

Mass Spectrometric Proof of Predicted Peptides: Novel Adipokinetic Hormones in Insects


Abstract
:1. Introduction
2. Results
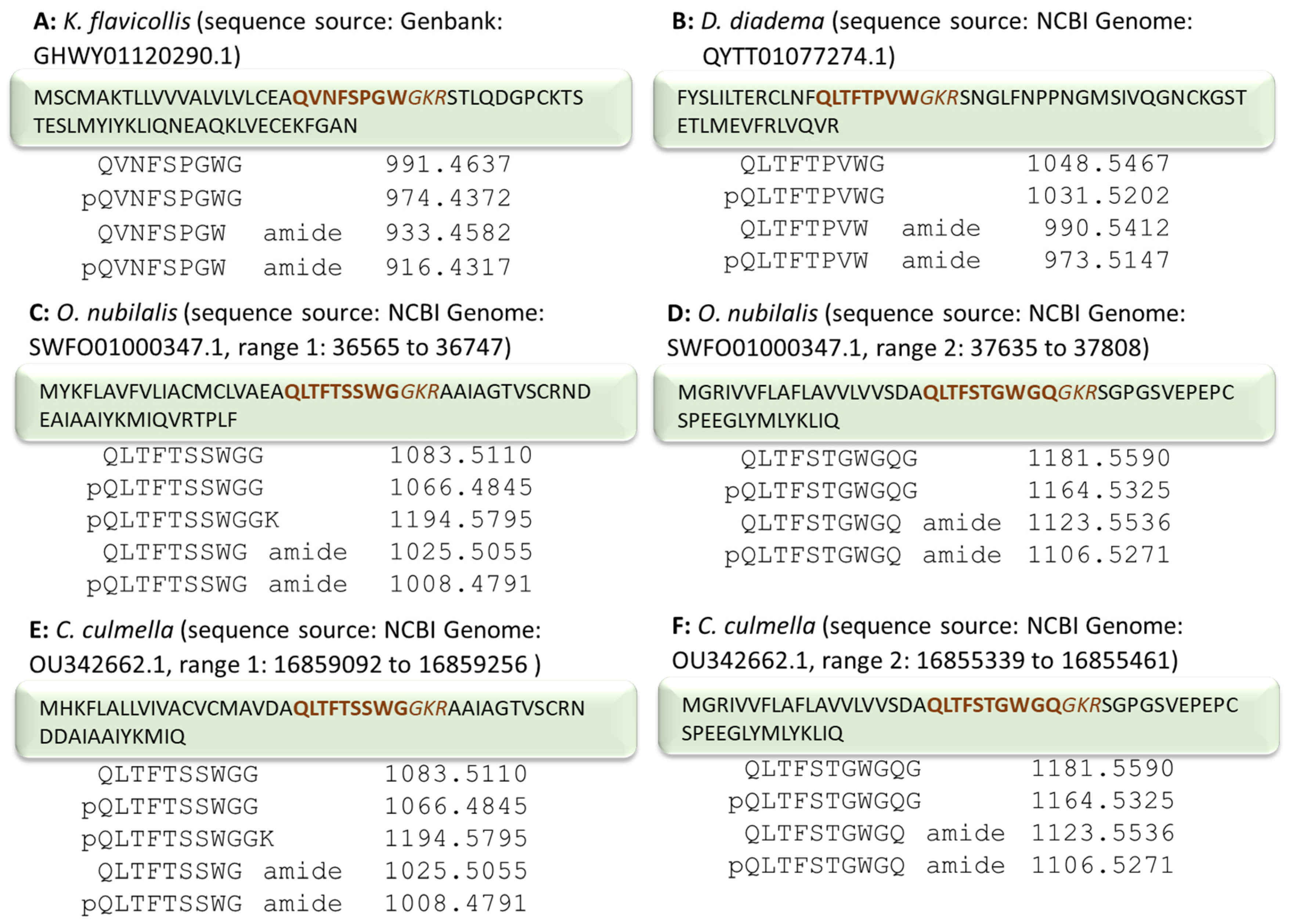
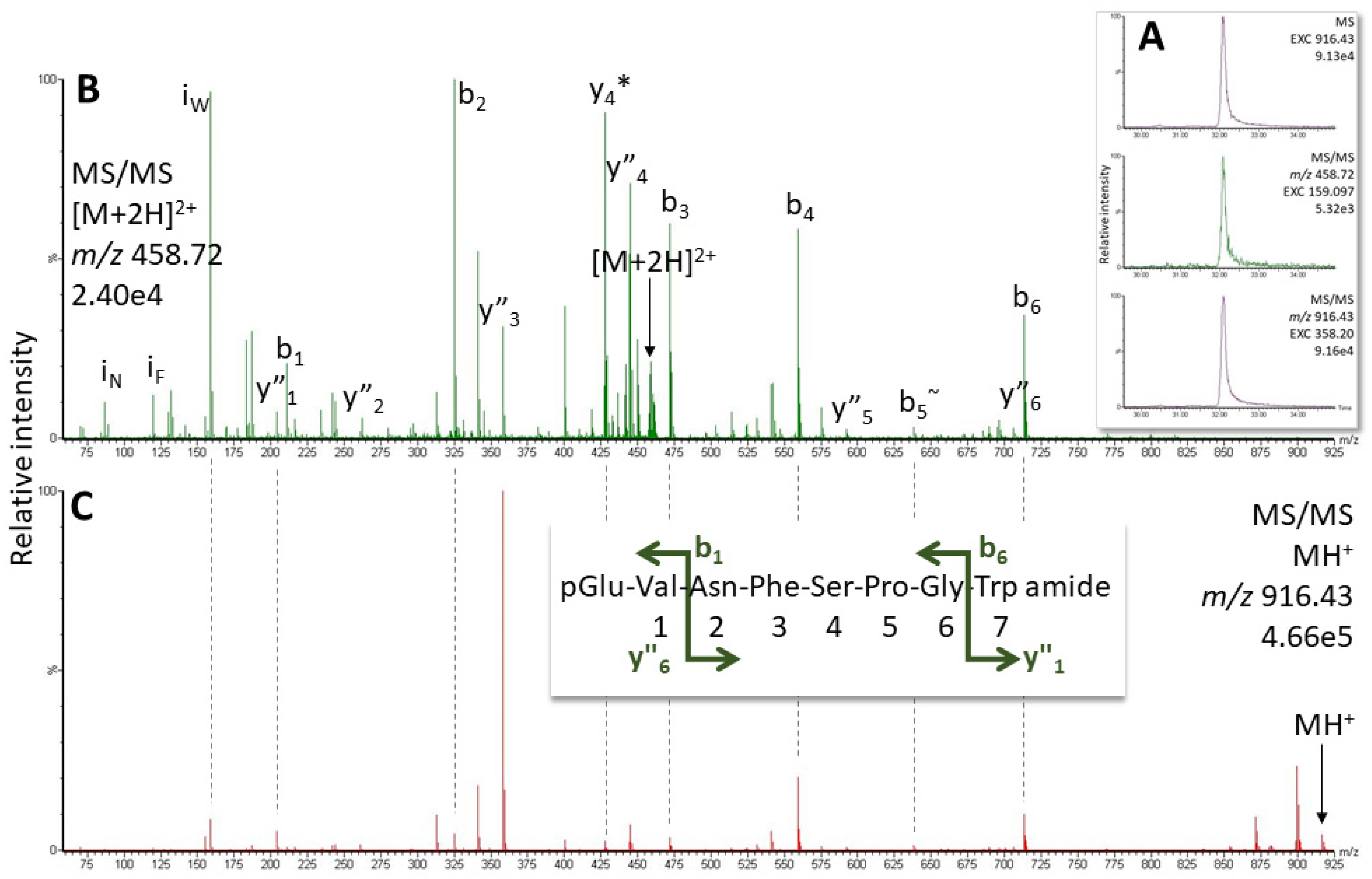
2.1. AKH of the Termite Kalotermes flavicollis, Family Kalotermitidae
2.2. AKHs of Flies (Diptera)
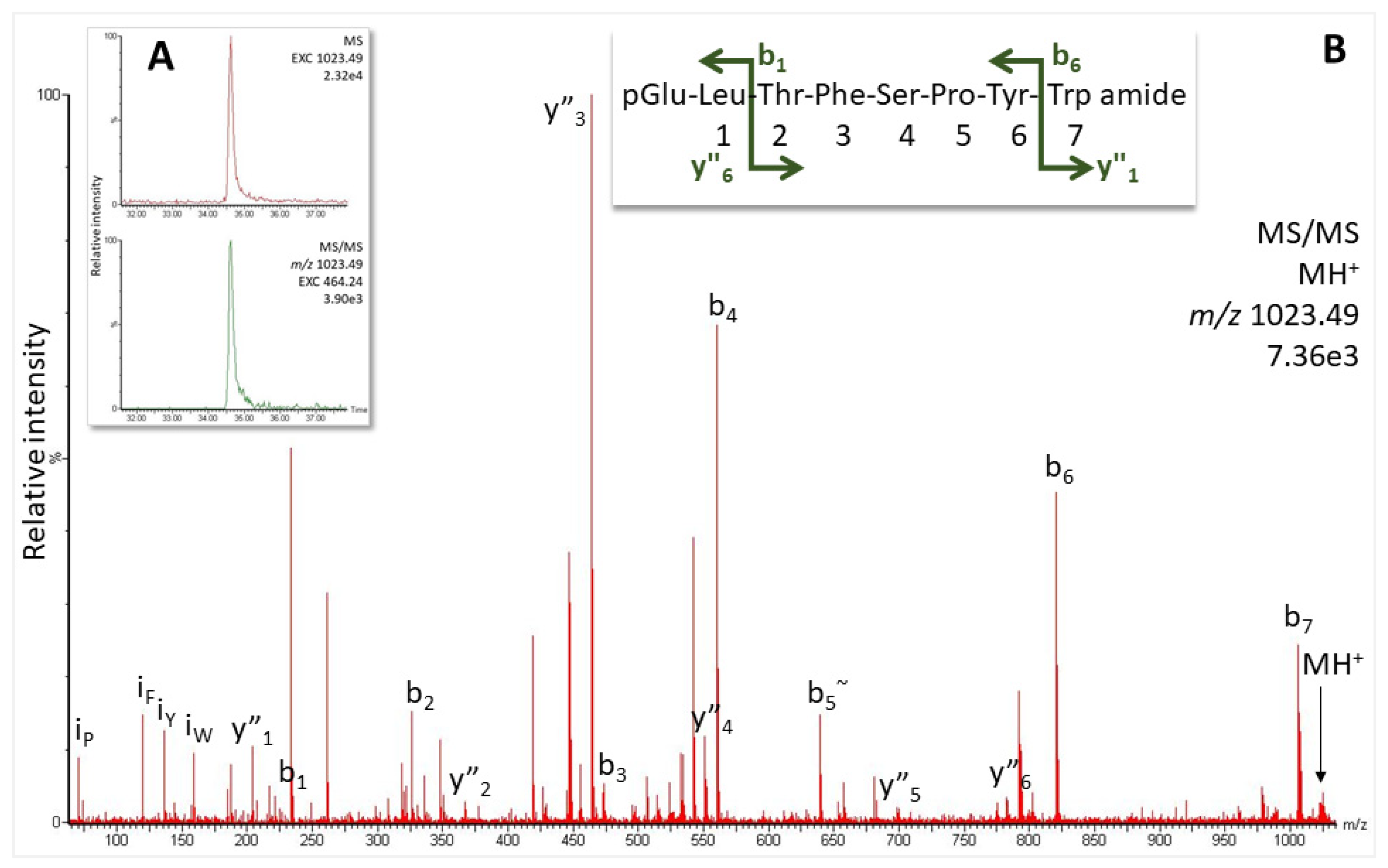
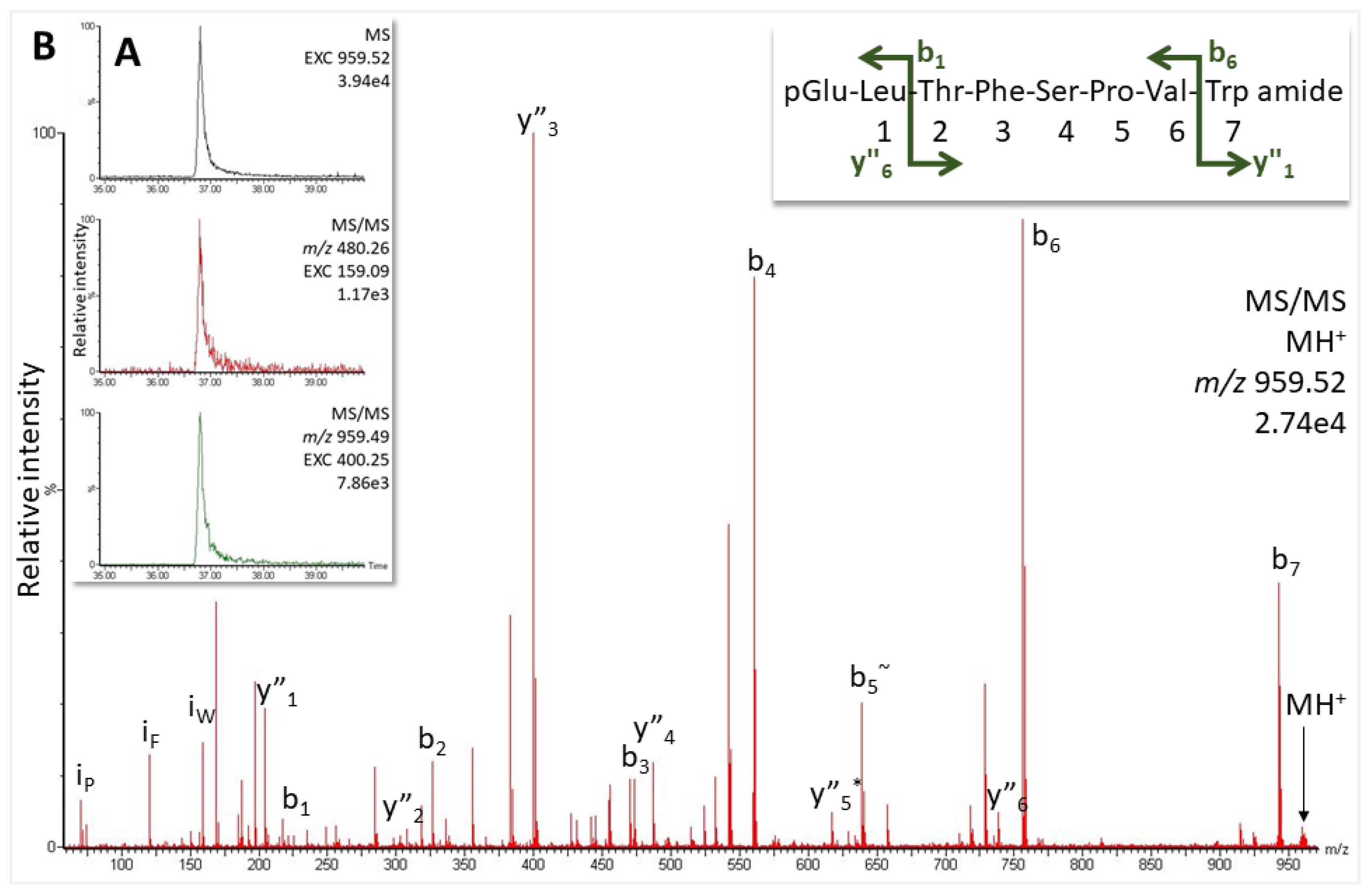
2.2.1. Robber Fly Pegesimallus tapulus, Family Asilidae
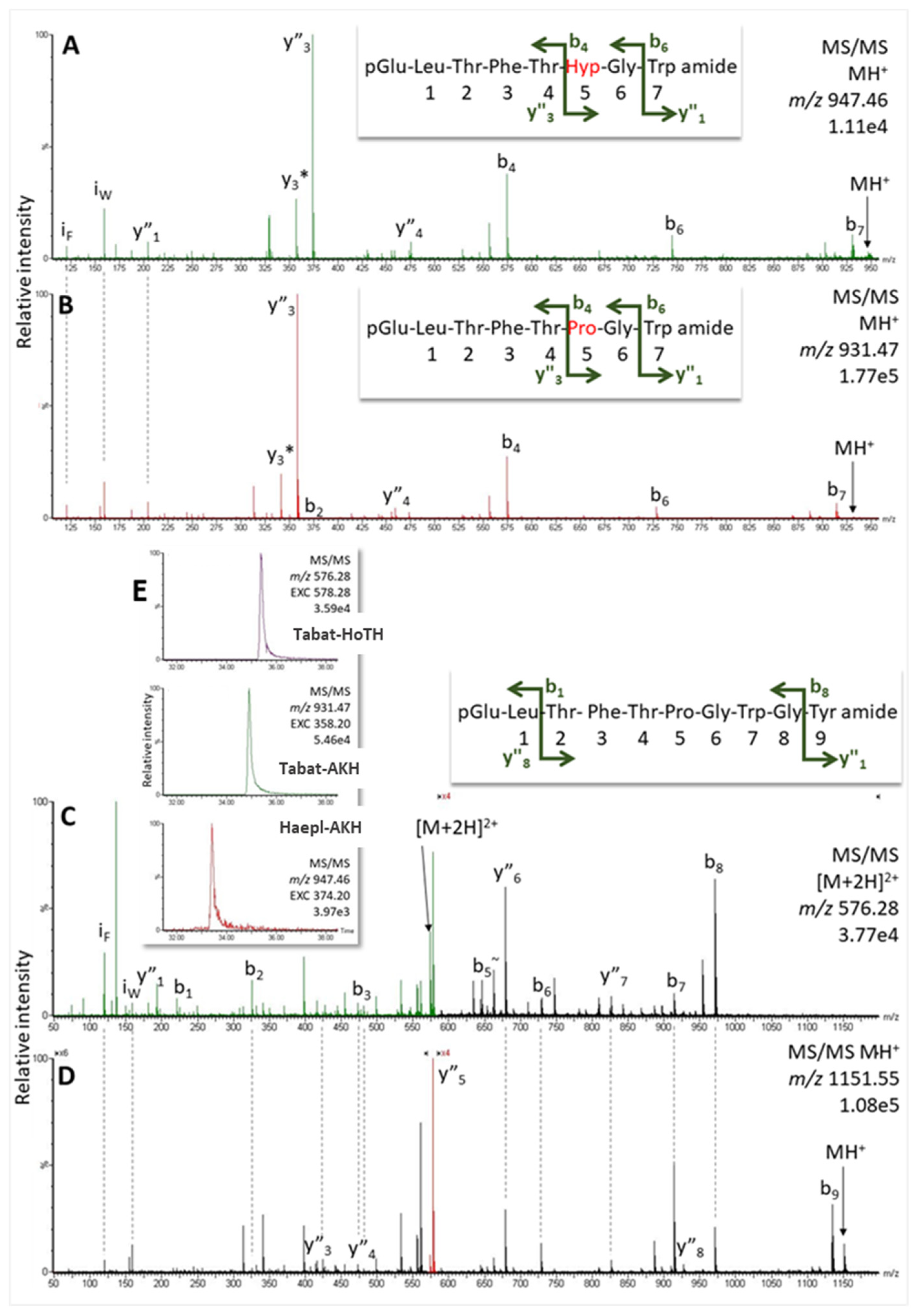
2.2.2. Horse Fly Haematopota pluvialis, Family Tabanidae
2.3. AKHs of Moths (Lepidoptera)
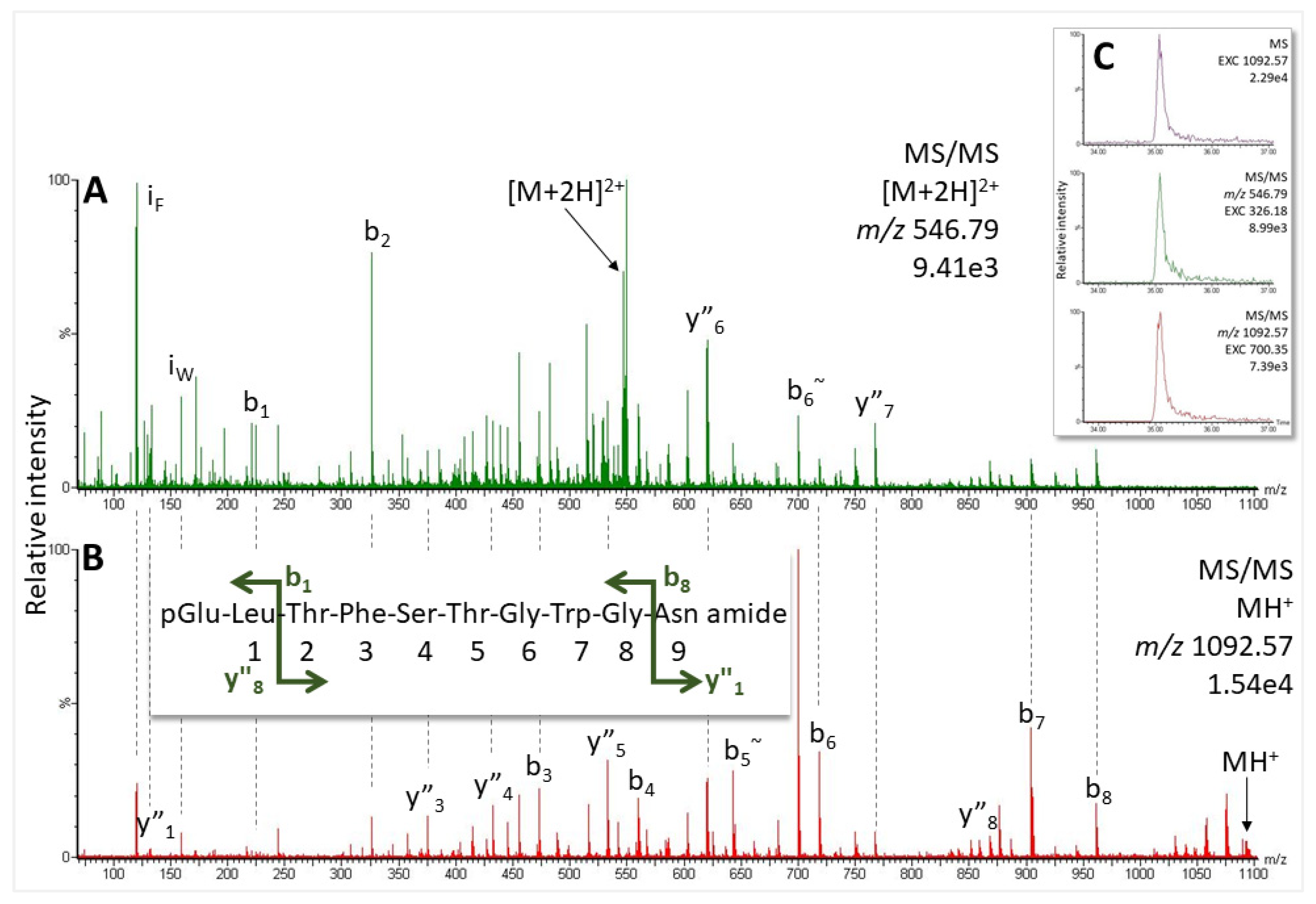
2.3.1. European Corn Borer Ostrinia nubilalis, Family Crambidae
2.3.2. Garden Grass-Veneer Chrysoteuchia culmella, Family Crambidae
3. Discussion
4. Materials and Methods
4.1. Insects and CC Preparation
4.2. Mining of AKH Sequences from Publicly Available Databases
4.3. Synthetic Peptides
4.4. Structure Elucidation by LC-MS
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Nässel, D.R.; Winther, Å. Drosophila neuropeptides in regulation of physiology and behavior. Prog. Neurobiol. 2010, 92, 42–104. [Google Scholar] [CrossRef]
- Gäde, G.; Hoffmann, K.H.; Spring, J.H. Hormonal regulation in insects: Facts, gaps, and future directions. Physiol. Rev. 1997, 77, 963–1032. [Google Scholar] [CrossRef]
- Gäde, G.; Marco, H.G. Structure, function and mode of action of select arthropod neuropeptides. In Bioactive Natural Products (Part M); Atta-Ur-Rahman, Ed.; Elsevier: Amsterdam, The Netherlands, 2006; Volume 33, pp. 69–139. [Google Scholar]
