Effects of Genetically Engineered Stem Cells Expressing Cytosine Deaminase and Interferon-Beta or Carboxyl Esterase on the Growth of LNCaP Prostate Cancer Cells


Abstract
:1. Introduction
2. Results
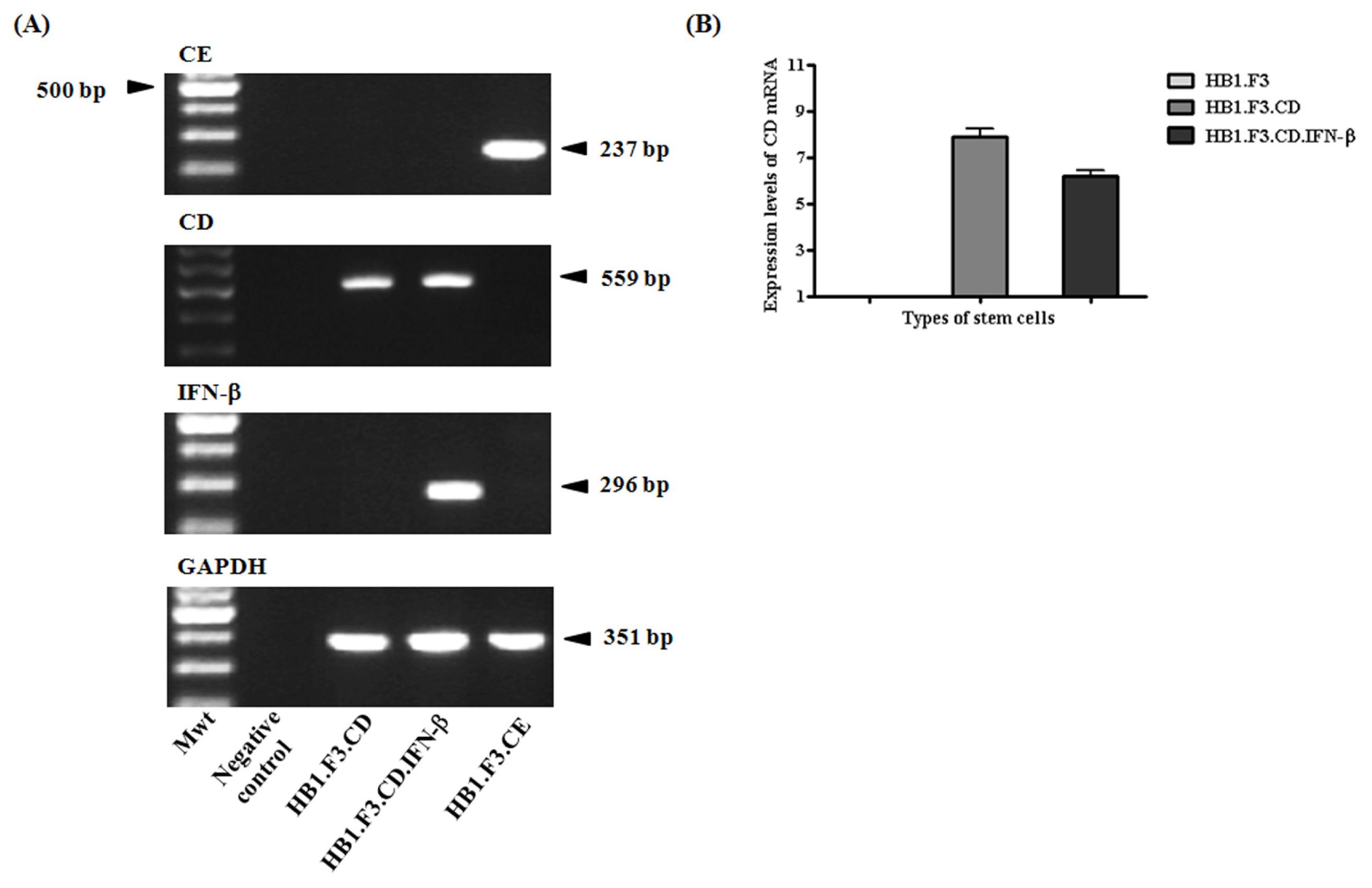
2.1. Expression of Therapeutic Genes in the Genetically Engineered Stem Cells
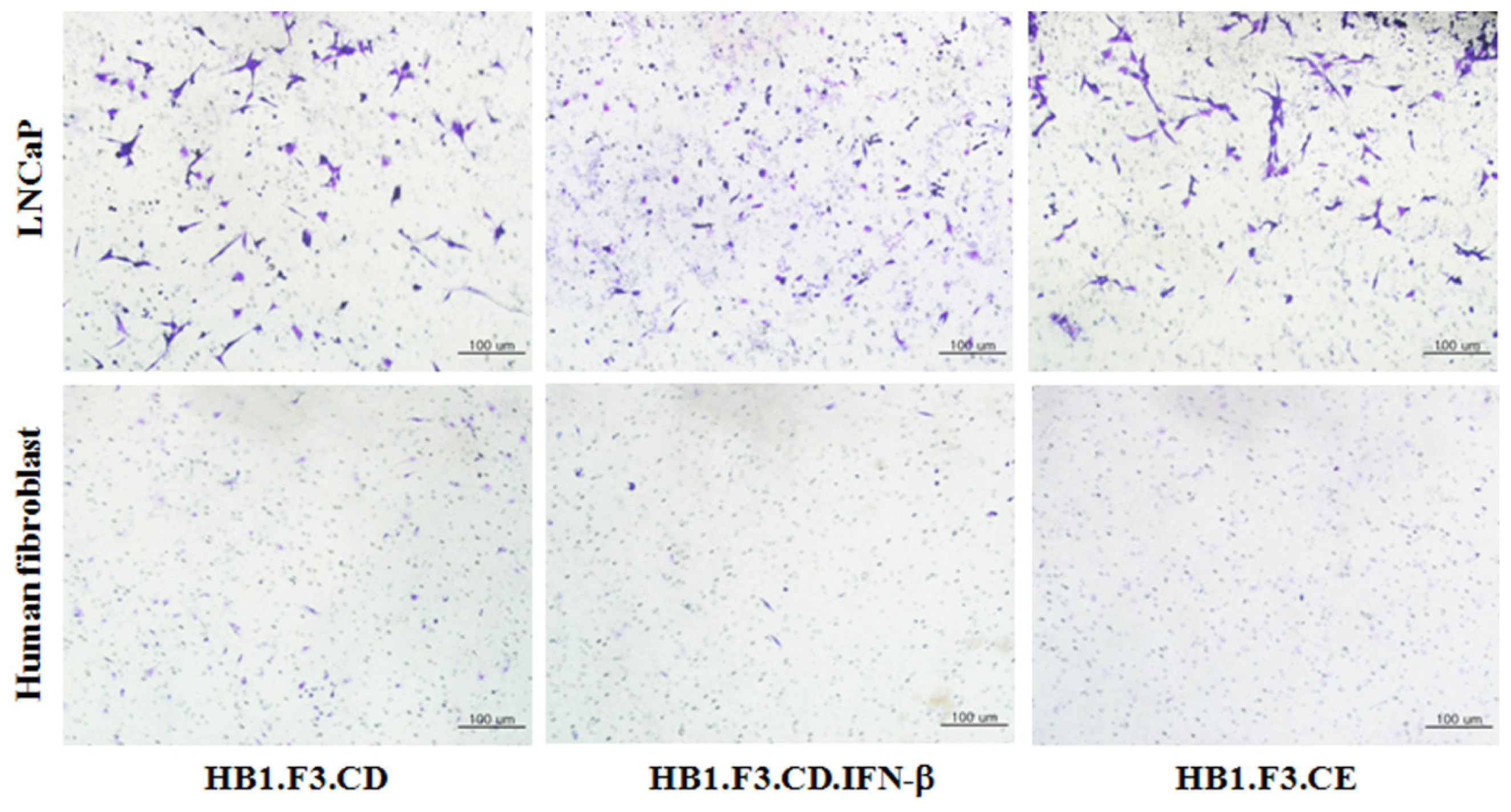
