

Natural Product 2-Oxokolavenol Is a Novel FXR Agonist

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
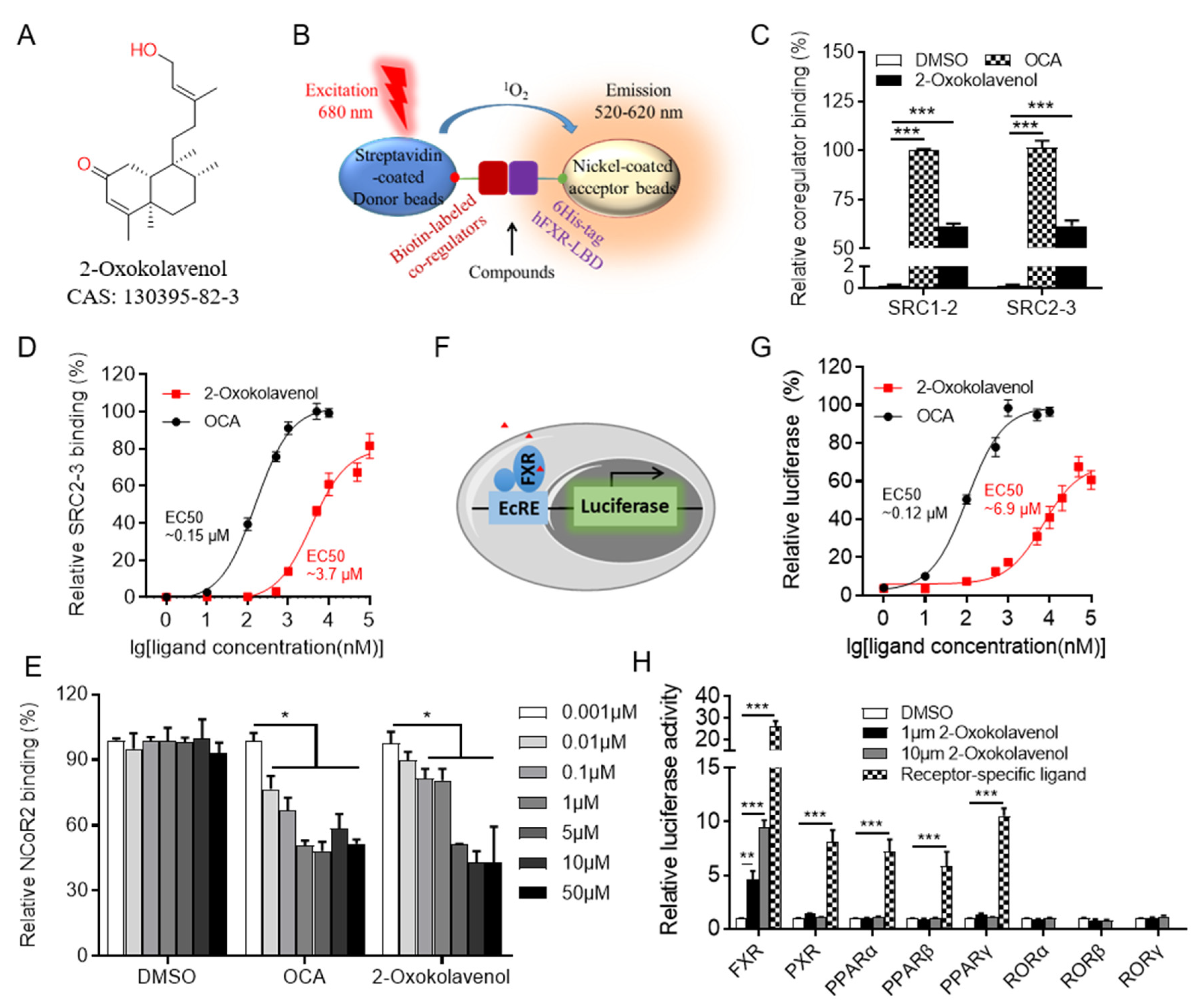
2.1. Identification of the Natural Product 2-Oxokolavenol as a Novel FXR Agonist
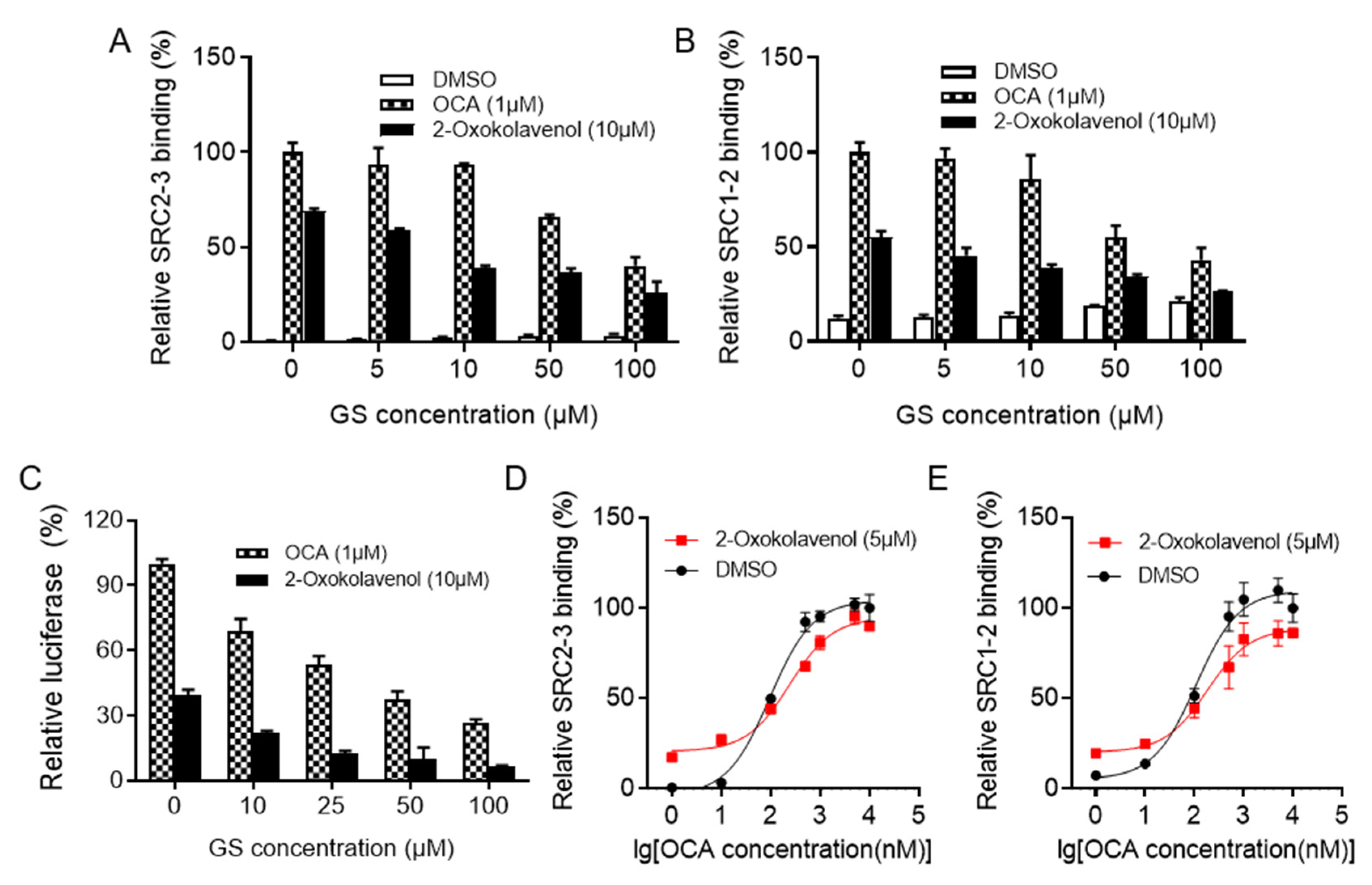
2.2. 2-Oxokolavenol Acts as an Agonist through Direct Interaction with FXR
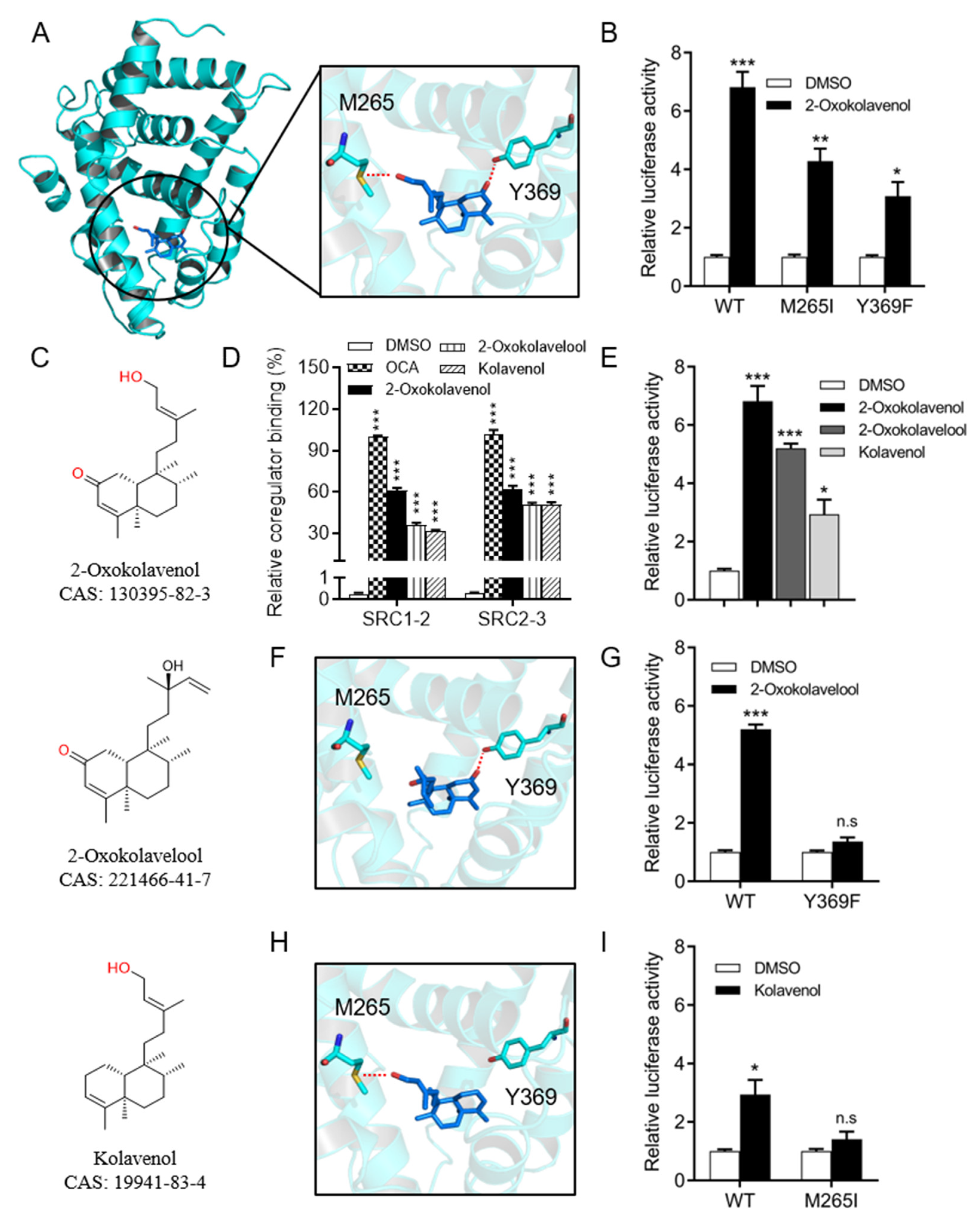
2.3. Molecular Docking of 2-Oxokolavenol/FXR-LBD Complex and Functional Correlation with FXR Interactions
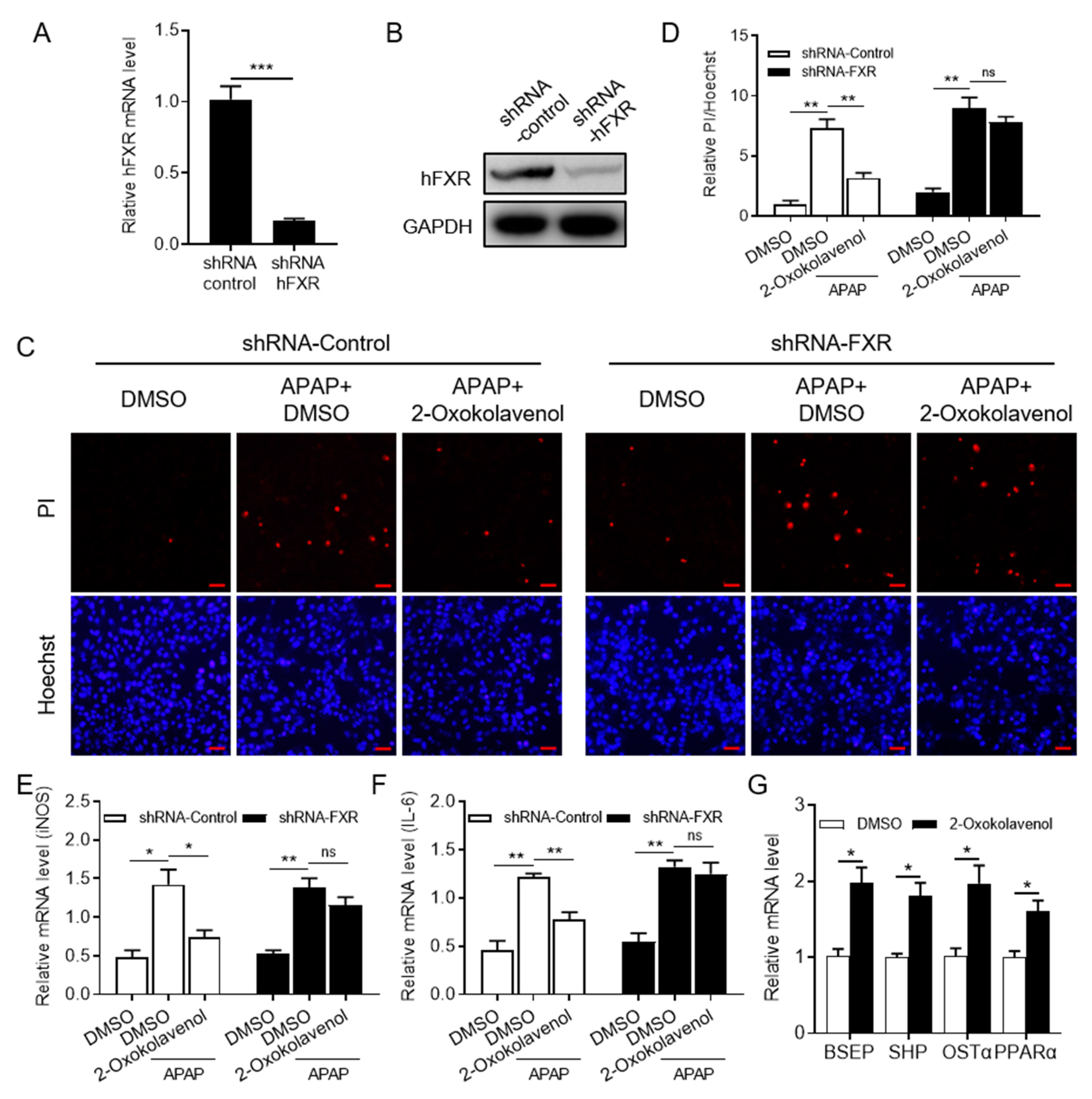
2.4. 2-Oxokolavenol Alleviates APAP-Induced Hepatocytes Damage in an FXR-Dependent Manner in Human Liver WRL68 Cells
3. Discussion
4. Materials and Methods
4.1. Protein Expression and Purification
4.2. AlphaScreen Co-Regulators Binding Assay
4.3. Cell-Based Lluciferase Reporter Assay
4.4. Docking between FXR-LBD and Compounds
4.5. Establishment of WRL68 Cell Line with the Stable Silence of FXR
4.6. APAP-Induced Hepatocytes Damage Cell Model
4.7. RNA Isolation and qPCR
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Goldman, R.D. Acetaminophen in children: An old drug with new warnings. Can. Fam. Physician Med. De Fam. Can. 2013, 59, 1065–1066. [Google Scholar]
- Schilling, A.; Corey, R.; Leonard, M.; Eghtesad, B. Acetaminophen: Old drug, new warnings. Clevel. Clin. J. Med. 2010, 77, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Larson, A.M.; Polson, J.; Fontana, R.J.; Davern, T.J.; Lalani, E.; Hynan, L.S.; Reisch, J.S.; Schiodt, F.V.; Ostapowicz, G.; Shakil, A.O.; et