
Increasing Sugar Content in Source for Biofuel Production Using Agrochemical and Genetic Approaches at the Stages of BioMass Preharvesting and Harvesting

 , , ,
, , ,
Abstract
:1. Introduction
Raw Materials Used in the Production of Bioethanol
2. Methods of Increasing Sugar Level
2.1. Pre- and Postharvest Factors
2.2. Genetic Modifications
2.3. Chemical Ripening Methods
2.4. Alternative Methods
3. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ning, P.; Yang, G.; Hu, L.; Sun, J.; Shi, L.; Zhou, Y. Recent advances in the valorization of plant biomass. Biotechnol. Biofuels 2021, 14, 102. [Google Scholar] [CrossRef] [PubMed]
- Zolotareva, D.; Zazybin, A.; Rafikova, K.; Dembitsky, V.M.; Dauletbakov, A.; Yu, V. Ionic liquids assisted desulfurization and denitrogenation of fuels. Vietnam J. Chem. 2019, 55, 133–163. [Google Scholar] [CrossRef]
- Kandpal, R., II; Singh, R. Renewable Energy Sources—A Review. ECS Trans. 2022, 107, 8133. [Google Scholar] [CrossRef]
- Van Heerden, P.D.R.; Donaldson, R.A.; Watt, D.A.; Singels, A. Biomass accumulation in sugarcane: Unravelling the factors underpinning reduced growth phenomena. J. Exp. Bot. 2010, 61, 2877–2887. [Google Scholar] [CrossRef]
- Valladares-Diestra, K.K.; de Souza Vandenberghe, L.P.; Soccol, C.R. A biorefinery approach for pectin extraction and second-generation bioethanol production from cocoa pod husk. Bioresour. Technol. 2022, 346, 126635. [Google Scholar] [CrossRef]
- Chen, W.-H.; Jang, M.-F.; Jheng, S.-L.; Lo, C.-J.; Wang, W. Cellulosic sugars from biomass: Effect of acid presoaking on pretreatment efficiency and operating cost estimation for sugar production. Bioresour. Technol. Rep. 2019, 7, 100259. [Google Scholar] [CrossRef]
- Alonso-Gomez, A.; Bello-Perez, A.L. Four generations of raw materials used for ethanol production: Challenges and opportunities. Agrociencia 2018, 52, 967–990. [Google Scholar]
- Jayaraman, P.; Livingstone, A.; Harikrishnan, S.; Vinoth, S.; Logamba, R. Evaluation of ethanol production using various carbon substrates by Sacharomyces cerevisiae and Schizosacharomyces pombae. J. Pure Appl. Microb. 2017, 11, 1469–1478. [Google Scholar] [CrossRef]
- Jonker, J.G.G.; van der Hilst, F.; Junginger, H.M.; Cavalett, O.; Chagas, M.F.; Faaij, A.P.C. Outlook for ethanol production costs in Brazil up to 2030, for different biomass crops and industrial technologies. Appl. Energy 2015, 147, 593–610. [Google Scholar] [CrossRef]
- Kim, S.M.; Khullar, E.; Liu, W.; Lanahan, M.; Lessard, P.; Dohle, S.; Emery, J.; Raab, R.M.; Singh, V. Rice straw with altered carbohydrate content: Feedstock for ethanol production. ASABE 2015, 58, 523–528. [Google Scholar] [CrossRef]
- Canilha, L.; Chandel, A.K.; Dos Santos Milessi, T.S.; Antunes, F.A.F.; Da Costa Freitas, W.L.; Das Graças Almeida, F.M.; Da Silva, S.S. Bioconversion of sugarcane biomass into ethanol: An overview about composition, pretreatment methods, detoxification of hydrolysates, enzymatic saccharification, and ethanol fermentation. J. Biomed. Biotechnol. 2012, 2012, 1–16. [Google Scholar] [CrossRef]
- Sudhakar, M.P.; Merlyn, R.; Arunkumar, K.; Perumal, K. Characterization, pretreatment and saccharification of spent seaweed biomass for bioethanol production using baker’s yeast. Biomass Bioenergy 2016, 90, 148–154. [Google Scholar] [CrossRef]
- Konda, N.V.S.N.M.; Singh, S.; Simmons, B.A.; Klein-Marcuschamer, D. An Investigation on the Economic Feasibility of Macroalgae as a Potential Feedstock for Biorefineries. Bioenergy Res. 2015, 8, 1046–1056. [Google Scholar] [CrossRef]
- Aro, E.-M. From first generation biofuels to advanced solar biofuels. Ambio 2015, 45, 24–31. [Google Scholar] [CrossRef]
