1. Introduction
Scutellaria lateriflora, commonly known as American skullcap, is one of the most widely utilized nerve tonics in Western herbal medicine. For over 200 years,
S. lateriflora has been used for the treatment of anxiety and the promotion of a healthy menstrual cycle. Other traditional uses of skullcap include hysteria, delirium tremens, withdrawal from barbiturates and tranquilizers, seizures, bronchitis, dysentery, diarrhea, jaundice, hepatitis, tumors, hypertension, and thrombosis [
1]. Chemically, the aerial parts of this plant contain various classes of polyphenols, essential oils, diterpenoids, and amino acids. The polyphenolic profile of the hydroalcoholic extract used in this study, obtained from the aerial parts of the plant
S. lateriflora, was previously determined using ultra-high-performance liquid chromatography coupled with a Q-Exactive hybrid quadrupole orbitrap mass spectrometer. This analysis allowed for the identification of over one hundred compounds, including flavonoids (baicalein, baicalin, dihydrobaicalin, oroxylin A, wogonin, lateriflorein, lateriflorin, scutellarin, ikonnikoside I, oroxylin A-7-
O-glucuronide, and 2′-methoxy-chrysin-7-
O-glucuronide), and phenolic acids (cinnamic acid, caffeic acid, ferulic acid, and
p-coumaric acid) [
2,
3].
The Italian Health Ministry guidelines on the traditional effects of vegetable extracts used in food supplements report that
S. lateriflora shows relaxation activity, but the scientific literature only reports studies on the antistress effects of another species of the genus Scutellaria,
Scutellaria baicalensis, while studies on
S. lateriflora are absent. Udintsev et al. [
4] demonstrated a normalization of all hormonal metabolic alterations, including levels of insulin, glucose, urea, corticotropin, and hydroxycorticosteroids, in rats exposed to stress by fixation. Ryu et al. [
5] and Lee et al. [
6] reported the antistress effects of
S. baicalensis extract in rodent experimental models, where the extract showed a considerable decrease in the blood levels of corticosterone.
Cortisol, a stress hormone, is released by the hypothalamic-pituitary-adrenal (HPA) axis and acutely and chronically affects the metabolic, cardiovascular, and nervous systems. Thus, a balanced regulation of the release of stress-induced cortisol is essential to maintaining homeostasis and a healthy status [
7]. The amygdala sends signals to the hypothalamus in response to stress, and this activates the sympathetic nervous system, resulting in the release of catecholamines and cortisol by the adrenal glands. Catecholamines, such as adrenaline, increase heart and respiratory rates, while cortisol provides energy to the body and allows the body to stay on high alert in response to stress [
8,
9]. Cortisol release is regulated by a negative feedback mechanism in the central nervous system, where basal nonstressed cortisol secretion follows the circadian rhythm, with levels in the body rising sharply upon awakening and declining over the course of the day [
10].
Dysregulated and excessive secretion of cortisol is associated with psychiatric issues, mainly insomnia, anxiety, and depression. Literature data have shown a significant relationship between sleep deprivation and HPA axis hyperactivation, resulting in neuroendocrine dysregulation [
11]. One study reports a positive correlation between polysomnographic indices of insomnia in adults and urinary levels of free cortisol [
12]. Insomnia patients usually present with high cortisol levels, mainly in the evening and at the onset of sleep, suggesting that hypercortisolism could be a marker of corticotropin-releasing hormone and noradrenaline activity during the night [
13]. Importantly, HPA axis-induced sleep fragmentation and the associated increase in cortisol levels suggest that the HPA axis contributes not only to the establishment, but also to the continuation of chronic insomnia [
14]. Moreover, approximately 50% of newly diagnosed patients with depression present elevated levels of cortisol [
15]. In addition, increased levels of cortisol in the body may also contribute to osteopenia and osteoporosis by decreasing bone mineral density. It alters bone turnover, impairs intestinal absorption and renal reabsorption of calcium, and inhibits reproductive hormones in postmenopausal women [
16].
There are a number of drugs that have long been proven to alter cortisol synthesis and/or release, including serotonin antagonists, dopaminergic agonists, metyrapone, ketoconazole, fluconazole, reserpine, valproic acid, and somatostatin analogs. However, these drugs present questionable efficacy and tolerability and are only recommended for severe forms of hypercortisolism, such as Cushing syndrome [
17,
18]. Patients suffering from insomnia, anxiety, and mild-to-moderate depression, which are not eligible for the available anticortisol therapies, are usually treated with conventional drugs targeting the symptoms of the disease. Thus, there is a strong need for the development of new alternative agents with cortisol inhibitory properties and favorable safety profiles, which can then be used in the treatment of mild-to-moderate hypercortisolism, such as sleep disorders, stress, and anxiety.
The present study is aimed at an in vitro evaluation of cortisol inhibition activities as a possible mechanism of action of a chemically characterized S. lateriflora extract. In view of its use as a food supplement ingredient, the in vitro bioaccessibility and bioavailability of the extract were also studied.
3. Discussion
In this investigation, a commercial extract rich in polyphenols obtained from the aerial parts of
S. lateriflora, previously characterized using ultra-high-performance liquid chromatography (UHPLC) coupled with a Q-Exactive hybrid quadrupole orbitrap mass spectrometer [
3] was submitted to in vitro studies, including the determination of bioaccessibility, bioavailability, and investigation of cortisol release inhibition as a possible mechanism of action through which
S. lateriflora exerts its protective effects against the numerous health concerns including psychiatric issues (i.e., stress, insomnia, anxiety, and depression) and nonpsychiatric issues (i.e., osteopenia and osteoporosis) associated to an excess in cortisol release.