- Marco, H.G.; Gäde, G. Adipokinetic hormone: A hormone for all seasons? In Advances in Invertebrate (Neuro)Endocrinology: A Collection of Reviews in the Post-Genomic Era; Saleuddin, S., Lange, A.B., Orchard, I., Eds.; Apple Academic Press: Palm Bay, FL, USA, 2020; Volume 2, pp. 126–170. [Google Scholar]
- Roch, G.J.; Busby, E.R.; Sherwood, N.M. GnRH receptors and peptides: Skating backward. Gen. Comp. Endocrinol. 2014, 209, 118–134. [Google Scholar] [CrossRef]
- Zandawala, M.; Tian, S.; Elphick, M.R. The evolution and nomenclature of GnRH-type and corazonin-type neuropeptide signalling systems. Gen. Comp. Endocrinol. 2018, 264, 64–77. [Google Scholar] [CrossRef] [PubMed]
- Bradfield, J.Y.; Keeley, L.L. Adipokinetic hormone gene sequence from Manduca sexta. J. Biol. Chem. 1989, 264, 12791–12793. [Google Scholar] [CrossRef]
- O’Shea, M.; Rayne, R.C. Adipokinetic hormones: Cell and molecular biology. Experientia 1992, 48, 430–438. [Google Scholar] [CrossRef]
- Rayne, R.C.; O’Shea, M. Reconstitution of adipokinetic hormone biosynthesis in vitro indicates steps in prohormone processing. Eur. J. Biochem. 1994, 219, 781–789. [Google Scholar] [CrossRef] [PubMed]
- Gäde, G. Peptides of the adipokinetic hormone/red pigment-concentrating hormone family—A new take on biodiversity. In Trends in Comparative Endocrinology and Neurobiology; Vaudry, H., Roubos, E.W., Coast, G.M., Vallarino, M., Eds.; Annals of the New York Academy of Science: Hoboken, NJ, USA; Wiley Blackwell: Hoboken, NJ, USA, 2009; Volume 1163, pp. 125–136. [Google Scholar]