al. Acetaminophen-induced acute liver failure: Results of a United States multicenter, prospective study. Hepatology 2005, 42, 1364–1372. [Google Scholar] [CrossRef] [PubMed]
- Hodgman, M.J.; Garrard, A.R. A review of acetaminophen poisoning. Crit. Care Clin. 2012, 28, 499–516. [Google Scholar] [CrossRef]
- Flatt, B.; Martin, R.; Wang, T.L.; Mahaney, P.; Murphy, B.; Gu, X.H.; Foster, P.; Li, J.; Pircher, P.; Petrowski, M.; et al. Discovery of XL335 (WAY-362450), a highly potent, selective, and orally active agonist of the farnesoid X receptor (FXR). J. Med. Chem. 2009, 52, 904–907. [Google Scholar] [CrossRef]
- Tully, D.C.; Rucker, P.V.; Chianelli, D.; Williams, J.; Vidal, A.; Alper, P.B.; Mutnick, D.; Bursulaya, B.; Schmeits, J.; Wu, X.; et al. Discovery of Tropifexor (LJN452), a Highly Potent Non-bile Acid FXR Agonist for the Treatment of Cholestatic Liver Diseases and Nonalcoholic Steatohepatitis (NASH). J. Med. Chem. 2017, 60, 9960–9973. [Google Scholar] [CrossRef] [PubMed]
- Sepe, V.; Distrutti, E.; Fiorucci, S.; Zampella, A. Farnesoid X receptor modulators 2014–present: A patent review. Expert Opin. Ther. Pat. 2018, 28, 351–364. [Google Scholar] [CrossRef]
- Wang, H.B.; Chen, J.; Hollister, K.; Sowers, L.C.; Forman, B.M. Endogenous bile acids are ligands for the nuclear receptor FXR BAR. Mol. Cell 1999, 3, 543–553. [Google Scholar] [CrossRef]