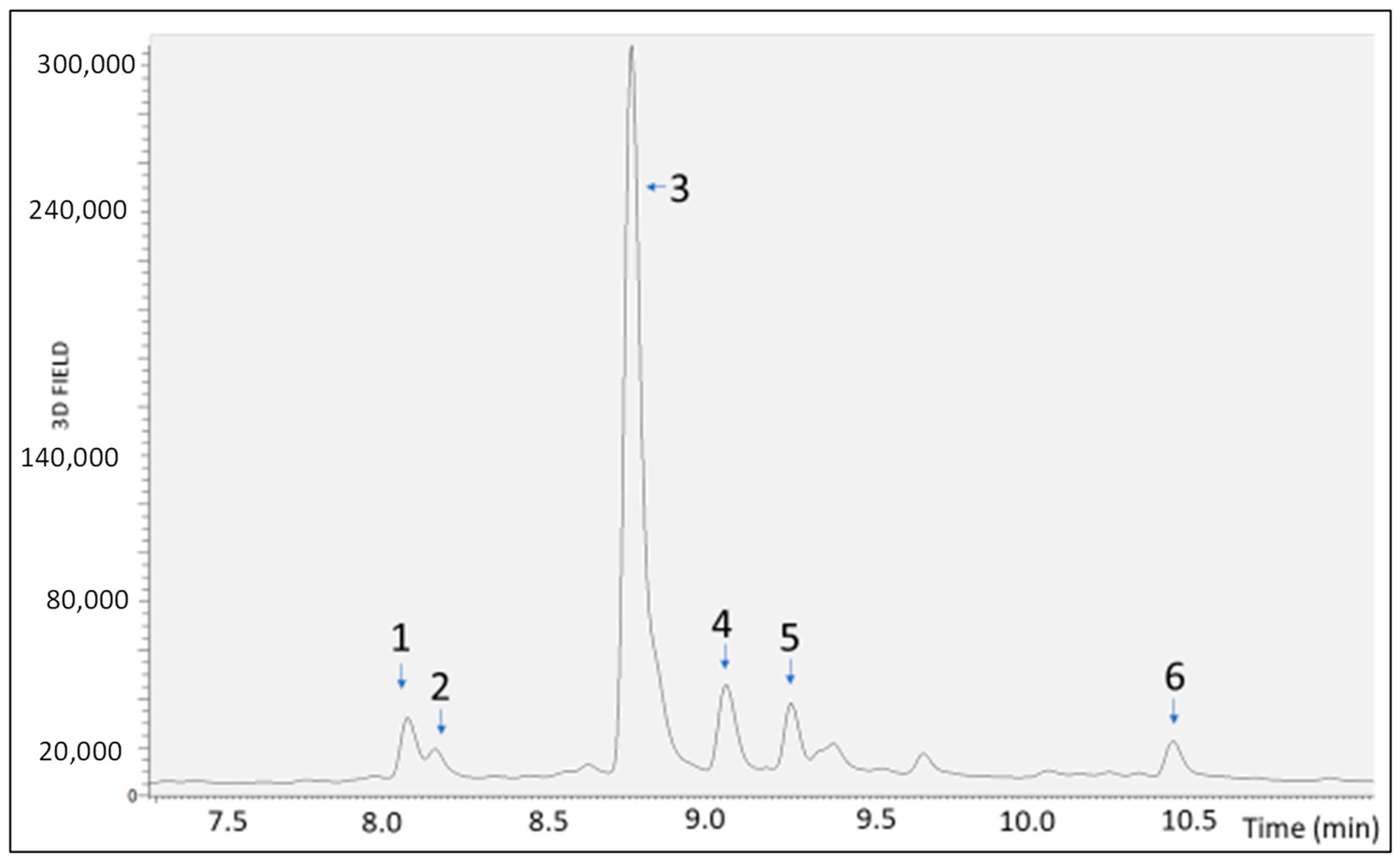
In vitro simulated gastric, duodenal, and gastroduodenal digestion processes revealed a significant reduction in the peak area of some of the most represented polyphenolic compounds, including flavones (apigenin derivative, scutellarin, baicalein 6-glucuronide, and oroxilyn A glucuronide), a flavanone (naringenin 7-O-d-hexoside 6″ acetate), and an isoflavone (genistein). Postdigestion, greater degradation was observed for scutellarin and genistein, while the degradation was less pronounced for the other compounds.
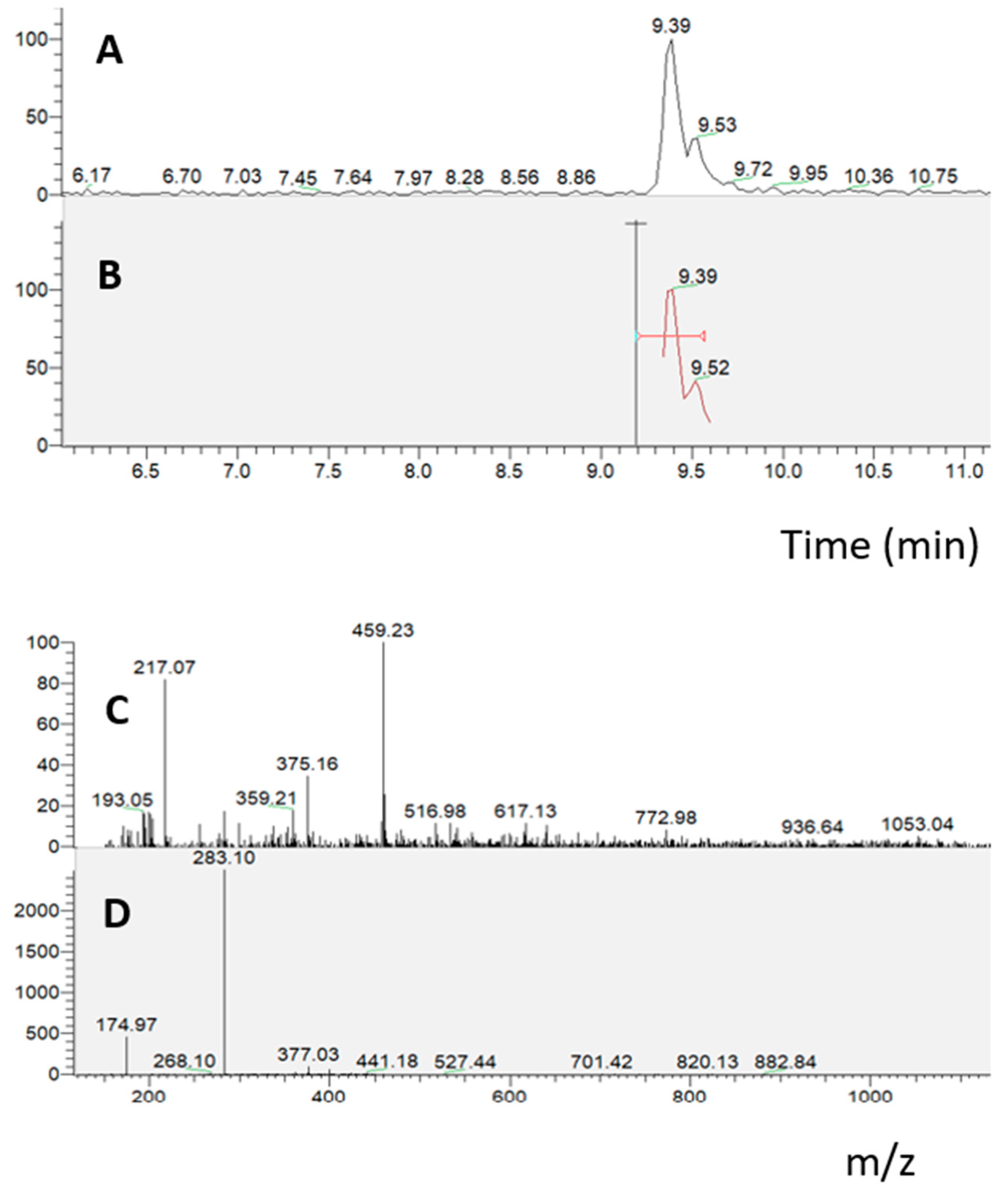
The bioavailability assays (Caco-2 cells grown on transwell insert assay and PAMPA assay) showed oroxylin A glucuronide and oroxylin A as the compounds of interest, as they were the only compounds that were able to cross the Caco-2 cell membrane and the artificial lipid membrane, respectively. It is interesting that, in agreement with hydrophilic and lipophilic features, oroxylin A, which is more liposoluble than its glucuronide form, crosses the artificial lipid membrane, while the glucuronide form crosses the cell monolayer.