- Gäde, G.; Marco, H.G. The invertebrate AKH/RPCH family: Updated Chapter. In The Handbook of Biologically Active Peptides, 2nd ed.; Kastin, A.J., Ed.; Elsevier: Amsterdam, The Netherlands, 2013; Chapter 28; pp. 185–190. [Google Scholar]
- Gäde, G.; Kellner, R.; Rinehart, K.L.; Proefke, M.L. A tryptophan-substituted member of the AKH/RPCH family isolated from a stick insect corpus cardiacum. Biochem. Biophys. Res. Commun. 1992, 189, 1303–1309. [Google Scholar] [CrossRef]
- Munte, C.E.; Gäde, G.; Domogalla, B.; Kremer, W.; Kellner, R.; Kalbitzer, H.R. C-mannosylation in the hypertrehalosaemic hormone from the stick insect Carausius morosus. FEBS J. 2008, 275, 1163–1173. [Google Scholar] [CrossRef]
- Malik, A.; Gäde, G.; Lange, A.B. Sequencing and biological effects of an adipokinetic/hypertrehalosemic peptide in the stick insect, Baculum extradentatum. Peptides 2012, 34, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Gäde, G.; Šimek, P.; Clark, K.D.; Auerswald, L. Unique translational modification of an invertebrate neuropeptide: A phosphorylated member of the adipokinetic hormone peptide family. Biochem. J. 2006, 393, 705–713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gäde, G.; Šimek, P.; Marco, H.G. An invertebrate [hydroxyproline]-modified neuropeptide: Further evidence for a close evolutionary relationship between insect adipokinetic hormone and mammalian gonadotropin hormone family. Biochem. Biophys. Res. Commun. 2011, 414, 592–597. [Google Scholar] [CrossRef] [PubMed]