- Lefebvre, P.; Cariou, B.; Lien, F.; Kuipers, F.; Staels, B. Role of Bile Acids and Bile Acid Receptors in Metabolic Regulation. Physiol. Rev. 2009, 89, 147–191. [Google Scholar] [CrossRef] [Green Version]
- Nevens, F.; Andreone, P.; Mazzella, G.; Strasser, S.I.; Bowlus, C.; Invernizzi, P.; Drenth, J.P.H.; Pockros, P.J.; Regula, J.; Beuers, U.; et al. A Placebo-Controlled Trial of Obeticholic Acid in Primary Biliary Cholangitis. New Engl. J. Med. 2016, 375, 631–643. [Google Scholar] [CrossRef]
- Younossi, Z.M.; Ratziu, V.; Loomba, R.; Rinella, M.; Anstee, Q.M.; Goodman, Z.; Bedossa, P.; Geier, A.; Beckebaum, S.; Newsome, P.N.; et al. Obeticholic acid for the treatment of non-alcoholic steatohepatitis: Interim analysis from a multicentre, randomised, placebo-controlled phase 3 trial. Lancet 2019, 394, 2184–2196. [Google Scholar] [CrossRef] [PubMed]
- Ratziu, V.; Sanyal, A.J.; Loomba, R.; Rinella, M.; Harrison, S.; Anstee, Q.M.; Goodman, Z.; Bedossa, P.; MacConell, L.; Shringarpure, R.; et al. REGENERATE: Design of a pivotal, randomised, phase 3 study evaluating the safety and efficacy of obeticholic acid in patients with fibrosis due to nonalcoholic steatohepatitis. Contemp. Clin. Trials 2019, 84, 105803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, Y.; Wang, H.; Cheng, X.; Wu, Y.; Cao, L.; Wu, M.; Xie, W.; Wang, G.; Hao, H. Farnesoid X receptor activation promotes cell proliferation via