- Srivastava, N.; Srivastava, M.; Mishra, P.K.; Gupta, V.K. Biofuel Production Technologies: Critical Analysis for Sustainability. Clean Energy Production Technologies; Springer: Singapore, 2020. [Google Scholar] [CrossRef]
- Ramos, M.D.N.; Milessi, T.S.; Candido, R.G.; Mendes, A.A.; Aguiar, A. Enzymatic catalysis as a tool in biofuels production in Brazil: Current status and perspectives. Energy Sustain. Dev. 2022, 68, 103–119. [Google Scholar] [CrossRef]
- Huang, H.; Long, S.; Singh, V. Techno-economic analysis of biodiesel and ethanol co-production from lipid producing sugarcane. Biofuels Bioprod. Biorefining 2016, 10, 299–315. [Google Scholar] [CrossRef]
- Awobemi, O.; Kallon, D.V.V.; Onuh, E.I.; Aigbodion, V.S. An Overview of the Classification, Production and Utilization of Biofuels for Internal Combustion Engine Applications. Energies 2021, 14, 5687. [Google Scholar] [CrossRef]
- Siddiqui, H.; Sami, F.; Hayat, S. Glucose: Sweet or bitter effects in plants-a review on current and future perspective. Carbohydr. Res. 2020, 487, 107884. [Google Scholar] [CrossRef]
- Ishmael, U.; Rashid, S.; Basheer, E. Chemical Methods of Pretreatment, Sugar Yields and Economic Costs: A Review. J. Biotechnol. Sci. Res. 2014, 1, 30–38. [Google Scholar]
- An, S.; Li, W.; Liu, Q.; Li, M.; Ma, Q.; Ma, L.; Chang, H. A two-stage pretreatment using acidic dioxane followed by dilute hydrochloric acid on sugar production from corn stover. RSC Adv. 2017, 7, 32452–32460. [Google Scholar] [CrossRef]
- Khan, M.F.S.; Akbar, M.; Xu, Z.; Wang, H. A review on the role of pretreatment technologies in the hydrolysis of lignocellulosic biomass of corn stover. Biomass Bioenergy 2021, 155, 106276. [Google Scholar] [CrossRef]
- Soltanian, S.; Aghbashlo, M.; Almasi, F.; Hosseinzadeh-Bandbafha, H.; Nizami, A.-S.; Sik Ok, Y.; Lam, S.S.; Tabatabaei, M. A critical review of the effects of pretreatment methods on the exergetic aspects of lignocellulosic biofuels. Energy Convers. Manag. 2020, 212, 112792. [Google Scholar] [CrossRef]
- Harmsen, P.; Huijgen, W.; Bermudez, L.; Bakker, R.R. Literature Review of Physical and Chemical Pretreatment Processes for Lignocellulosic Biomass; Wageningen UR Food & Biobased Research: Wageningen, The Netherlands, 2010. [Google Scholar]
- Kniss, A. Long-term trends in the intensity and relative toxicity of herbicide use. Nat. Commun. 2017, 8, 14865. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.S.; Zhou, Y.; Jiang, Z.; Wang, L.E.; Huang, J.J.; Zhang, T.Y.; Zhao, Y.; Shen, W.; Zou, S.H.; Zang, L.L. The in vitro effects of gibberellin on human sperm motility. Aging 2019, 11, 3080–3093. [Google Scholar] [CrossRef]
- Kollerup Madsen, B.; Hilscher, M.; Zetner, D.; Rosenberg, J. Adverse reactions of dimethyl sulfoxide in humans: A systematic review. F1000Research 2018, 7, 1746. [Google Scholar] [CrossRef]
- Sulpice, R.; Flis, A.; Ivakov, A.A.; Apelt, F.; Krohn, N.; Encke, B. Arabidopsis Coordinates the Diurnal Regulation of Carbon Allocation and Growth across a Wide Range of Photoperiods. Mol. Plant 2014, 7, 137–155. [Google Scholar] [CrossRef]
- Chen, X.l.; Wang, L.C.; Li, T.; Yang, Q.C.; Guo, W.Z. Sugar accumulation and growth of lettuce exposed to different lighting modes of red and blue LED light. Sci. Rep. 2019, 9, 6926. [Google Scholar] [CrossRef]
- Thompson, M.; Gamage, D.; Hirotsu, N.; Martin, A.; Seneweera, S. Effects of Elevated Carbon Dioxide on Photosynthesis and Carbon Partitioning: A Perspective on Root Sugar Sensing and Hormonal Crosstalk. Front. Physiol. 2017, 8, 578. [Google Scholar] [CrossRef]