Extensive literature data indicates a considerable reduction in the polyphenolic content of vegetable extracts [
19,
20,
21] after gastroduodenal digestion. None of the previous studies reported the bioaccessibility of
S. lateriflora extracts, and this is the first study to report the bioaccessibility of the main polyphenolics of
S. lateriflora extract following gastroduodenal digestion. A study by Ullah et al. [
3] on the in vitro bioaccessibility of a
S. lateriflora extract after oral digestion showed a modest reduction in the concentrations of the main components of the extract, with a greater reduction in the chromatographic peak area of scutellarin (8.9%). While assessing the in vitro bioaccessibility of
Eugenia pyriformis Cambess. fruit, De Paulo Farias et al. [
22] observed a significant reduction in the total flavonoid contents (including apigenin hexoside) following gastric (48%) and intestinal (70%) digestion. Conversely, our results appear to contradict those reported by Walsh et al. [
23], which demonstrated the good bioaccessibility of isoflavones due to their higher stability under gastroduodenal digestion conditions, although the authors concluded that this good stability of phenolic compounds may be attributed to an increased secretion of bile acid stimulated by the presence of proteins and fats in food samples. Baicalein has been reported as having poor water solubility (16.82 μg/mL) that reduces its bioaccessibility and, in turn, limits the bioavailability of the compound [
24,
25]. It has also been documented that polyphenols are highly sensitive to the mild alkaline conditions in the small intestine, where degradation of most polyphenolic compounds takes place or, in other cases, transformation into other metabolites occurs [
26].
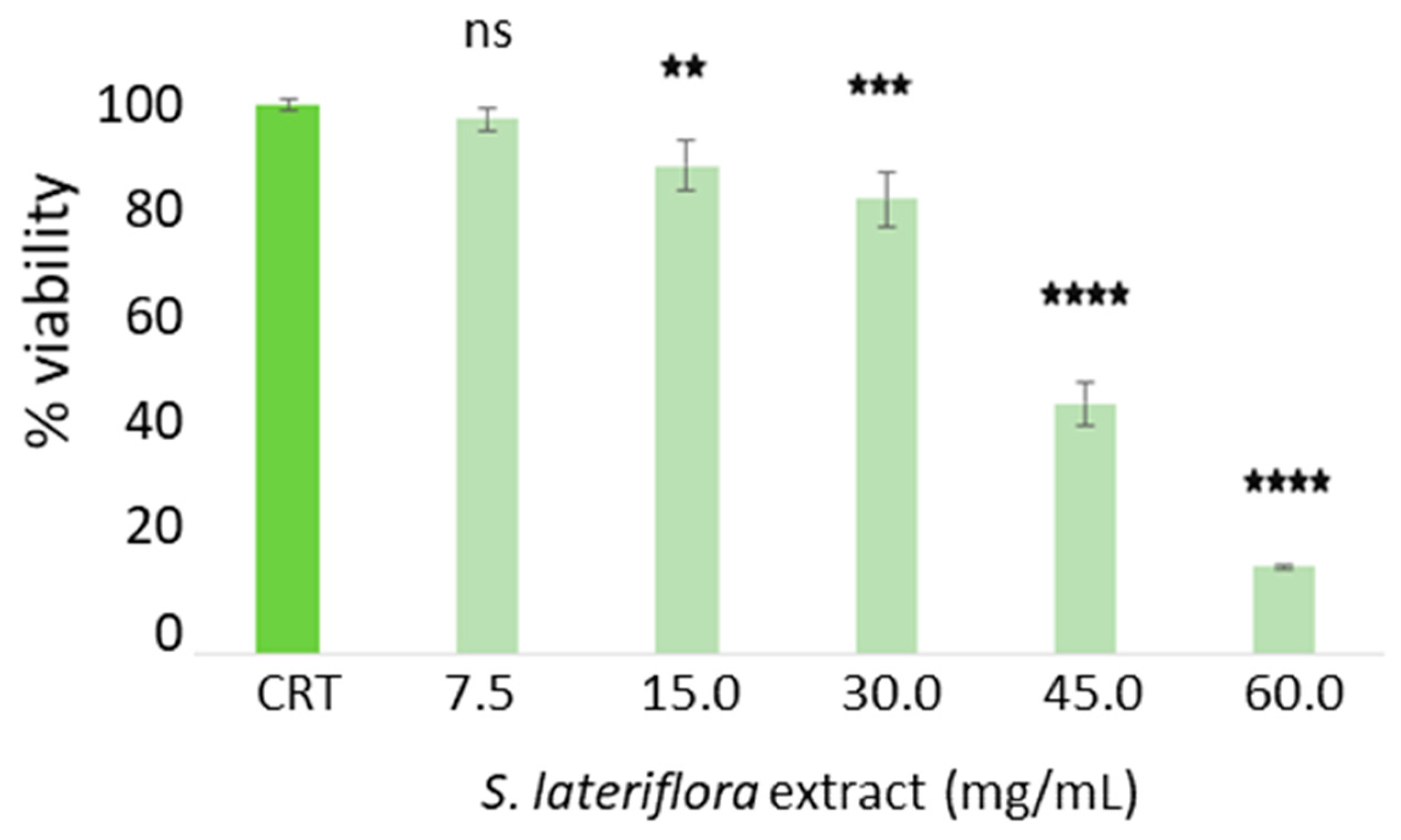
Prior to the assessment of the bioavailability of
S. lateriflora extract components, the noncytotoxic concentration of
S. lateriflora extract, corresponding to a cell viability higher than 70%, was evaluated by MTT assay and was found to be 30 mg/mL. Our results, showing the low bioavailability of the polyphenolic compounds occurring in
S. lateriflora extract, are in agreement with the available literature, where low bioavailability of flavonoids (i.e., apigenin, scutellarein, genistein, baicalein, quercetin, and naringenin) is reported across the studies [
26,
27,
28,
29,
30,
31,
32]. The main properties limiting the bioavailability of polyphenols are their poor water solubility and low potential to penetrate the intestinal membrane, as reported in the case of scutellarin [
33,
34], which has a water solubility of only approximately 14–20 µg/mL and does not cross the intestinal membrane. Another factor that might contribute to the low bioavailability of polyphenols is their strong association with cell walls, which may result in their retention in the nondigested fraction [
35]. Moreover, in the present study,
S. lateriflora extract was subjected only to simulate in vitro duodenal digestion before the bioavailability assays, without fermentation with gut microbiota. This could be a limitation, as most flavonoids generally require metabolic transformation through the intestinal microflora prior to absorption through the intestinal cell membrane [
36]. Although it is difficult to compare data obtained from in vitro tests with data obtained in vivo on experimental animals, however the in vivo pharmacokinetics analysis of Scutellariae radix, with the determination of baicalin, wogonoside, and oroxylin, indicated the presence of all these flavonoids in blood samples in their glycoside and aglycone forms after oral administration of rats with this plant extract [
37]. Partially in agreement with our study, oroxylin A glucuronide was the glycoside present in the highest concentration, while baicalein was the flavonoid with the poorest intestinal permeability as both glucuronide and aglycone. Another study showed the rapid absorption, tissue distribution, and elimination of oroxylin A aglycone and glucuronide after oral administration [
38].
Oroxylin A is a flavone, containing one methoxy group at carbon 6 and two hydroxyl (OH) groups at carbons 5 and 7. Oroxylin A 7-
O-β-
d-glucuronide is one of the principle metabolites of oroxylin A, and is a monomethoxy and dihydroxy flavone derivative of the parent compound [
39]. Based on its chemistry, oroxylin A could be a flavonoid with strong bioactivity. Indeed, some studies have revealed a wide spectrum of biological activities of oroxylin A, such as anti-inflammatory, anticancer, anti-invasive, neuroprotective, and antiangiogenic effects [
40]. However, the cortisol inhibition effects of oroxylin A have not been reported to date, but the potential neurological effects (i.e., dopamine reuptake inhibition, regulation of brain-derived neurotrophic factor (BDNF) expression, improved memory and cognitive functions, neuroprotective actions via decreasing markers for neuroinflammation, improvement of the attention-deficit hyperactivity disorder (ADHD)-like behaviors) do indicate its activity within the nervous system [
40]. A study showed beneficial effects against chronic stress through the regulation of hippocampal BDNF expression and neurogenesis in C57BL/6J mice [
41].