PDK4-controlled metabolic reprogramming. Sci. Rep. 2016, 6, 18751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, Y.; Zheng, W.; Lin, S.; Guo, F.; Zhu, Y.; Wei, Y.; Liu, X.; Jin, S.; Jin, L.; Li, Y. Identification of an Oleanane-Type Triterpene Hedragonic Acid as a Novel Farnesoid X Receptor Ligand with Liver Protective Effects and Anti-inflammatory Activity. Mol. Pharmacol. 2018, 93, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Ricketts, M.L.; Boekschoten, M.V.; Kreeft, A.J.; Hooiveld, G.J.E.J.; Moen, C.J.A.; Muller, M.; Frants, R.R.; Kasanmoentalib, S.; Post, S.M.; Princen, H.M.G.; et al. The cholesterol-raising factor from coffee beans, cafestol, as an agonist ligand for the farnesoid and pregnane X receptors. Mol. Endocrinol. 2007, 21, 1603–1616. [Google Scholar] [CrossRef] [Green Version]
- Downes, M.; Verdecia, M.A.; Roecker, A.J.; Hughes, R.; Hogenesch, J.B.; Kast-Woelbern, H.R.; Bowman, M.E.; Ferrer, J.L.; Anisfeld, A.M.; Edwards, P.A.; et al. A chemical, genetic, and structural analysis of the nuclear bile acid receptor FXR. Mol. Cell 2003, 11, 1079–1092. [Google Scholar] [CrossRef]
- Fang, S.; Suh, J.M.; Reilly, S.M.; Yu, E.; Osborn, O.; Lackey, D.; Yoshihara, E.; Perino, A.; Jacinto, S.; Lukasheva, Y.; et al. Intestinal FXR agonism promotes adipose tissue browning and reduces obesity and insulin resistance. Nat. Med. 2015, 21, 159–165. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.W.; Wang, J.; Liu, Q.Y.; Harnish, D.C. Farnesoid X receptor agonist WAY-362450 attenuates liver inflammation and fibrosis in murine model of non-alcoholic steatohepatitis. J. Hepatol. 2009, 51, 380–388. [Google Scholar] [CrossRef]
- Jin, L.H.; Feng, X.H.; Rong, H.; Pan, Z.F.; Inaba, Y.; Qiu, L.; Zheng, W.L.; Lin, S.C.; Wang, R.; Wang, Z.; et al. The antiparasitic drug ivermectin is a novel FXR ligand that regulates metabolism. Nat. Commun. 2013, 4, 1937. [Google Scholar] [CrossRef] [Green Version]
- Jin, L.H.; Wang, R.; Zhu, Y.L.; Zheng, W.L.; Han, Y.P.; Guo, F.S.; Bin Ye, F.; Li, Y. Selective targeting of nuclear receptor FXR by avermectin analogues with therapeutic effects on nonalcoholic fatty liver disease. Sci. Rep. 2015, 5, 17288. [Google Scholar] [CrossRef] [Green Version]
- Ratziu, V.; Rinella, M.E.; Neuschwander-Tetri, B.A.; Lawitz, E.; Denham, D.; Kayali, Z.; Sheikh, A.; Kowdley, K.V.; Desta, T.; Elkhashab, M.; et al. EDP-305 in patients with NASH: A phase II double-blind placebo-controlled dose-ranging study. J. Hepatol. 2022, 76, 506–517. [Google Scholar] [CrossRef] [PubMed]
- Harrison, S.A.; Bashir, M.; Shim-Lopez, J.; Song, K.; Chen, H. Met409, a Sustained Fxr Agonist, Decreased Hepatic Fat and Improved Liver Function without Raising Ldl-C after 28 Days in Nash Patients. Hepatology 2019, 70, 1257a. [Google Scholar]