- Zheng, Y.; Li, F.; Hao, L.; Yu, J.; Guo, L.; Zhou, H.; Ma, C. Elevated CO2 concentration induces photosynthetic down-regulation with changes in leaf structure, non-structural carbohydrates and nitrogen content of soybean. BMC Plant Biol. 2019, 19, 255. [Google Scholar] [CrossRef]
- Aranjuelo, I.; Irigoyen, J.J.; Sánchez-Díaz, M.; Nogués, S. Carbon partitioning in N2 fixing Medicago sativa plants exposed to different CO2 and temperature conditions. Funct. Plant Biol. 2008, 35, 306–317. [Google Scholar] [CrossRef]
- Lee-Ho, E.; Walton, L.J.; Reid, D.M.; Yeung, E.C.; Kurepin, L.V. Effects of elevated carbon dioxide and sucrose concentrations on Arabidopsis thaliana root architecture and anatomy. Can. J. Bot. 2007, 85, 324–330. [Google Scholar] [CrossRef]
- Yang, L.Y.; Yang, S.L.; Li, J.Y.; Ma, J.H.; Pang, T.; Zou, C.M.; He, B.; Gong, M. Effects of different growth temperatures on growth, development, and plastid pigments metabolism of tobacco (Nicotiana tabacum L.) plants. Bot. Stud. 2018, 59, 5. [Google Scholar] [CrossRef] [PubMed]
- Mathieu, A.S.; Tinel, C.; Dailly, H.; Quinet, M.; Lutts, S. Impact of high temperature on sucrose translocation, sugar content and inulin yield in Cichorium intybus L. var. sativum. Plant Soil 2018, 432, 273–288. [Google Scholar] [CrossRef]
- Uehara, N.; Sasaki, H.; Aoki, N.; Ohsugi, R. Effects of the Temperature Lowered in the Daytime and Night-time on Sugar Accumulation in Sugarcane. Plant Prod. Sci. 2009, 12, 420–427. [Google Scholar] [CrossRef]
- Badri, M.A.; Minchin, P.E.H.; Lapointe, L. Effects of temperature on the growth of spring ephemerals: Crocus vernus. Physiol. Plant. 2007, 130, 67–76. [Google Scholar] [CrossRef]
- Aluko, O.O.; Li, C.; Wang, Q.; Liu, H. Sucrose Utilization for Improved Crop Yields: A Review Article. Int. J. Mol. Sci. 2021, 22, 4704. [Google Scholar] [CrossRef]
- Liu, X.; Huang, B. Carbohydrate Accumulation in Relation to Heat Stress Tolerance in Two Creeping Bentgrass Cultivars. J. Am. Soc. Hort. Sci. 2000, 125, 442–447. [Google Scholar] [CrossRef]
- Xuan, T.D.; Phuong, N.T.; Khanh, T.D. Effects of Fertilizers on Biomass, Sugar Content and Ethanol Production of Sweet Sorghum. In Biomass Volume Estimation and Valorization for Energy; IntechOpen: London, UK, 2017. [Google Scholar] [CrossRef]
- Morrow, J.L.; Sa, P.T.; Beattie, G.A.C.; Milham, P.J.; Riegler, M.; Spooner-Hart, R.; Holford, P. Additions of sugar and nitrogenous fertiliser affect plant nitrogen status and soil microbial communities. Appl. Soil Ecol. 2019, 139, 47–55. [Google Scholar] [CrossRef]
- Barłóg, P.; Szczepaniak, W.; Grzebisz, W.; Pogłodziński, R. Sugar beet response to different K, Na and Mg ratiosin applied fertilizers. Plant Soil Environ. 2018, 64, 173–179. [Google Scholar] [CrossRef]
- Levitt, G.; Smith, W.F. Method of Increasing Sugar Content in Sugarcane and Sorghum. U.S. Patent 4,362,549, 7 December 1982. [Google Scholar]
- Zarzecka, K.; Gugala, M.; Mystkowska, I.; Baranowska, A.; Sikorska, A. Effect of herbicides on the content dry matter and sugars in edible potato tubers. Rom. Agric. Res. 2017, 34, 371–375. [Google Scholar]
- Mikkelsen, D.S.; Griffith, R.B.; Ririe, D. Sugar Beet Response to Maleic Hydrazide Treatment. Agron. J. 1952, 44, 533–536. [Google Scholar] [CrossRef]
- Sabba, R.P.; Holman, P.; Drilias, M.J.; Bussan, A.J. Influence of maleic hydrazide on yield and sugars in Atlantic, freedom russet and White Pearl potato tubers. Am. J. Potato Res. 2009, 86, 272–277. [Google Scholar] [CrossRef]
- He, H.; Zhiting, X.; Minjing, L.; Shuanglian, X.; Shenglan, L.; Mba, F.O. Effect of cadmium and herbicides on the growth, chlorophyll and soluble sugar content in rice seedlings. Wuhan Univ. J. Nat. Sci. 2006, 11, 742–748. [Google Scholar] [CrossRef]