Prior to the assessment of its in vitro inhibition of cortisol release, the noncytotoxic concentration of
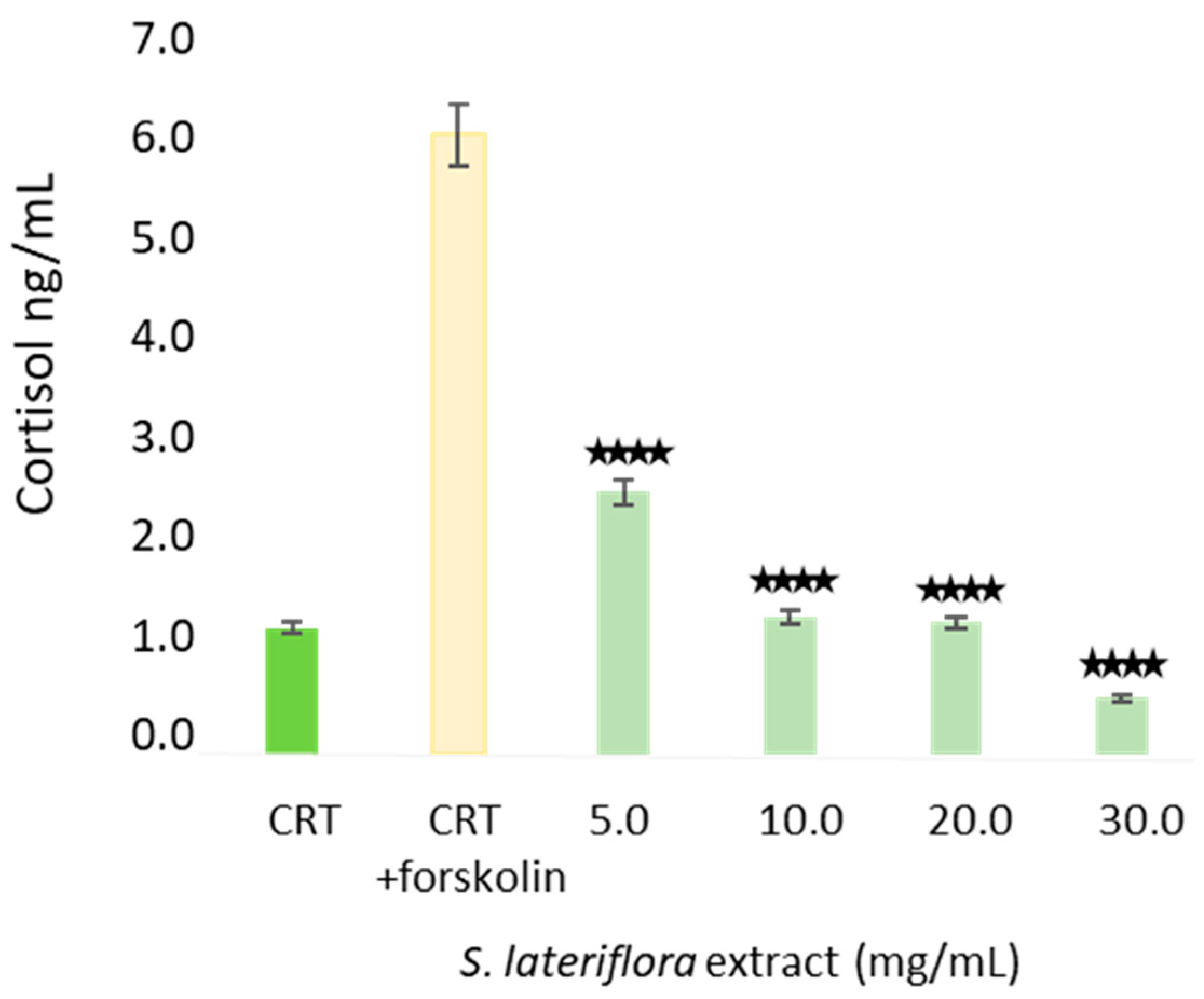
S. lateriflora extract, corresponding to a cell viability higher than 70%, was evaluated by MTT assay and was found to be 30 mg/mL. The treatment of H295R cells with noncytotoxic concentrations of
S. lateriflora extract showed a significant inhibition of cortisol release for all tested doses in a dose-dependent manner, with 30 mg/mL being the most potent concentration, resulting in 91% of cortisol inhibition. Increasing evidence suggests the potential link between circulating cortisol levels and stress-related disorders, including depression and anxiety, as stress disorders are linked with the activity of the HPA axis. In early investigations, patients with coexisting panic disorders, depression, and agoraphobia were found to have significantly elevated urinary cortisol levels compared to healthy individuals, and benzodiazepines were found to be effective in decreasing these free cortisol levels [
42]. Another study showed elevated nocturnal levels of stress hormones, including cortisol, epinephrine, and norepinephrine, in subjects with panic disorders [
43]. An investigation by Vedhara et al. [
44] exhibited a nonlinear relationship between cortisol levels and the time of day, where this nonlinear phenomenon was largely based on stress and anxiety levels. The study determined a change in cortisol levels at specific times of day, but changes in absolute levels of the hormone were not observed. Vreeburg et al. [
45] demonstrated significantly higher salivary levels of cortisol early in the morning (i.e., 1 h cortisol awakening response) in patients with panic disorders comorbid with agoraphobia and depression.
This work has its limitations and strengths. The main limitation of this study is the exclusion of fermentation by the gut microbiota, due to which the potential impact of the gut microbiome on the bioaccessibility and bioavailability of polyphenolic compounds remains undetermined. Another limitation of the present study is that it does not explore the inhibition of cortisol release at the molecular level. Hence, this study could be considered a preliminary evaluation, contributing scientific evidence to the theory that the relaxation properties of the S. lateriflora extract may be justified by inhibition of cortisol release. The major strength of this study is that the property of S. lateriflora extracts to the inhibit cortisol release is determined for the first time, suggesting a possible mechanism of action behind the relaxation activity traditionally ascribed to S. lateriflora. Another strength of this study is that the bioaccessibility and bioavailability of S. lateriflora extract were investigated for the first time, providing a picture of the bioavailable compounds and suggesting the need for gastroprotective dosage forms.
In conclusion, the S. lateriflora extract could be a good agent against the cortisol-mediated stress response and associated pathologies. This effect could also be useful for counteracting bone loss and osteoporosis induced by high cortisol levels, since, as previously mentioned, cortisol levels and bone density are frequently linked, especially in high-risk and postmenopausal women. However, the low bioaccessibility of the S. lateriflora extract suggests the use of a gastroprotectant formulation for the possible applications of this extract as a food supplement ingredient for patients complaining of insomnia and sleep disorders. A clinical trial assessing the efficacy and tolerability of a S. lateriflora extract-based food supplement for maintaining a correct sleep-wake cycle is currently in progress.