- Gäde, G.; Šimek, P.; Marco, H.G. A sulfothreonine adipokinetic peptide—A novel post-translational modification revealed in the twig wilter Holopterna alata (Hemiptera, Coreidae). In Proceedings of the Abstracts of the 27th Conference of European Comparative Endocrinologists, Rennes, France, 25–29 August 2014; p. 68. [Google Scholar]
- König, S.; Bayer, M.; Marco, H.G.; Gäde, G. The hypertrehalosaemic neuropeptide conformational twins of cicadas consist of only L-amino acids: Are they cistrans isomers? Amino. Acids 2019, 51, 1023–1028. [Google Scholar] [CrossRef] [PubMed]
- Gäde, G.; Auerswald, L. Mode of action of neuropeptides from the adipokinetic hormone family. Gen. Comp. Endocrinol. 2003, 132, 10–20. [Google Scholar] [CrossRef]
- Gäde, G. Regulation of intermediary metabolism and water balance of insects by neuropeptides. Annu. Rev. Entomol. 2004, 49, 93–113. [Google Scholar] [CrossRef]
- Adams, M.D.; Celniker, S.E.; Holt, R.A.; Evans, C.A.; Gocayne, J.D.; Amanatides, P.G.; Scherer, S.E.; Li, P.W.; Hoskins, R.A.; Galle, R.F.; et al. The genome sequence of Drosophila melanogaster. Science 2000, 287, 2185–2195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, B.; Predel, R.; Neupert, S.; Hauser, F.; Tanaka, Y.; Cazzamali, G.; Williamson, M.; Arakane, Y.; Verleyen, P.; Schoofs, L.; et