- Bickers, U.; Lehr, S.; Bonfig-Picard, G.; Schmidt, M.; Hills, M.; Moreno, J.P.R.-S.; Hacker, E. Use of Quinoline Derivatives for Improving Plant Yield. U.S. Patent No. WO2014135481, 12 September 2019. [Google Scholar]
- Nasonova, D. Kagatnik is an effective drug for increasing the sugar content and preservation of sugar beet roots. Plant Prot. Quar. 2014, 8, 42–43. (In Russian) [Google Scholar]
- Darko, E.; Ve’gh, B.; Khalil, R.; Marček, T.; Szalai, G.; Pa’l, M. Metabolic responses of wheat seedlings to osmotic stress induced by various osmolytes under iso-osmotic conditions. PLoS ONE 2019, 14, e0226151. [Google Scholar] [CrossRef]
- Nemati, I.; Moradi, F.; Gholizadeh, S.; Esmaeili, M.A.; Bihamta, M.R. The effect of salinity stress on ions and soluble sugars distribution in leaves, leaf sheaths and roots of rice (Oryza sativa L.) seedlings. Plant Soil Environ. 2011, 57, 26–33. [Google Scholar] [CrossRef]
- Amirjani, M.R. Effect of Salinity Stress on Growth, Sugar Content, Pigments and Enzyme Activity of Rice. Int. J. Bot. 2011, 7, 73–81. [Google Scholar] [CrossRef]
- Lemoine, R.; La Camera, S.; Atanassova, R.; Dédaldéchamp, F.; Allario, T.; Pourtau, N. Source-to-sink transport of sugar and regulation by environmental factors. Front. Plant Sci. 2013, 4, 272. [Google Scholar] [CrossRef]
- Javid, M.G.; Sorooshzadeh, A.; Sanavy, S.A.M.M.; Allahdadi, I.; Moradi, F. Effects of the exogenous application of auxin and cytokinin on carbohydrate accumulation in grains of rice under salt stress. Plant Growth Regul. 2011, 65, 305–313. [Google Scholar] [CrossRef]
- Duan, Y.; Sangani, C.B.; Muddassir, M.; Soni, K.V. Copper, Chromium and Nickel Heavy Metal Effects on Total Sugar and Protein Content in Glycine Max; Research Square: Durham, NC, USA, 2020. [Google Scholar]
- Fryzova, R.; Pohanka, M.; Martinkova, P.; Cihlarova, H.; Brtnicky, M.; Hladky, J.; Kynicky, J. Oxidative Stress and Heavy Metals in Plants. Rev. Environ. Contam. Toxicol. 2017, 245, 129–156. [Google Scholar] [CrossRef]
- Jacobs, R.; Schweiger, R.; Muller, C. Aphid infestation leads to plant part-specific changes in phloem sap chemistry, which may indicate niche construction. New Phytol. 2018, 221, 503–514. [Google Scholar] [CrossRef] [PubMed]
- Agudelo-Romero, P.; Erban, A.; Rego, C. Transcriptome and metabolome reprogramming in Vitis vinifera cv. Trincadeira berries upon infection with Botrytis cinerea. J. Exp. Bot. 2015, 66, 1769–1785. [Google Scholar] [CrossRef] [PubMed]
- Lima, M.F.; Eloy, N.B.; Batista de Siqueira, J.A.; Inze, D.; Hemerly, A.S.; Ferreira, P.C.G. Molecular mechanisms of biomass increase in plants. Biotechnol. Res. Innov. 2017, 1, 14–25. [Google Scholar] [CrossRef]
- Jha, Y.; Subramanian, R.B. Effect of Root-Associated Bacteria on Soluble Sugar Metabolism in Plant Under Environmental Stress. In Plant Metabolites and Regulation Under Environmental Stress; Academic Press: Cambridge, MA, USA, 2018; pp. 231–240. [Google Scholar] [CrossRef]
- Baltazari, A.; Mtui, H.D.; Mwatawala, M.W.; Chove, L.M.; Msogoya, T.; Samwel, J.; Subramanian, J. Effects of Storage Conditions, Storage Duration and Post-Harvest Treatments on Nutritional and Sensory Quality of Orange (Citrus sinensis (L.) Osbeck) Fruits. Int. J. Fruit Sci. 2019, 20, 737–749. [Google Scholar] [CrossRef]
- Godoy, J.G.V.; Tesso, T.T. Analysis of Juice Yield, Sugar Content, and Biomass Accumulation in Sorghum. Crop Sci. 2013, 53, 1288–1297. [Google Scholar] [CrossRef]
- Hoang, N.V.; Furtado, A.; Botha, F.C.; Simmons, B.A.; Henry, R.J. Potential for Genetic Improvement of Sugarcane as a Source of Biomass for Biofuels. Front. Bioeng. Biotechnol. 2015, 3, 182. [Google Scholar] [CrossRef]