4. Materials and Methods
4.1. Chemicals, Reagents and Biological Materials
Caco-2 and H295R cells were purchased from the American Type Culture Collection (ATCC CRL-2128™; distribution by LGC group, London, UK). The cell culture medium, fetal bovine serum (FBS), nonessential amino acids and lucifer yellow (LY) were bought from M & M Biotech (Naples, Italy). All other cell culture reagents were purchased from Sigma-Aldrich (St. Louis, MO, USA). Finally, transwell PET inserts were obtained from Sterlitech Corporation (Washington, DC, USA). Human COR ELISA Kit (3455-HP-2) was bought from FineTest® (Prodotti Gianni S.r.l., Milan, Italy).
Three batches of commercial, dry, powdered hydroalcoholic extract of S. lateriflora (standardized to contain ≥10% of baicalin, and maize maltodextrin as a carrier agent) obtained from the aerial parts of the plants, were provided by EPO S.R.L. (Milan, Italy). All the compounds used for in vitro gastric and duodenal digestion processes have been reported as follows: potassium chloride (KCl), dihydrogen potassium phosphate (KH2PO4), sodium carbonate (NaHCO3), magnesium chloride (MgCl2), ammonium carbonate (NH4)2CO3, calcium chloride (CaCl2), sodium chloride (NaCl), hydrochloric acid (HCl), sodium hydroxide (NaOH). All compounds were provided by Carlo Erba (Milan, Italy). Pancreatin from a porcine pancreas (extract of pig bile), pepsin from porcine gastric mucosa and porcine bile extract, formic acid solution (1 M), water, methanol, acetonitrile LC-MS grade, and dimethyl sulfoxide (DMSO) were purchased from Sigma-Aldrich, Merck KGaA (Milan, Italy).
4.2. In Vitro Bioaccessibility of S. lateriflora Extract
In vitro digestion of
S. lateriflora extract was performed by simulated gastric, duodenal, and gastroduodenal digestion processes following protocols set by Minekus et al. with slight modifications [
46].
4.2.1. In Vitro Simulated Gastric Digestion
To evaluate the effect of the gastric digestion process on S. lateriflora extract, 1.0 g of sample was dissolved in 2 mL of bidistilled water. Simulated gastric fluid (SGF) electrolyte stock solution (1.5 mL) was then added, followed by 0.32 mL porcine pepsin stock solution (25,000 U/mL prepared in SGF) and 1 μL of CaCl2 (0.3 M). Finally, 0.2 mL of HCl (1 M) was added to achieve pH 3.0 and made up with bidistilled water to obtain a final volume of 4 mL. The process was repeated, replacing the sample with a blank consisting of 2 mL of bidistilled water. The reaction vessels were placed onto a shaking platform at 37 °C for 2 h. At the end of the digestion process, the samples were kept at −20 °C prior to RP-UHPLC-PDA-ESI-MS/MS analysis.
4.2.2. In Vitro Simulated Duodenal Digestion
Similarly, as described before, 1.0 g of sample was dissolved in 4 mL of bidistilled water, mixed with 2.2 mL of simulated intestinal fluid (SIF) electrolyte stock solution, 1 mL of a pancreatin solution (800 U/mL prepared in SIF electrolyte stock solution), 0.5 mL fresh bile (160 mM in fresh bile) and 8 μL of CaCl2 (0.3 M). Once the reaction tube was prepared, 0.15 mL of NaOH (1 M) was added to reach pH 7.0 with bidistilled water to obtain a final volume of 8 mL. Here too, the process was repeated in order to have a negative control. Finally, the reaction vessels were placed onto a shaking platform at 37 °C for 2 h. At the end of the digestion process, the samples were maintained at −20 °C prior to RP-UHPLC-PDA-ESI-MS/MS analysis.
4.2.3. In Vitro Simulated Gastroduodenal Digestion
To assess the effect of the full gastroduodenal process on S. lateriflora extract, 1.0 g of sample was dissolved in 2 mL of bidistilled water, and 0.3 mL of SGF electrolyte stock solution was then added, followed by 0.064 mL porcine pepsin stock solution (25,000 U/mL prepared in SGF) and 1 μL of CaCl2 (0.3 M). Subsequently, 0.2 mL of HCl (1 M) was added to reach a solution of pH 3.0 with bidistilled water to obtain a total volume of 4 mL. A 4 mL blank sample of bidistilled water was put through the same process. The reaction vessels were then placed onto a shaking platform at 37 °C for 2 h. Then, the simulated gastric samples were added to 2.2 mL of SIF electrolyte stock solution, 1 mL of a pancreatin solution (800 U/mL prepared in SIF electrolyte stock solution), 0.5 mL fresh bile (160 mM in fresh bile), and 8 μL of CaCl2 (0.3 M). Moreover, 0.15 mL of NaOH (1 M) were added to reach pH 7.0, with bidistilled water to reach the final volume of 8 mL. Finally, the reaction vessels were placed onto a shaking platform at 37 °C for 2 h. At the end of the digestion process, the samples were maintained at −20 °C prior to RP-UHPLC-PDA-ESI-MS/MS analysis.
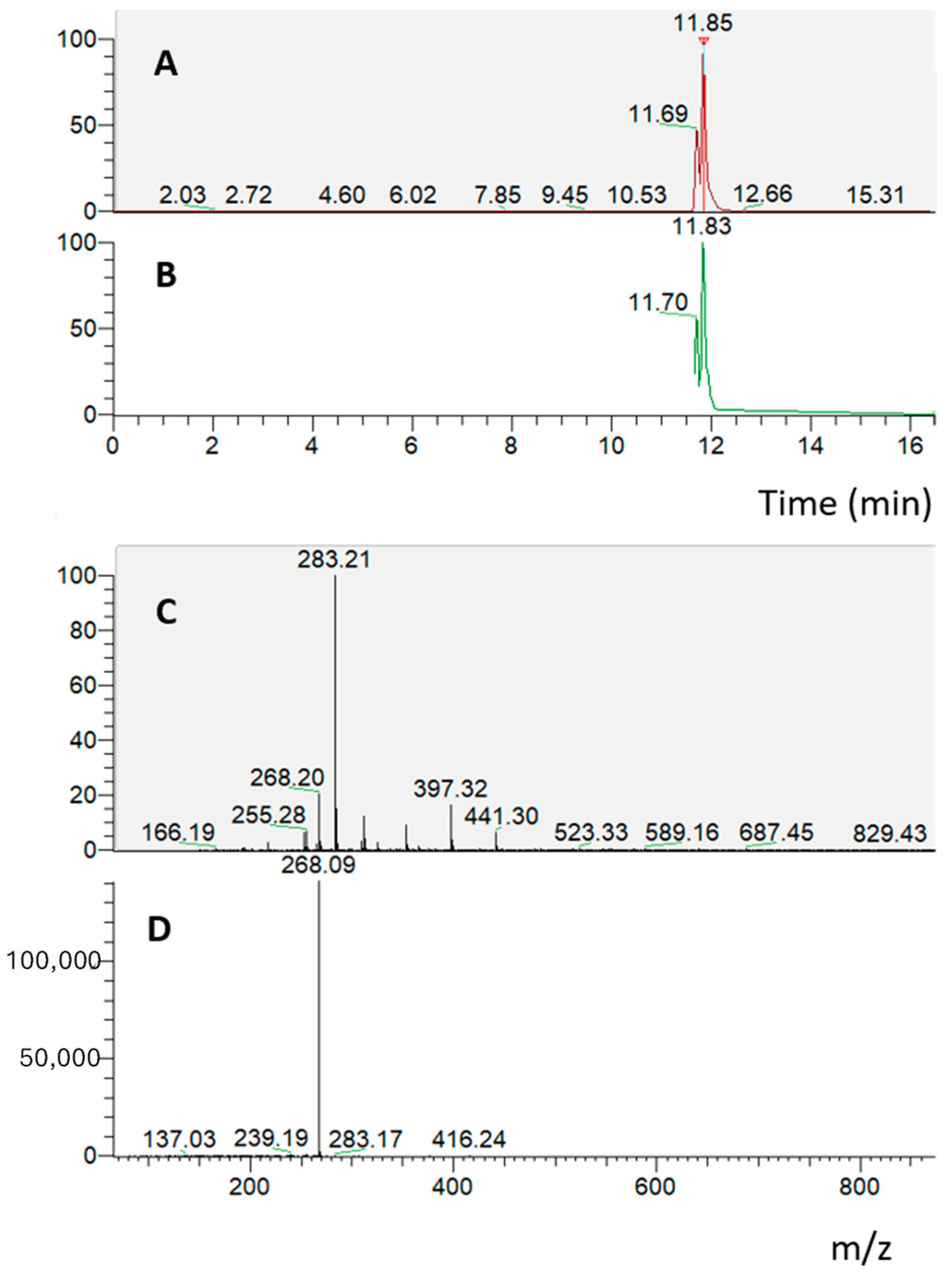
4.2.4. RP-UHPLC-PDA-ESI-MS/MS Analysis