al. Genomics, transcriptomics, and peptidomics of neuropeptides and protein hormones in the red flour beetle Tribolium castaneum. Genome Res. 2008, 18, 113–122. [Google Scholar] [CrossRef] [Green Version]
- Hull, J.J.; Gross, R.J.; Brent, C.S.; Christie, A.E. Filling in the gaps: A reevaluation of the Lygus hesperus peptidome using an expanded de novo assembled transcriptome and molecular cloning. Gen. Comp. Endocrinol. 2021, 303, 113708. [Google Scholar] [CrossRef] [PubMed]
- Jaffe, H.; Raina, A.K.; Riley, C.T.; Fraser, B.A.; Nachman, R.J.; Vogel, V.W.; Zhang, Y.-S.; Hayes, D.K. Primary structure of two neuropeptide hormones with adipokinetic and hypotrehalosemic activity isolated from the corpora cardiaca of horse flies (Diptera). Proc. Natl. Acad. Sci. USA 1989, 86, 8161–8164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gäde, G.; Šimek, P.; Marco, H.G. The adipokinetic peptides in Diptera: Structure, function, and evolutionary trends. Front. Endocrin. 2020, 11, 153. [Google Scholar] [CrossRef] [PubMed]
- Marco, H.G.; Šimek, P.; Gäde, G. Unique members of the adipokinetic hormone family in butterflies and moths (Insecta, Lepidoptera). Front. Physiol. 2020, 11, 614552. [Google Scholar] [CrossRef] [PubMed]
- Köllisch, G.V.; Lorenz, M.W.; Kellner, R.; Verhaert, P.D.; Hoffmann, K.H. Structure elucidation and biological activity of an unusual adipokinetic hormone from corpora cardiaca of the butterfly, Vanessa cardui. Eur. J. Biochem. 2000, 267, 5502–5508. [Google Scholar] [CrossRef] [Green Version]