- Wu, L.; Birch, R.G. Doubled sugar content in sugarcane plants modified to produce a sucrose isomer. Plant Biotechnol. J. 2007, 5, 109–117. [Google Scholar] [CrossRef]
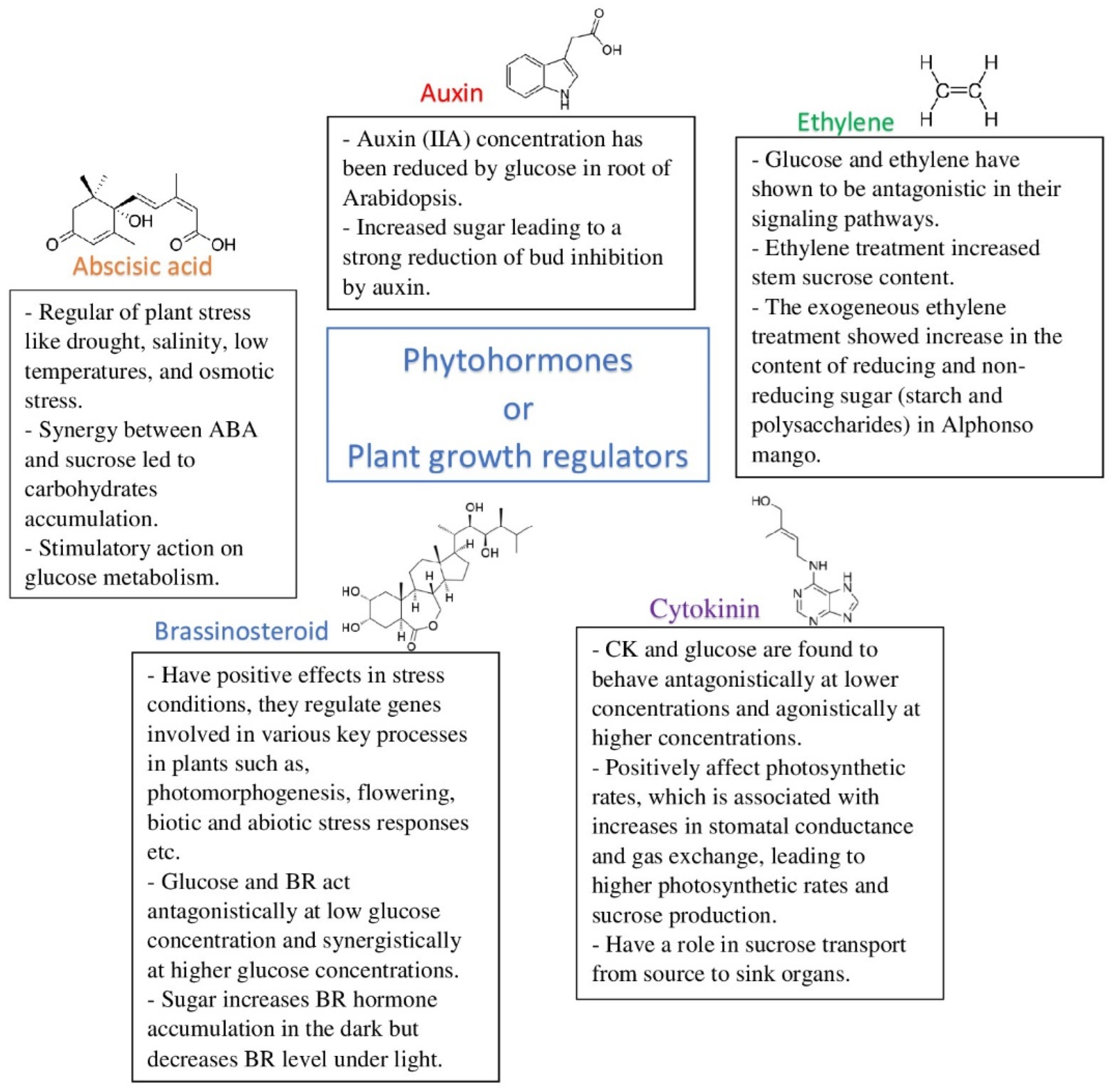
- Vanstraelen, M.; Benkova, E. Hormonal Interactions in the Regulation of Plant Development. Annu. Rev. Cell Dev. Biol. 2012, 28, 463–487. [Google Scholar] [CrossRef]
- Sah, S.K.; Reddy, K.R.; Li, J. Abscisic Acid and Abiotic Stress Tolerance in Crop Plants. Front. Plant Sci. 2016, 7, 571. [Google Scholar] [CrossRef]
- Nambara, E. Abscidic acid. Regulators of growth. In Encyclopedia of Applied Plant Sciences, 2nd ed.; Elsevier Ltd.: Toronto, ON, Canada, 2017; Volume 1, pp. 361–366. [Google Scholar] [CrossRef]
- Chen, T.; Li, G.; Islam, M.R.; Fu, W.; Feng, B.; Tao, L.; Fu, G. Abscisic acid synergizes with sucrose to enhance grain yield and quality of rice by improving the source-sink relationship. BMC Plant Biol. 2019, 19, 525. [Google Scholar] [CrossRef]
- Kobashi, K.; Sugaya, S.; Gemma, H.; Iwahori, S. Effect of abscisic acid (ABA) on sugar accumulation in the flesh tissue of peach fruit at the start of the maturation stage. Plant Growth Regul. 2001, 35, 215–223. [Google Scholar] [CrossRef]
- Ghate, T.; Barvkar, V.; Deshpande, S.; Bhargava, S. Role of ABA Signaling in Regulation of Stem Sugar Metabolism and Transport under Post-Flowering Drought Stress in Sweet Sorghum. Plant Mol. Biol. Rep. 2019, 37, 303–313. [Google Scholar] [CrossRef]
- LeClere, S.; Schmelz, E.A.; Chourey, P.S. Sugar Levels Regulate Tryptophan-Dependent Auxin Biosynthesis in Developing Maize Kernels. Plant Physiol. 2010, 153, 306–318. [Google Scholar] [CrossRef] [PubMed]
- Bertheloot, J.; Barbier, F.; Boudon, F.; Perez-Garcia, M.D.; Peron, T.; Citerne, S. Sugar availability suppresses the auxin-induced strigolactone pathway to promote bud outgrowth. New Phytol. 2020, 225, 866–879. [Google Scholar] [CrossRef] [PubMed]
- Das, P.K.; Shin, D.H.; Choi, S.-B.; Yoo, S.-D.; Choi, G.; Park, Y.-I. Cytokinins enhance sugar-induced anthocyanin biosynthesis in Arabidopsis. Mol. Cells 2012, 34, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Le Gourrierec, J.; Jiao, F.; Demotes-Mainard, S.; Perez-Garcia, M.-D.; Ogé, L. Convergence and Divergence of Sugar and Cytokinin Signaling in Plant Development. Int. J. Mol. Sci. 2021, 22, 1282. [Google Scholar] [CrossRef]
- McIntyre, K.E.; Bush, D.R.; Argueso, C.T. Cytokinin Regulation of Source-Sink Relationships in Plant-Pathogen Interactions. Front. Plant Sci. 2021, 12, 677585. [Google Scholar] [CrossRef]
- Chen, Z.; Qin, C.; Wang, M.; Liao, F.; Liao, Q.; Liu, X.; Huang, D. Ethylene-mediated improvement in sucrose accumulation in ripening sugarcane involves increased sink strength. BMC Plant Biol. 2019, 19, 285. [Google Scholar] [CrossRef]
- Nishikawa, F.; Iwama, T.; Kato, M.; Hyodo, H.; Ikoma, Y.; Yano, M. Effect of sugars on ethylene synthesis and responsiveness in harvested broccoli florets. Postharvest Biol. Technol. 2005, 36, 157–165. [Google Scholar] [CrossRef]
- Chidley, H.G.; Deshpande, A.B.; Oak, P.S.; Pujari, K.H.; Giri, A.P.; Gupta, V.S. Effect of postharvest ethylene treatment on sugar content, glycosidase activity and its gene expression in mango fruit. J. Sci. Food Agric. 2016, 97, 1624–1633. [Google Scholar] [CrossRef]