RP-UHPLC-PDA-ESI-MS/MS analyses were performed using an Ultimate 3000 SD UHPLC system, equipped with a quaternary pump, autosampler, and column heater compartment. The chromatographic system was connected to a Surveyor UV–Vis photodiode array detector (PDA), using an ESI source at an LTQ XL Linear Ion Trap Mass Spectrometer (Thermo Fischer Scientific, Waltham, MA, USA). For UHPLC analysis a Kinetex® EVOTM 150 mm × 2.1 mm, 2.6 µm (L × I.D, particle size, Phenomenex®, Bologna, Italy) column was employed at a flow rate of 0.4 mL/min. The mobile phases consisted of (A) 0.1% CH3COOH in H2O and (B) can plus 0.1% CH3COOH. Analysis was performed in gradient as follows: 0–10.0 min, 2–35% B; 10–12 min, 35–50% B; 12–13 min, 50–60%; 13–25 min, 70–75% B; 25–30 min, 75–95% B; 30–31 min, 95–5% B; 31–32 min, 95–2% B; hold for 2 min; return to initial conditions in 0.1 min. Column oven was set to 40 °C, and 5 µL of extracts were injected. Chromatogram measurements were taken at 254, 280, and 324 nm; spectral data were taken between 200 and 800 nm for every peak. HRMS analysis was performed with a full MS scan (m/z 100–850) and data-dependent acquisition (dd-MS2 top N = 5). A resolution of 70,000 and 15,000 FWHM at m/z 200 was selected. A collision energy (NCE) value of 35 were used. Negative ESI was employed. Aliquots of 10 mg of freeze-dried digested samples were dissolved in 2 mL of H2O/MeOH: 50/50 prior to chromatographic analysis.
4.3. In Vitro Bioavailability
4.3.1. Cell Culture
Human colon carcinoma Caco-2 (ATCC, HTB-37tm), from the American Type Culture Collection, was cultivated in Dulbecco’s modified Eagle’s medium (DMEM) media supplemented with 10% fetal bovine serum (FBS), 1% nonessential amino acids (Gibco), 100 U/mL penicillin, and 100 μg/mL streptomycin. The cells were kept at 37 °C in a 5% CO2 incubator and the complete media was replaced every 2 days before reaching 90% confluence.
4.3.2. Cytotoxic Activity of Duodenal Digested S. lateriflora Extract on Caco-2 Cells
The cell viability assay (MTT test) was also performed on the Caco-2 cell line, to determine the maximum noncytotoxic concentration of duodenal digested
S. lateriflora extract. The MTT assay measures cellular metabolic activity as an indicator of vitality, proliferation, and cellular cytotoxicity [
47]. Cells were cultured in DMEM and supplemented with 1% penicillin-streptomycin and 10% fetal bovine serum, at 37 °C with 5% CO
2 in a humid environment, as previously described. A density of 3
× 10
4 cells/well was seeded into 96-well plates, and cells were cultured at 37 °C with 5% CO
2 in a humid environment for 4 h, as previously described. After 4 h of incubation, the cells were treated with different concentrations of duodenal digested
S. lateriflora extract (60–7.5 mg/mL). After the incubation period, the culture medium was removed from each well and the cells rinsed with phosphate-buffered saline (PBS) before adding the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT, Sigma-Aldrich, St. Louis, MO, USA) at a final concentration of 0.5 mg/mL. The cells were incubated for 3 h at 37 °C. The formazane crystals were then solubilized with 100 μL DMSO/well for 10 min and the viability rate was recorded at OD at 570 nm using a Multiskan™ GO microplate reader (Thermo Scientific Microplate Spectrophotometer, Thermo Fisher Scientific, Waltham, MA, USA). The experiment was performed in triplicate.
4.3.3. PAMPA Assay
The PAMPA assay is a noncell-based assay, based on the ability of the compound to diffuse from a donor compartment into an acceptor compartment, through a PVDF membrane filter, pretreated with a lipid solution dissolved in organic solvent. The test was conducted following the Manufacturer’s instructions (MultiScreen Permeability Plates from MilliPore Sigma, Burlington, MA, USA). Therefore, a 1% solution (
w/
v) of lecithin in dodecane was prepared to mimic the phospholipid membrane and sonicated until completely dissolved.
S. lateriflora samples were prepared and 1 mg of the duodenal digested sample was weighed and resuspended in 1 mL of PBS 5% DMSO pH 7.4 (buffer solution) to obtain a stock solution at a final concentration of 10 mg/mL. Finally, each test used a positive control with high permeability (caffeine) and a negative control with low permeability (furosemide) as according to the Biopharmaceutical Classification System (BCS), both solubilized in the same conditions of the duodenal digested
S. lateriflora extract. An aliquot of a 5 uL lecithin/dodecane mixture was carefully pipetted into each Donor plate. Next, 300 μL of PBS 5% DMSO was added to each well of the Acceptor plate, and 150 μL of the duodenal digested
S. lateriflora samples were added into each well of the Donor compartment. After that, the Donor and Acceptor plates were assembled, making sure the underside of the membrane was in contact with the buffer solution, and incubated at room temperature for 16 h. At the end of the incubation period, 100 μL/well from the Donor compartment and 250 μL from the Acceptor compartment were taken to measure UV/Vis absorption from 250 to 500 nm. In addition, to calculate the log P
e (log of the effective permeability of the compounds) as follows below, the theoretical equilibrium concentrations for each compound (and relative concentration tested) were measured.
To evaluate membrane integrity, calibration curves for caffeine and furosemide were prepared at concentrations ranging from 500 to 7.81 μM for the caffeine solution and from 500 to 31.25 μM for the furosemide solution. Spectrophotometric analysis was perfomed using a multiplate reader (Perkin Elmer, Waltham, MA, USA) and a 96-well quartz plate (Hellma Analytics, Müllheim, Germany).