- Marco, H.G.; Gäde, G. Structure and function of adipokinetic hormones of the large white butterfly Pieris brassicae. Physiol. Entomol. 2017, 42, 103–112. [Google Scholar] [CrossRef]
- Xu, G.; Gu, G.-X.; Teng, Z.-W.; Wu, S.-F.; Huang, J.; Song, Q.-S.; Ye, G.-Y.; Fang, Q. Identification and expression profiles of neuropeptides and their G protein-coupled receptors in the rice stem borer Chilo suppressalis. Sci. Rep. 2016, 6, 28976. [Google Scholar] [CrossRef] [Green Version]
- Gäde, G.; Marco, H.G.; Šimek, P.; Marais, E. The newly discovered insect order Mantophasmatodea contains a novel member of the adipokinetic hormone family of peptides. Biochem. Biophys. Res. Commun. 2005, 330, 598–603. [Google Scholar] [CrossRef] [PubMed]
- Predel, R.; Neupert, S.; Huetteroth, W.; Kahnt, J.; Waidelich, D.; Roth, S. Peptidomics-based phylogeny and biogeography of Mantophasmatodea (Hexapoda). Syst. Biol. 2012, 61, 609–629. [Google Scholar] [CrossRef] [PubMed]
- Gäde, G.; Marco, H.G. The adipokinetic peptides of Hemiptera: Structure, function, and evolutionary trends. Front. Insect Sci. 2022, 2, 891615. [Google Scholar] [CrossRef]
- Jiang, S.; Marco, H.G.; Scheich, N.; He, S.; Gäde, G.; McMahon, D.P. Comparative analysis of adipokinetic hormones (AKHs) and AKH receptors (AKHRs) in termites and cockroaches (Blattodea). Insect Biochem. Mol. Biol. 2022. to be submitted. [Google Scholar]