- Jeong, S.-W.; Das, P.K.; Jeoung, S.C.; Song, J.-Y.; Lee, H.K.; Kim, Y.-K.; Kim, W.J.; Park, Y.I.; Yoo, S.-D.; Choi, S.-B.; et al. Ethylene Suppression of Sugar-Induced Anthocyanin Pigmentation in Arabidopsis. Plant Physiol. 2010, 154, 1514–1531. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.; Singh, M.; Laxmi, A. Multiple Interactions between Glucose and Brassinosteroid Signal Transduction Pathways in Arabidopsis Are Uncovered by Whole-Genome Transcriptional Profiling. Plant Physiol. 2015, 168, 1091–1105. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; He, J. Sugar-induced plant growth is dependent on brassinosteroids. Plant Signal. Behav. 2015, 10, e1082700. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Sun, Y.; Jiang, X.; Wang, W.; Wang, Z.-Y. Sugar inhibits brassinosteroid signaling by enhancing BIN2 phosphorylation of BZR1. PLoS. Genet. 2021, 17, e1009540. [Google Scholar] [CrossRef] [PubMed]
- Dong, H.; Liu, J.; He, G.; Liu, P.; Sun, J. Photoexcited phytochrome B interacts with brassinazole resistant 1 to repress brassinosteroid signaling in Arabidopsis. J. Integr. Plant Biol. 2020, 62, 652–667. [Google Scholar] [CrossRef]
- Liang, T.; Mei, S.; Shi, C.; Yang, Y.; Peng, Y.; Ma, L. UVR8 Interacts with BES1 and BIM1 to Regulate Transcription and Photomorphogenesis in Arabidopsis. Dev. Cell. 2018, 44, 512–523.e5. [Google Scholar] [CrossRef]
- Hansch, F.; Jaspar, H.; von Sivers, L.; Bitterlich, M.; Franken, P.; Kühn, C. Brassinosteroids and sucrose transport in mycorrhizal tomato plants. Plant Signal. Behav. 2020, 15, 2. [Google Scholar] [CrossRef]
- Van Heerden, P.D.; Eggleston, G.; Donaldson, R.A. Ripening and postharvest deterioration. In Sugarcane: Physiology, Biochemistry, and Functional Biology; Moore, P.H., Botha, F.C., Eds.; Wiley: Blackwell, UK, 2013; pp. 55–84. [Google Scholar] [CrossRef]
- Roberto, G.G.; Cunha, C.; Sales, C.R.; Silveira, N.M.; Ribeiro, R.V.; Machado, E.C.; Lagôa, A.M. Variation of photosynthesis and carbohydrate levels induced by ethephon and water deficit on the ripening stage of sugarcane. Bragantia 2015, 74, 379–386. [Google Scholar] [CrossRef]
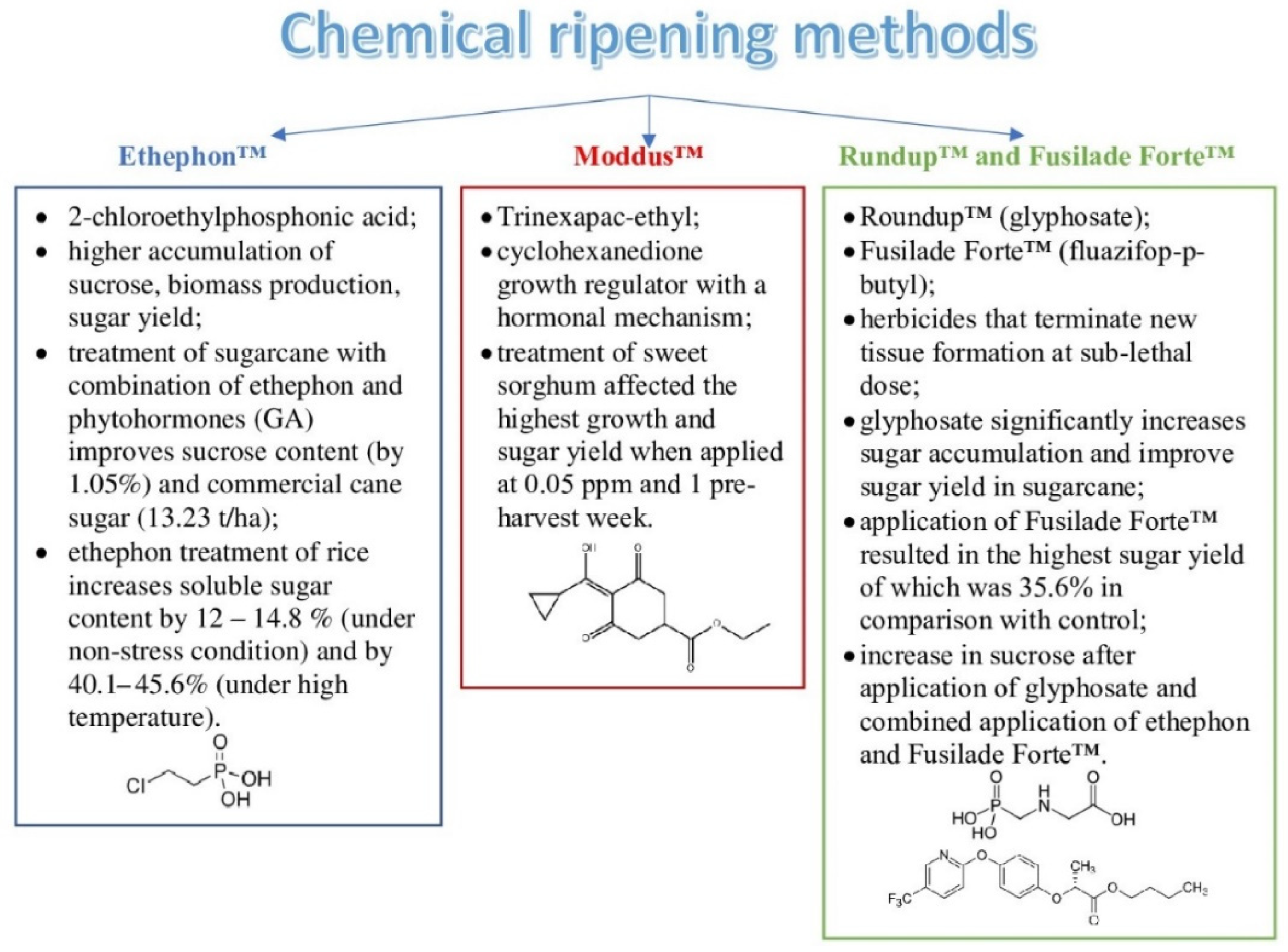
- Li, Y.; Solomon, S. Ethephon: A versatile growth regulator for sugar cane industry. Sugar Tech. 2003, 5, 213–223. [Google Scholar] [CrossRef]
- Praharaj, S.; Singh, D.; Guru, S.K. Yield and juice quality of sugarcane as influenced by ethephon and gibberellic acid. Int. J. Curr. Microbiol. Appl. Sci. 2018, 7, 1647–1652. [Google Scholar] [CrossRef]
- Gautam, H.; Fatma, M.; Sehar, Z.; Iqbal, N.; Albaqami, M.; Khan, N.A. Exogenously-sourced ethylene positively modulates photosynthesis, carbohydrate metabolism, and antioxidant defense to enhance heat tolerance in Rice. Int. J. Mol. Sci. 2022, 23, 1031. [Google Scholar] [CrossRef] [PubMed]
- Van Heerden, P.D.R.; Mbatha, T.P.; Ngxaliwe, S. Chemical ripening of sugarcane with TRINEXAPAC-Ethyl (Moddus®)—Mode of action and comparative efficacy. Field Crops Res. 2015, 181, 69–75. [Google Scholar] [CrossRef]
- Yoosukyingsataporn, S.; Detpiratmongkol, S. Influences of trinexapac-ethyl on development and sugar content of sorghum bicolor. Int. J. Agric. Technol. 2019, 15, 1053–1062. [Google Scholar]
- Su, L.Y.; Dela Cruz, A.; Moore, P.H.; Maretzki, A. The relationship of glyphosate treatment to sugar metabolism in sugarcane: New physiological insights. J. Plant Physiol. 1992, 140, 168–173. [Google Scholar] [CrossRef]
- Kouamé, K.D.; Péné, B.C.; N’guessan, A.C.; Boua, B.M.; Ouattara, Y.; Zouzou, M. Effect of glyphosate used as a sugarcane chemical ripener in Cote d’Ivoire. Afr. J. Plant Sci. 2017, 11, 341–350. [Google Scholar] [CrossRef]
- Muhwiridzwa, M.; Manenji, B.; Madanzi, T.; Mahohoma, W. Evaluating the effects of chemical ripening with fluazifop-p-butyl on sugarcane (saccharum officinarum) yield and sugar content. Int. J. Plant Soil Sci. 2016, 9, 1–8. [Google Scholar] [CrossRef]
- Morgan, T.; Jackson, P.; McDonald, L.; Holtum, J. Chemical ripeners increase early season sugar content in a range of sugarcane varieties. Aust. J. Agric. Res. 2007, 58, 233. [Google Scholar] [CrossRef]