4.3.4. Caco-2 Transwell Model System
For the transport experiments, the protocol according to Husbatrsch et al. [
48] was followed, with some modifications. Caco-2 cells were seeded at a density of 3 × 10
5 cells/well on transwell PET inserts (0.4 μm pore size, CellQART
®-Sterlitech, Auburn, WA, USA) and cultured in complete DMEM medium for 21 days, with the medium being changed three times a week. The transport experiment was performed in the apical-to-basolateral direction, with 0.5 mL of medium in the apical compartment and 1.5 mL medium in the basolateral compartment. Before carrying out the experiments, the monolayer integrity was evaluated by measuring the trans-epithelial electrical resistance (TEER) with an EVOM2™ Epithelial Voltohmmeter (World Precision Instrument, Sarasota, FL, USA). TEER was measured at days 7, 15 and 21 and TEER was measured after subtraction of the intrinsic resistance of the cell free insert according with the following formula:
Moreover, once the permeability test was completed, the permeability of a paracellular marker, LY (Invitrogen, Waltham, MA, USA, Lucifer Yellow CH, St. Louis, MO, USA, Lithium Salt, Qinghai, China), was assessed by analyzing the fluorescence using a fluorescence microplate reader (Envision microplate reader, Perkin Elmer, Waltham, MA, USA) at an excitation wavelength of 425 nm and an emission wavelength of 528 nm. The confluent monolayers were then used for further experiments.
Prior to the permeability assay, the inserts were washed twice and equilibrated for 30 min with prewarmed transport medium, Hank’s balanced salt solution (HBSS), pH 7.4. Thus, cells were treated with the duodenal digested S. lateriflora extract at a final concentration of 10 mg/mL and the transwell plates were incubated at 37 °C under agitation (VWR® Microplate Mixer, Radnor, PA, USA). After 30, 60, 90, 150 and 210 min, aliquots (200 μL) were collected from the acceptor compartment and replaced with the same volume of prewarmed buffered HBSS to maintain sink conditions. Finally, whole medium was collected from the donor compartment and submitted to the chromatographic analysis.
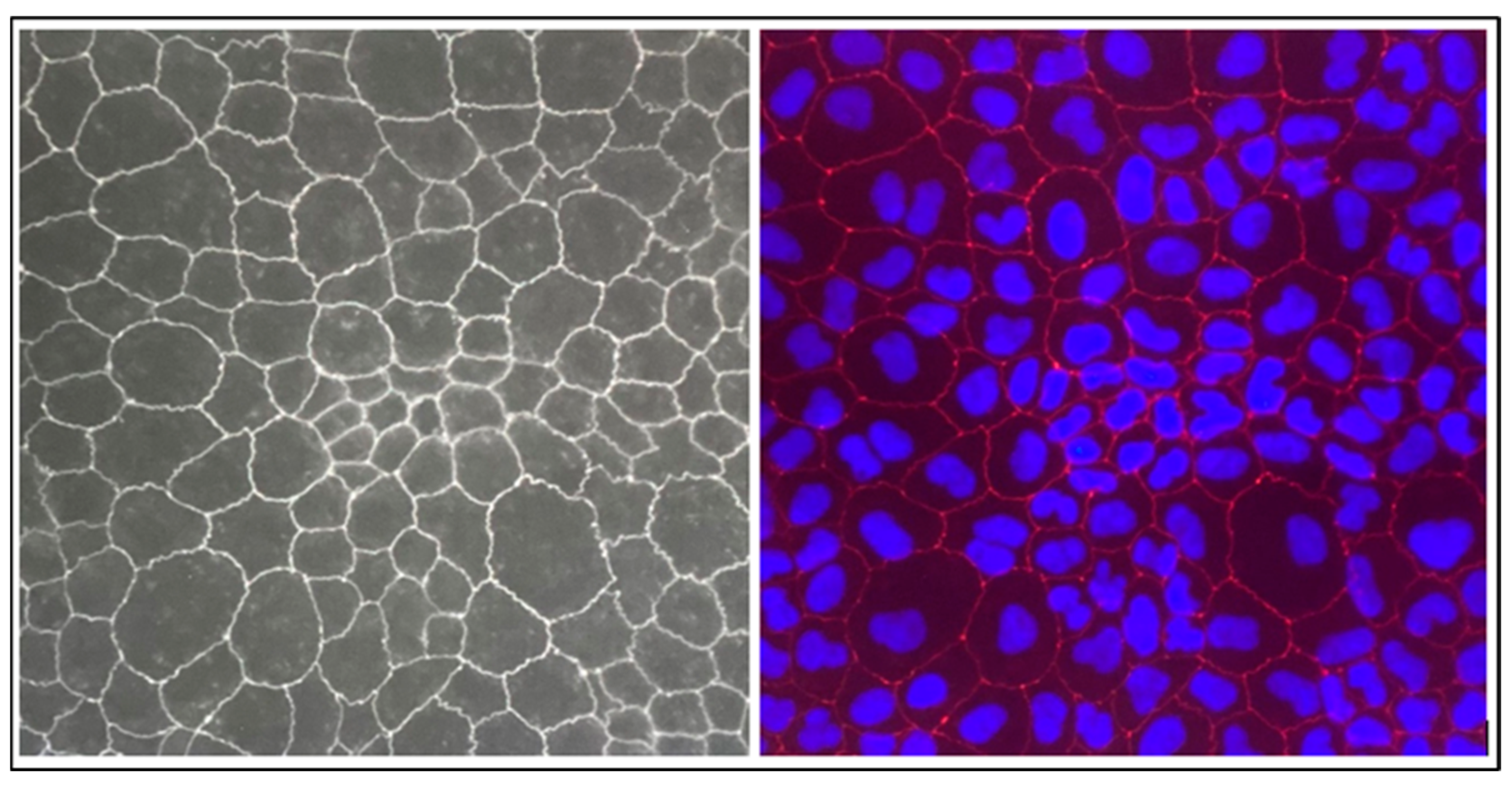
4.3.5. Immunochemistry
After 21 days culture, the medium in the 12-well transwell chamber was exhausted, and cells were washed three times with 500 μL PBS for 5 min each. Then, 500 μL paraformaldehyde 4% was added to each chamber, and the cells were fixed for 10 min. After permeabilization for 10 min with 0.25% Triton X-100 (in PBS), cells were blocked with 5% BSA in PBS for 1 h at room temperature. A working solution of ZO-1/TJP1 Antibody, Alexa Fluor® 594 conjugate (ZO1-1A12), was prepared at a concentration of 5 µg/mL in 1% BSA from a more concentrated stock solution. Subsequently, cells were labelled and incubated for 3 h at room temperature. Afterward, DAPI (Invitrogen) was used for counterstaining of the nucleus. Finally, each insert membrane was cut out, placed between two cover slips, and analyzed using a fluorescent optical microscope (DM2000 LED-Leica, Leica Microsystems, Wetzlar, Germany).
4.4. In Vitro Efficacy of S. lateriflora Extract on the Release of Cortisol
4.4.1. Cell Culture
H295R cells were maintained in Dulbecco’s modified Eagle’s medium/Hams’s F-12 (DMEM-F12) supplemented with 5% fetal bovine serum (FBS), insulin, apotransferrin, selenium and antibiotic (penicillin/streptomycin 100 g/mL). An atmosphere of 5% CO2 at 37 °C was provided and the medium was replaced every 48 h.