- Genin, M.J.; Bueno, A.B.; Francisco, J.A.; Manninen, P.R.; Bocchinfuso, W.P.; Montrose-Rafizadeh, C.; Cannady, E.A.; Jones, T.M.; Stille, J.R.; Raddad, E.; et al. Discovery of 6-(4-[1]piperidin-1-yl)-1-methyl-1H-indole-3-carboxylic Acid: A Novel FXR Agonist for the Treatment of Dyslipidemia. J. Med. Chem. 2015, 58, 9768–9772. [Google Scholar] [CrossRef] [PubMed]
- Kowdley, K.V.; Minuk, G.Y.; Pagadala, M.R.; Gulamhusein, A.; Swain, M.G.; Neff, G.W.; Zogg, D.; Bowlus, C.L.; Agarwal, K.; Yoshida, E.M.; et al. The Nonsteroidal Farnesoid X Receptor (Fxr) Agonist Cilofexor Improves Liver Biochemistry in Patients with Primary Biliary Cholangitis (Pbc): A Phase 2, Randomized, Placebo-Controlled Trial. Hepatology 2019, 70, 31a–32a. [Google Scholar]
- Loomba, R.; Noureddin, M.; Kowdley, K.V.; Kohli, A.; Sheikh, A.; Neff, G.; Bhandari, B.R.; Gunn, N.; Caldwell, S.H.; Goodman, Z.; et al. Combination Therapies Including Cilofexor and Firsocostat for Bridging Fibrosis and Cirrhosis Attributable to NASH. Hepatology 2021, 73, 625–643. [Google Scholar] [CrossRef]
- Badman, M.K.; Chen, J.; Desai, S.; Vaidya, S.; Neelakantham, S.; Zhang, J.; Gan, L.; Danis, K.; Laffitte, B.; Klickstein, L.B. Safety, Tolerability, Pharmacokinetics, and Pharmacodynamics of the Novel Non-Bile Acid FXR Agonist Tropifexor (LJN452) in Healthy Volunteers. Clin. Pharmacol. Drug Dev. 2020, 9, 395–410. [Google Scholar] [CrossRef]