- Ayele, N.; Tana, T.; Van Heerden, P.D.; Tsadik, K.W.; Alemayehu, Y. Ripening response of sugarcane varieties to chemical ripeners and economic benefits during the early period of harvesting at Wonji-Shoa and Metahara sugarcane plantations, Central Rift Valley of Ethiopia. Int. J. Agron. 2021, 2021, 1–9. [Google Scholar] [CrossRef]
- Bhonsle, N.; Kheraliya, S. Super Absorbent Polymer and a Method of Increasing Sugar Content in Plants. U.S. Patent No. WO2020202092, 8 October 2020. [Google Scholar]
- Islam, M.R.; Xue, X.; Mao, S.; Zhao, X.; Eneji, A.E.; Hu, Y. Superabsorbent polymers (SAP) enhance efficient and eco-friendly production of corn (Zea mays L.) in drought affected areas of northern China. Afr. J. Biotechnol. 2011, 10, 4887–4894. [Google Scholar] [CrossRef]
- Pačuta, V.; Rašovský, M.; Michalska-Klimczak, B.; Wyszyňski, Z. Impact of Superabsorbent Polymers and Variety on Yield, Quality and Physiological Parameters of the Sugar Beet (Beta vulgaris prov. Altissima Doell). Plants 2021, 10, 757. [Google Scholar] [CrossRef]
- Vlasyuk, P.; Prihodko, N.; Borisyuk, V.; Bystryi, V.; Kushitsky, M.; Zub, M. Methods for Increasing the Yield and Sugar Content of Sugar. Beet. Patent No. SU444532A1, 30 September 1974. (In Russian). [Google Scholar]
- Tretyakov, G.; Tretyakova, Y.; Tretyakov, A. Sugar Beet Processing. Method. Patent No. RU2286673, 10 November 2006. (In Russian). [Google Scholar]
- Murakami, N.T.; Kurimura, H.; Kirimura, K.; Kino, K.; Usami, S. Continuous degradation of dimethyl sulfoxide to sulfate ion by Hyphomicrobium denitrificans WUK217. J. Biosci. Bioeng. 2002, 94, 52–56. [Google Scholar] [CrossRef]
- Kumar, B.; Gangwar, M.S.; Rathore, V.S. Effect of dimethylsulfoxide (DMSO) on zinc availability (L-value), growth and metabolic activities of rice plants. Plant Soil 1976, 45, 235–246. [Google Scholar] [CrossRef]
- Zhang, X.-H.; Yu, X.-Z.; Yue, D.-M. Phytotoxicity of dimethyl sulfoxide (DMSO) to Rice Seedlings. Int. J. Environ. Sci. Technol. 2015, 13, 607–614. [Google Scholar] [CrossRef]
- Ma, L.; Lin, Q.; Song, Y.; Zhao, B.; Fan, M. Toxic effect of three imidazole ionic liquids on two terrestrial plants. Open Life Sci. 2020, 15, 466–475. [Google Scholar] [CrossRef] [PubMed]
- Bernot, R.J.; Brueseke, M.A.; Evans-White, M.A.; Lamberti, G.A. Acute and chronic toxicity of imidazolium-based ionic liquids on Daphnia Magna. Environ. Toxicol. Chem. 2005, 24, 87. [Google Scholar] [CrossRef]
- Bubalo, M.C.; Radošević, K.; Redovniković, I.R.; Slivac, I.; Srček, V.G. Toxicity mechanisms of ionic liquids. Arh. Hyg. Rada Toxicol. 2017, 68, 171–179. [Google Scholar] [CrossRef]
- Parthasarathi, R.; Sun, J.; Dutta, T. Activation of lignocellulosic biomass for higher sugar yields using aqueous ionic liquid at low severity process conditions. Biotechnol. Biofuels 2016, 9, 160. [Google Scholar] [CrossRef]
- Hassan El-Sayed, R.E.; Mutelet, F.; Moise, J.-C. The pretreatment of miscanthus using ionic liquids; a way for biofuel production. MATEC Web Conf. 2013, 3, 01049. [Google Scholar] [CrossRef]
- Hassan El-Sayed, R.E.; Mutelet, F.; Moise, J.-C.; Brosse, N. Pretreatment of miscanthus using 1,3-dimethyl-imidazolium methyl phosphonate (DMIMMPh) ionic liquid for glucose recovery and ethanol production. RSC Adv. 2015, 5, 61455–61464. [Google Scholar] [CrossRef]
- Socha, A.; Parthasarathi, R.; Shi, J.; Pattathil, S.; Whyte, D.; Bergeron, M.; George, A.; Tran, K.; Stavila, V.; Venkatachalam, S.; et al. Efficient biomass pretreatment using ionic liquids derived from lignin and hemicellulose. Proc. Natl. Acad. Sci. USA 2014, 111, E3587–E3595. [Google Scholar] [CrossRef]