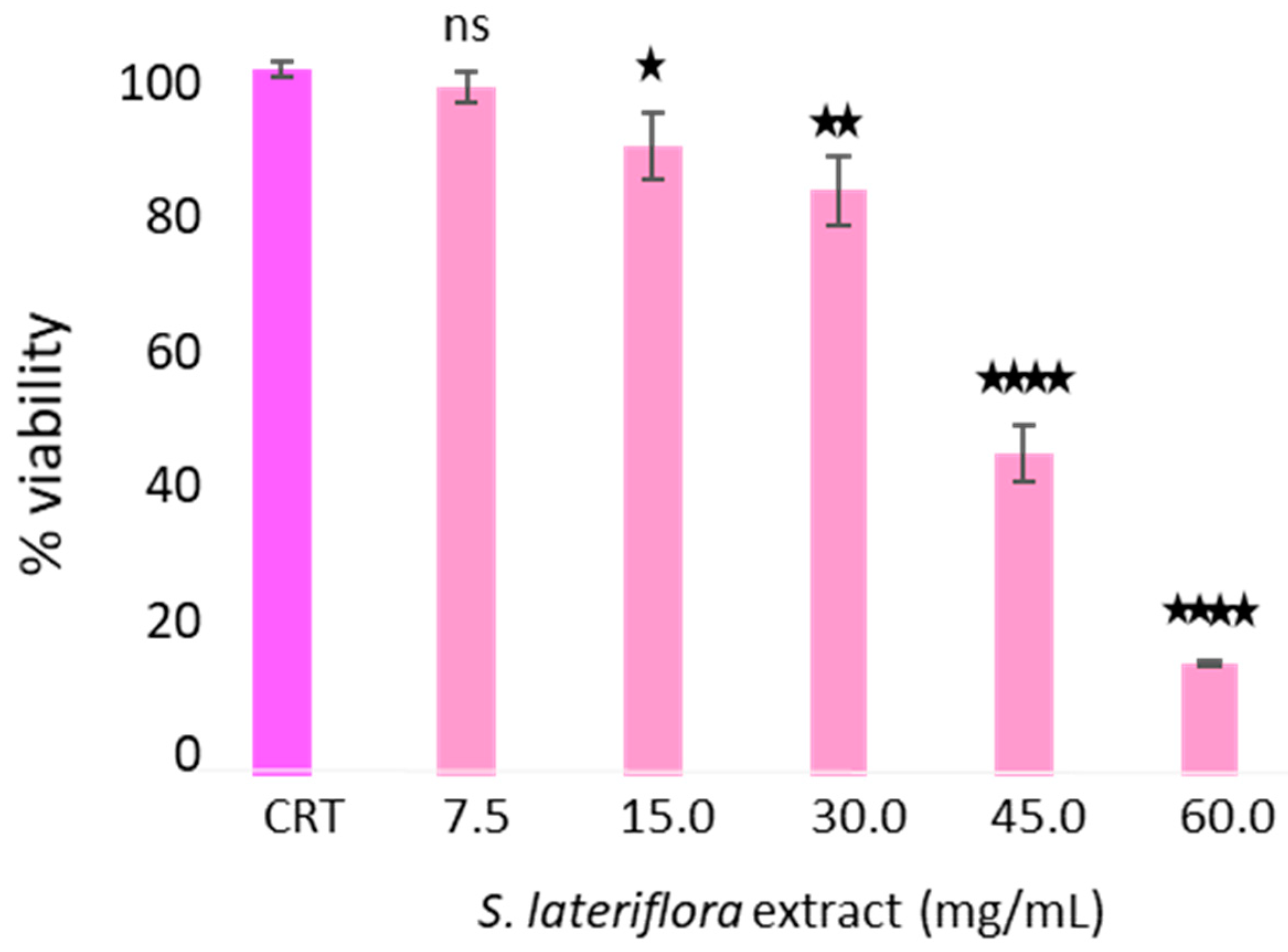
4.4.2. Cytotoxic Activity of S. lateriflora Extract on H295R Cells
To assess the maximum noncytotoxic concentration of
S. lateriflora extract on the H295R cell line, a 3-(4,5-dimethylthiazol-2-yl)-2,5-iphenyltetrazolium bromide (MTT) assay was performed. The MTT assay measures cellular metabolic activity as an indicator of vitality, proliferation, and cellular cytotoxicity [
47]. Cells were cultured in DMEM-F12 and supplemented with 5% fetal bovine serum, insulin, apotransferrin, selenium and 1% penicillin-streptomycin, at 37 °C, with 5% CO
2 in a humid environment, as previously described.
Thus, a density of 5 × 104 cells/well was seeded into 96-well plates and cells cultured at 37 °C with 5% CO2 in a humid environment for 24 h. The cells were treated with different concentrations of S. lateriflora extract (7.5–60.0 mg/mL). After the incubation period, the cells were treated with the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT, Sigma-Aldrich, St. Louis, MO, USA) with a final concentration of 0.5 mg/mL. The cells were incubated for 3 h at 37 °C. The formazane crystals were then solubilized with 100 μL DMSO/well for 10 min, and the viability rate was recorded at OD at 570 nm using a Multiskan™ GO microplate reader (Thermo Scientific Microplate Spectrophotometer). The current experiment was performed in triplicate.
4.4.3. In Vitro Evaluation of the S. lateriflora Extract on Inhibition of CORTISOL Release
In order to evaluate the effects of S. lateriflora extract on the release of cortisol, in vitro experiments were carried out by treating the cells with forskolin (20 μmol/L) to stimulate the production of cAMP and therefore steroidogenesis. Cells were seeded in sterile 6-multiwell plates in DMEM-F12 medium, supplemented with 5% FBS at a density of 4 × 105 cells. The cells were left in the incubator for 48 h. Subsequently, cells were harvested using low serum concentration medium (0.1% FBS) and the cells were incubated overnight at 37 °C with 5% CO2, followed by replacement of the test medium containing 0.1% FBS, varying S. lateriflora extract treatment concentrations (5.0–30.0 mg/mL) and forskolin. After 24 h of incubation, the cell culture medium was collected, and the cortisol concentration was analyzed through ELISA assay in accordance with the manufacturer’s instructions. The treatment and the determination of cortisol were performed in triplicate.
4.4.4. Evaluation of Cortisol Levels by ELISA Assay
The concentration of cortisol in H295R cell culture supernatant samples was evaluated using the enzyme-linked immunosorbent assay (ELISA) with commercially available test kits. Samples were diluted where necessary; the standards of known concentration and the blanks were prepared for the elaboration of calibration lines. Samples and standards were incubated together with the primary antibody, and then with the secondary antibody in multiwell plates. Finally, the peroxidase enzyme substrate conjugated to the secondary antibody was added to develop a colored product. The absorbance values are related to the cortisol concentration according to an exponential trend. The absorbance was recorded by spectrophotometer at a wavelength of 450 nm. The data obtained were processed with the MyAssay software (
https://www.myassays.com/index.html, accessed on 22 January 2024).
4.5. Statistical Analysis
For the MTT assay performed on the H295R cell line and the Caco2 cell line, the concentrations of cortisol tested with the ELISA assay, and the percentages of degradation of the S. lateriflora extract components analyzed by RP-UHPLC-PDA-ESI-MS/MS, data were expressed as mean ± standard deviation (SD) from three biological replicates.
Concerning the viability percentages of the H295R cell line and the Caco2 cell line, and the concentrations of cortisol, the statistical comparison among groups was conducted using one-way ANOVA followed by Dunnett’s post hoc test for multiple comparisons to determine significance, which was set to p < 0.05. The statistical analyses were performed using GraphPad Prism, version 8 (San Diego, CA, USA).



 ,
, 









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}