- Jiang, L.Y.; Xiao, D.S.; Li, Y.B.; Dai, S.Y.; Qu, L.Z.; Chen, X.J.; Guo, M.; Wei, H.D.; Chen, Y.H. Structural basis of tropifexor as a potent and selective agonist of farnesoid X receptor. Biochem. Biophys. Res. Commun. 2021, 534, 1047–1052. [Google Scholar] [CrossRef]
- Mudaliar, S.; Henry, R.R.; Sanyal, A.J.; Morrow, L.; Marschall, H.U.; Kipnes, M.; Adorini, L.; Sciacca, C.I.; Clopton, P.; Castelloe, E.; et al. Efficacy and safety of the farnesoid X receptor agonist obeticholic acid in patients with type 2 diabetes and nonalcoholic fatty liver disease. Gastroenterology 2013, 145, 574–582.e1. [Google Scholar] [CrossRef]
- Neuschwander-Tetri, B.A.; Loomba, R.; Sanyal, A.J.; Lavine, J.E.; Van Natta, M.L.; Abdelmalek, M.F.; Chalasani, N.; Dasarathy, S.; Diehl, A.M.; Hameed, B.; et al. Farnesoid X nuclear receptor ligand obeticholic acid for non-cirrhotic, non-alcoholic steatohepatitis (FLINT): A multicentre, randomised, placebo-controlled trial. Lancet 2015, 385, 956–965. [Google Scholar] [CrossRef] [Green Version]
- Atanasov, A.G.; Zotchev, S.B.; Dirsch, V.M.; Supuran, C.T.; Taskforce, I.N.P.S. Natural products in drug discovery: Advances and opportunities. Nat. Rev. Drug Discov. 2021, 20, 200–216. [Google Scholar] [CrossRef]
- Chen, K.Q.; Wu, F.; Lei, X.G. Function-Oriented Natural Product Synthesis. Chin. J Chem 2021, 39, 838–854. [Google Scholar] [CrossRef]
- Yang, S.M.; Wu, S.H.; Qin, X.D.; Luo, X.D.; Wu, D.G. Neoclerodane diterpenes from Amoora stellato-squamosa. Helv. Chim. Acta 2004, 87, 1279–1286. [Google Scholar] [CrossRef]