- Wiegmann, B.M.; Trautwein, M.D.; Winkler, I.S.; Barr, N.B.; Kim, J.-W.; Lambkin, C.; Bertone, M.A.; Cassel, B.K.; Bayless, K.M.; Heimberg, A.M.; et al. Episodic radiations in the fly tree of life. Proc. Natl. Acad. Sci. USA 2011, 108, 5690–5695. [Google Scholar] [CrossRef] [Green Version]
- Gäde, G.; Goldsworthy, G.J.; Kegel, G.; Keller, R. Single step purification of locust adipokinetic hormones I and II by reversed-phase high-performance liquid-chromatography, and amino acid composition of the hormone II. Hoppe-Seyler’s Z. Physiol. Chem. 1984, 365, 393–398. [Google Scholar] [CrossRef] [PubMed]
- König, S.; Marco, H.G.; Gäde, G. The proline effect and the tryptophan immonium ion assist in de novo sequencing of adipokinetic hormones. Anal. Chem. 2022. to be submitted. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Insect Species | Primary Sequence | MW | RT (min) | Peptide Name |
|---|---|---|---|---|
| Kalotermes flavicollis | pQ VNFSPG W amide | 915.424 | 32.1 | Manto-CC |
| Pegesimallus tapulus | pQ LTFSPY W amide | 1022.486 | 34.6 | Volpe-CC |
| Pegesimallus tapulus | pQ LTFSPV W amide | 958.491 | 36.8 | Pegta-AKH |
| Haematopota pluvialis | pQ LTFTPG WGY amide | 1150.545 | 35.4 | Tabat-HoTH |
| Haematopota pluvialis | pQ LTFTPG W amide | 930.460 | 34.9 | Tabat-AKH |
| Haematopota pluvialis | pQ LTFTHypG W amide | 946.455 | 33.4 | Haepl-AKH |
| Ostrinia nubilalis | pQ LTFTSS WG amide | 1007.471 | 34.3 | Manse-AKH |