- Jain, R.; Chandra, A.; Solomon, S. Impact of Exogenously Applied Enzymes Effectors on Sucrose Metabolizing Enzymes (SPS, SS and SAI) and Sucrose Content in Sugarcane. Sugar Tech. 2013, 15, 370–378. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Generation | Pretreatment Process | Raw Material |
|---|---|---|
| First (1G) | Need only milling, fermentation, distillation, and denaturalization (in case of human consumption). For use in mixtures with gasoline, the material must be dehydrated [7]. Fermentation by microorganisms Saccharomyces, Zymomonas, Kluyveromyces, and Zygosaccharomyces [8]. | Sugarcane, sugar beets, and sweet sorghum [9], high-starch content plants (cereals, tubers, and rhizomes) [10]. |
| Second (2G) | Pretreatments, enzyme hydrolysis and fermentation [7,11]. | Farm residue (cereal straw, leaves, dry branches of forest crops) or industrial residues (sugarcane bagasse and DDGS (distillers’ dried grain with solubles)) [7]. |
| Third (3G) | Pretreatments, enzyme hydrolysis, and fermentation [12]. | Perennial grasses, micro- and macro-algae, and cyanobacteria [13]. |
| Fourth (4G) | No-destruction of biomass, direct conversion of solar energy to fuel [14]. Acid/enzymatic hydrolysis, fermentation [15,16]. | Genetically or metabolically modified organisms (GMO), for example, GM sugarcane [17], algae [18]. |
| № | Method | Short Description |
|---|---|---|
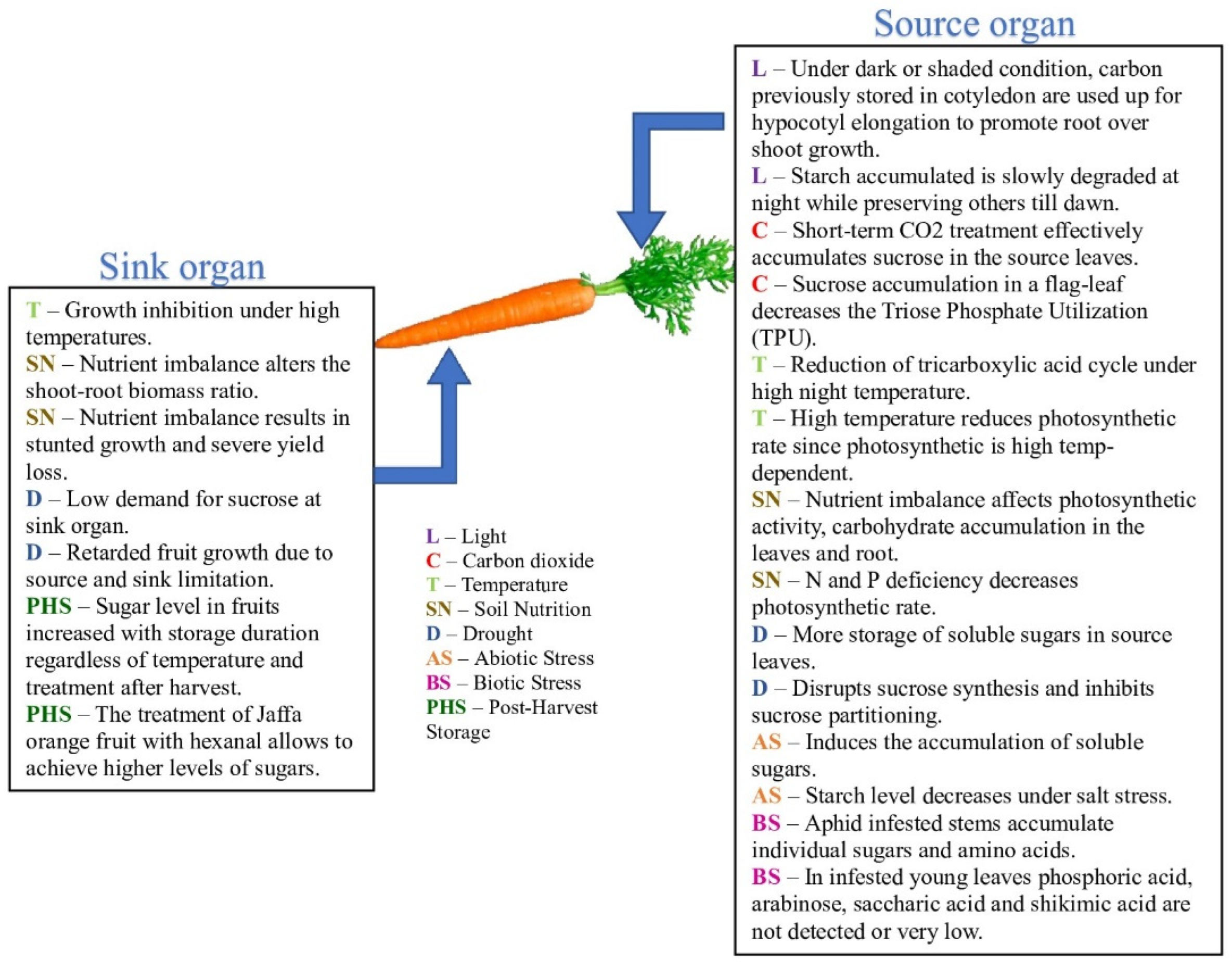
| 1 | Pre- and postharvest factors | Light |
| Carbon dioxide | ||
| Temperature | ||
| Soil nutrition | ||
| Abiotic stress | ||
| Biotic stress | ||
| Postharvest storage | ||
| 2 | Genetic modifications | Traditional breeding |
| Molecular genetic approaches | ||
| NGS (next-generation sequencing) technology | ||
| Phytohormones | ||
| 3 | Chemical ripening methods | Ethephon™ (2-chloroethylphosphonic acid) |
| Moddus™ (Trinexapac-ethyl) | ||
| Roundup™ (glyphosate) | ||
| Fusilade Forte™ (fluazifop-p-butyl) | ||
| 4 | Alternative methods | Superabsorbent polymer (SAP) |
| DMSO | ||
| DMSO/titanium tetrachloride | ||
| Ionic liquids | ||
| Enzyme effectors |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zolotareva, D.; Zazybin, A.; Belyankova, Y.; Dauletbakov, A.; Tursynbek, S.; Rafikova, K.; Ten, A.; Yu, V.; Bayazit, S.; Basharimova, A.; et al. Increasing Sugar Content in Source for Biofuel Production Using Agrochemical and Genetic Approaches at the Stages of BioMass Preharvesting and Harvesting. Molecules 2022, 27, 5210. https://doi.org/10.3390/molecules27165210
Zolotareva D, Zazybin A, Belyankova Y, Dauletbakov A, Tursynbek S, Rafikova K, Ten A, Yu V, Bayazit S, Basharimova A, et al. Increasing Sugar Content in Source for Biofuel Production Using Agrochemical and Genetic Approaches at the Stages of BioMass Preharvesting and Harvesting. Molecules. 2022; 27(16):5210. https://doi.org/10.3390/molecules27165210
Chicago/Turabian StyleZolotareva, Darya, Alexey Zazybin, Yelizaveta Belyankova, Anuar Dauletbakov, Saniya Tursynbek, Khadichahan Rafikova, Assel Ten, Valentina Yu, Sarah Bayazit, Anna Basharimova, and et al. 2022. "Increasing Sugar Content in Source for Biofuel Production Using Agrochemical and Genetic Approaches at the Stages of BioMass Preharvesting and Harvesting" Molecules 27, no. 16: 5210. https://doi.org/10.3390/molecules27165210
APA StyleZolotareva, D., Zazybin, A., Belyankova, Y., Dauletbakov, A., Tursynbek, S., Rafikova, K., Ten, A., Yu, V., Bayazit, S., Basharimova, A., & Aydemir, M. (2022). Increasing Sugar Content in Source for Biofuel Production Using Agrochemical and Genetic Approaches at the Stages of BioMass Preharvesting and Harvesting. Molecules, 27(16), 5210. https://doi.org/10.3390/molecules27165210