- Zhu, Y.L.; Xu, S.S.; Lu, Y.; Wei, Y.J.; Yao, B.Q.; Guo, F.S.; Zheng, X.; Wang, Y.M.; He, Y.; Jin, L.H.; et al. Repositioning an Immunomodulatory Drug Vidofludimus as a Farnesoid X Receptor Modulator with Therapeutic Effects on NAFLD. Front. Pharmacol. 2020, 11, 590. [Google Scholar] [CrossRef] [PubMed]
- Urizar, N.L.; Liverman, A.B.; Dodds, D.T.; Silva, F.V.; Ordentlich, P.; Yan, Y.Z.; Gonzalez, F.J.; Heyman, R.A.; Mangelsdorf, D.J.; Moore, D.D. A natural product that lowers cholesterol as an antagonist ligand for FXR. Science 2002, 296, 1703–1706. [Google Scholar] [CrossRef]
- Koehn, F.E.; Carter, G.T. The evolving role of natural products in drug discovery. Nat. Rev. Drug Discov. 2005, 4, 206–220. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, F.; Gao, Y.; Li, X.; Lei, X. Natural Product 2-Oxokolavenol Is a Novel FXR Agonist. Molecules 2022, 27, 8968. https://doi.org/10.3390/molecules27248968
Guo F, Gao Y, Li X, Lei X. Natural Product 2-Oxokolavenol Is a Novel FXR Agonist. Molecules. 2022; 27(24):8968. https://doi.org/10.3390/molecules27248968
Chicago/Turabian StyleGuo, Fusheng, Yihui Gao, Xiaobao Li, and Xiaoguang Lei. 2022. "Natural Product 2-Oxokolavenol Is a Novel FXR Agonist" Molecules 27, no. 24: 8968. https://doi.org/10.3390/molecules27248968
APA StyleGuo, F., Gao, Y., Li, X., & Lei, X. (2022). Natural Product 2-Oxokolavenol Is a Novel FXR Agonist. Molecules, 27(24), 8968. https://doi.org/10.3390/molecules27248968