| Ostrinia nubilalis | pQ LTFTSS WGGK | 1193.572 | 32.6 | Vanca-AKH |
| Ostrinia nubilalis | pQ LTFSTG WGQ amide | 1105.519 | 32.8 | Ostnu-AKH |
| Chrysoteuchia culmella | pQ LTFTSS WG amide | 1007.471 | 33.8 | Manse-AKH |
| Chrysoteuchia culmella | pQ LTFTSS WGGK | 1193.572 | 32.1 | Vanca-AKH |
| Chrysoteuchia culmella | pQ LTFSTG WGN amide | 1091.504 | 35.0 | Chipa-AKH |
| Species | Predicted AKH Precursor |
|---|---|
| C. medinalis | MYKYLVILVLVACICVATEAQLTFTSSWGGKRSGAITGGVSCRNDEAIAAIYKMIQ |
| C. culmella | MHKFLALLVIVACVCMAVDAQLTFTSSWGGKRAAIAGTVSCRNDDAIAAIYKMIQ |
| O. nubilalis | MYKFLAVFVLIACMCLVAEAQLTFTSSWGGKRAAIAGTVSCRNDEAIAAIYKMIQVRTPLF |
| O. furnacalis | MYKFLAVFVLIACMCLVAEAQLTFTSSWGGKRAAIAGTVSCRNDEAIAAIYKMIQNEAERFILCQKP |
| C. suppressalis | MYKLLVLLVVVACVCAAVEAQLTFTSSWGGKRAAIAGTVSCRNDEAIAAIYKMIQNEAERFILCQKP |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marco, H.G.; König, S.; Gäde, G. Mass Spectrometric Proof of Predicted Peptides: Novel Adipokinetic Hormones in Insects. Molecules 2022, 27, 6469. https://doi.org/10.3390/molecules27196469
Marco HG, König S, Gäde G. Mass Spectrometric Proof of Predicted Peptides: Novel Adipokinetic Hormones in Insects. Molecules. 2022; 27(19):6469. https://doi.org/10.3390/molecules27196469
Chicago/Turabian StyleMarco, Heather G., Simone König, and Gerd Gäde. 2022. "Mass Spectrometric Proof of Predicted Peptides: Novel Adipokinetic Hormones in Insects" Molecules 27, no. 19: 6469. https://doi.org/10.3390/molecules27196469