compared to the cultivations initiated from spores (
Figure 2a). In both cases, however, the production of oxytetracycline was markedly improved due to the co-inoculation with the spores of
S. noursei (
Figure 2a). The stimulatory effect was visible not only when the spores of
S. noursei were confronted with the spores of
S. rimosus, but also in the “
S. rimosus preculture versus
S. noursei spores” case. Confronting the precultures of both species did not result in a significant production enhancement relative to the control. Finally, when the developed biomass of
S. noursei was confronted with the spores of
S. rimosus, the domination of the former took place and only trace levels of oxytetracycline were detected (
Figure 2a). Hence, it was clear at this point that the co-culture initiated with the use of spores was effective in terms of improving oxytetracycline production. According to the results, however, this behavior was not associated with considerable differences in pH values (
Figure 2b) nor glucose consumption (
Figure 2c). It was also noted that the biosynthesis of rimocidin (
Figure 2d) was less affected by the method of inoculation (preculture or spores) compared to oxytetracycline (
Figure 2a) and desferrioxamine E (
Figure 2e).
The morphological forms developed in the flasks inoculated with the use of
S. rimosus precultures were generally of greater size than their counterparts in the spores-inoculated cultures (
Figure 3a,b). When
S. rimosus was confronted with
S. noursei preculture, the presence of
S. noursei led to a marked decrease in projected area compared to the control. This was not observed when
S. noursei was added to the medium in the form of spores (
Figure 3b). Providing
S. noursei with a 24 h growth advantage over
S. rimosus resulted in a clumped morphology more resembling the
S. noursei monoculture (see
Figure S3a) than the
S. rimosus monoculture (depicted in
Figure 3a). Considering the remaining morphological parameters, namely roughness (
Figure 3c), elongation (
Figure 3d), and morphology number (
Figure 3e), it was noted that the “spores versus spores” variant, effective in the context of oxytetracycline production, did not show the morphologies that would make it stand out from other investigated cultures.
3. Discussion
In the present study,
S. rimosus was co-cultivated with several diverse filamentous microorganisms under submerged conditions. Two of the tested microbial duos stood out among the tested variants, namely “
S. rimosus +
S. noursei” and “
S. rimosus +
M. racemosus”. The former turned out to be an effective combination in terms of oxytetracycline production, with one of the participating strains being able to dominate its partner from the very onset of cultivation. On the other hand, the latter pair of microorganisms could be regarded as an example of two fast-growing species that compete for nutrients and space within a limited liquid volume. This was reflected by the fact that in the early phase of the “
S. rimosus +
M. racemosus” co-culture the utilization of glucose was visibly stimulated compared to other tested variants (
Figure 1c) due to the parallel catabolic activities of both species. Moreover, the production of oxytetracycline and rimocidin, two major secondary metabolites of
S. rimosus, was aggravated relative to other cultures during the initial phase of the run with
M. racemosus (
Figure 1a,d). At the final days of the cultivation run, the domination of
M. racemosus over
S. rimosus was ultimately not observed, based on secondary metabolites production (
Figure 1) and the chemical profiles reflected by the TICs (
Figure S2). All in all, the clash between these “worthy microbial opponents” could not be regarded as effective in the context of elevating the levels of secondary metabolites. The issue of a producer strain being overgrown by the accompanying strain that, in principle, should only serve as a stimulating factor, was touched upon in previous efforts [
20,
21,
22]. As the present study was focused on oxytetracycline production, the domination of
S. rimosus over its partners in co-culture was not only not an issue, it was desired to achieve considerable production levels. In co-cultures, there was no visible inhibition exerted by
S. noursei on
S. rimosus in terms of the production capabilities. There was one exception, however.
S. rimosus was not able to dominate the co-culture if it was introduced in the form of spores against the already developed biomass of
S. noursei. The results indicated that granting the 24 h growth advantage to
S. noursei cannot be recommended in oxytetracycline-centered cultivation.
When investigating the production of secondary metabolites in relation to morphological development, the correlations between metabolite levels and morphological characteristics are always sought. One of the behaviors typically observed in past studies involved the decrease in pellet size through morphological engineering, leading in turn to the increase in target product levels (recently reviewed in [
23]). Here, when the morphological results obtained for the “
S. rimosus versus
S. noursei” co-cultivation were compared with the data gathered for other (less effective) co-cultures, there was no evidence that the morphology could be a key factor determining the effectiveness of this variant. It needs to be mentioned that the morphological variations recorded in the present work should be seen as rather subtle when compared with the ones found in our previous studies, e.g., when the high concentration of yeast extract resulted in the transition from the pelleted to loose morphology in
A. terreus [
24] or when the star-shaped pellets were formed in aluminum oxide-supplemented bioreactor cultures of
C. globosum [
17].
To the best of our knowledge, enhancing oxytetracycline production through co-cultivation has not been described before. However, the approach of improving secondary metabolites titers by performing microbial co-cultures was already reported. For example, undecylprodigiosin production by
Streptomyces coelicolor was demonstrated to be boosted by the presence of
E. coli [
22]. However, the co-cultures investigated in the mentioned study were initiated by using the precultures [
22], while in the present study the spores of the stimulating microorganism were applied for inoculation. This is a major difference if one considers the morphological development that occurs during the initial phases of submerged growth of pellet-forming filamentous microorganisms, most importantly the agglomeration events. If the spores of two species are confronted, there is a possibility of spores’ co-agglomeration and building a pellet around the two-species core, as was previously shown for the co-culture of
A. terreus and
C. globosum [
25]. As a result, one of the species is “trapped” inside the pellet with its growth being inhibited by the faster-growing more aggressive dominant partner, while still exerting stimulatory effects from within the filamentous structure. In the present study, the enhancement was also observed when the “young” pellets (after 24 h of growth) of
S. rimosus were contacted with the spores of
S. noursei. When the pellet is developing, its structure is still relatively loose and the increase in size takes place, so the spores of the partnering species can intertwine with the biomass and become the part of the filamentous scaffold that, ultimately, becomes a structural unit responsible for generating the target product. The mechanisms of the interactions that led to the improved oxytetracycline levels were not yet deciphered and therefore are not included in this communication, but further investigation is underway. In addition, a detailed bioprocess analysis focused on the influence of medium composition, pH value, ratio of the number of spores of the participating species, process scale, and the co-cultivation initiation strategies is planned to be performed for the “
S. rimosus + S. noursei” co-culture.
Several recent efforts addressed the regulatory and metabolic mechanisms associated with the enhanced production of antibiotics in
Streptomyces [
26,
27,
28]. Although in the present work, the stimulatory effect of co-cultivation was not considered in the context of molecular studies, one may speculate that the presence of
S. noursei spores and their possible co-agglomeration with
S. rimosus may have an influence on the oxygen levels inside the pellets and the fluxes through central carbon pathways in
S. rimosus that provide the precursors for biosynthetic processes. In the aforementioned technique of microparticle-enhanced cultivation, where the spores of the producing microorganism form co-agglomerates with the supplied mineral particles, the altered availability of oxygen leads to marked metabolic and productivity-related consequences [
29]. Furthermore, it cannot be excluded that the presence of
S. noursei triggers a specific defense response in
S. rimosus that involves the secretion of an antibacterial substance.


{kind=link}
{kind=link}
{kind=link}