2.2. Tumor-Tropic Effects of the Engineered Stem Cells against Prostate Cancer Cells
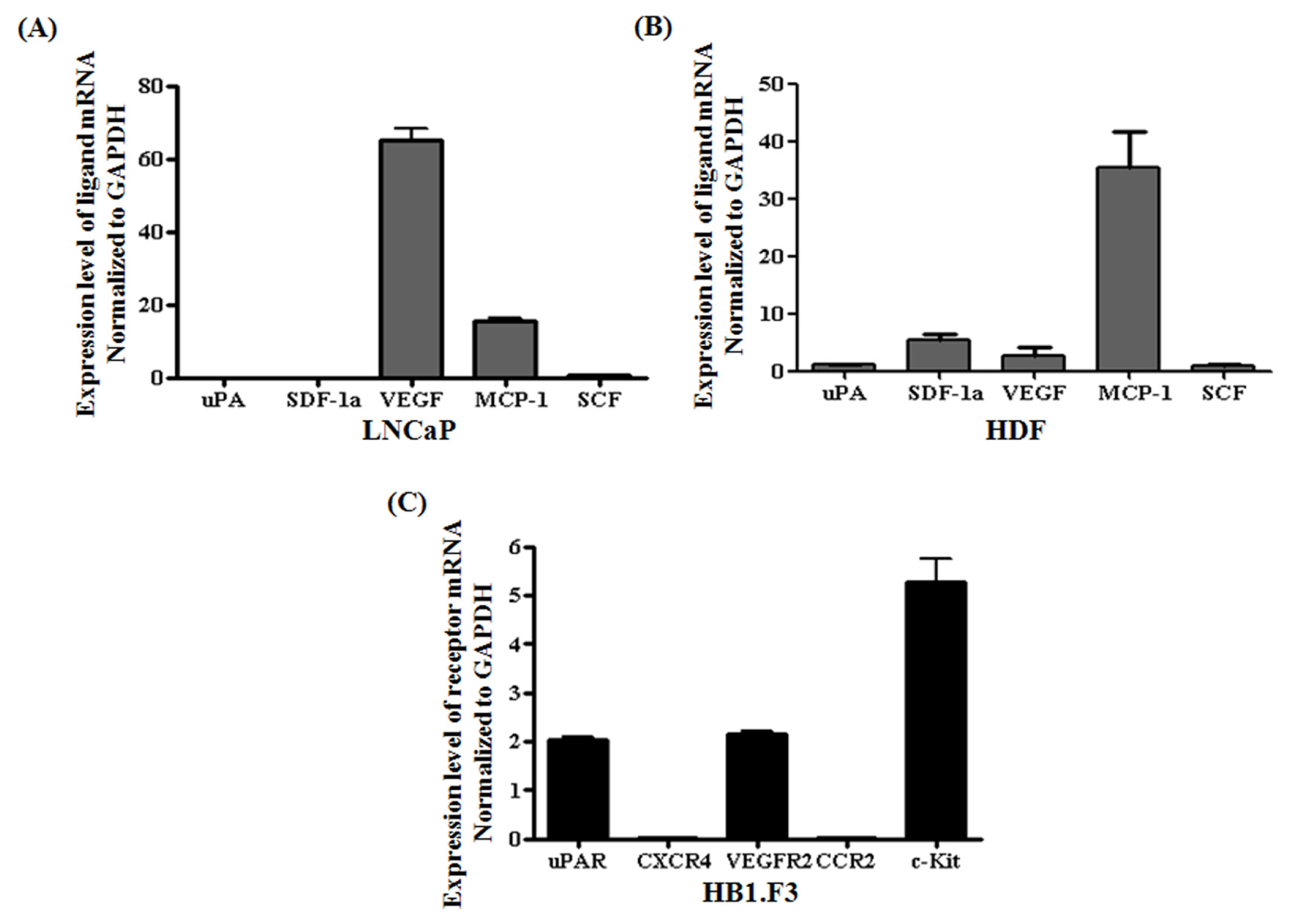
2.3. Chemoattractant Ligands and Receptors Regulating Stem Cells Migration
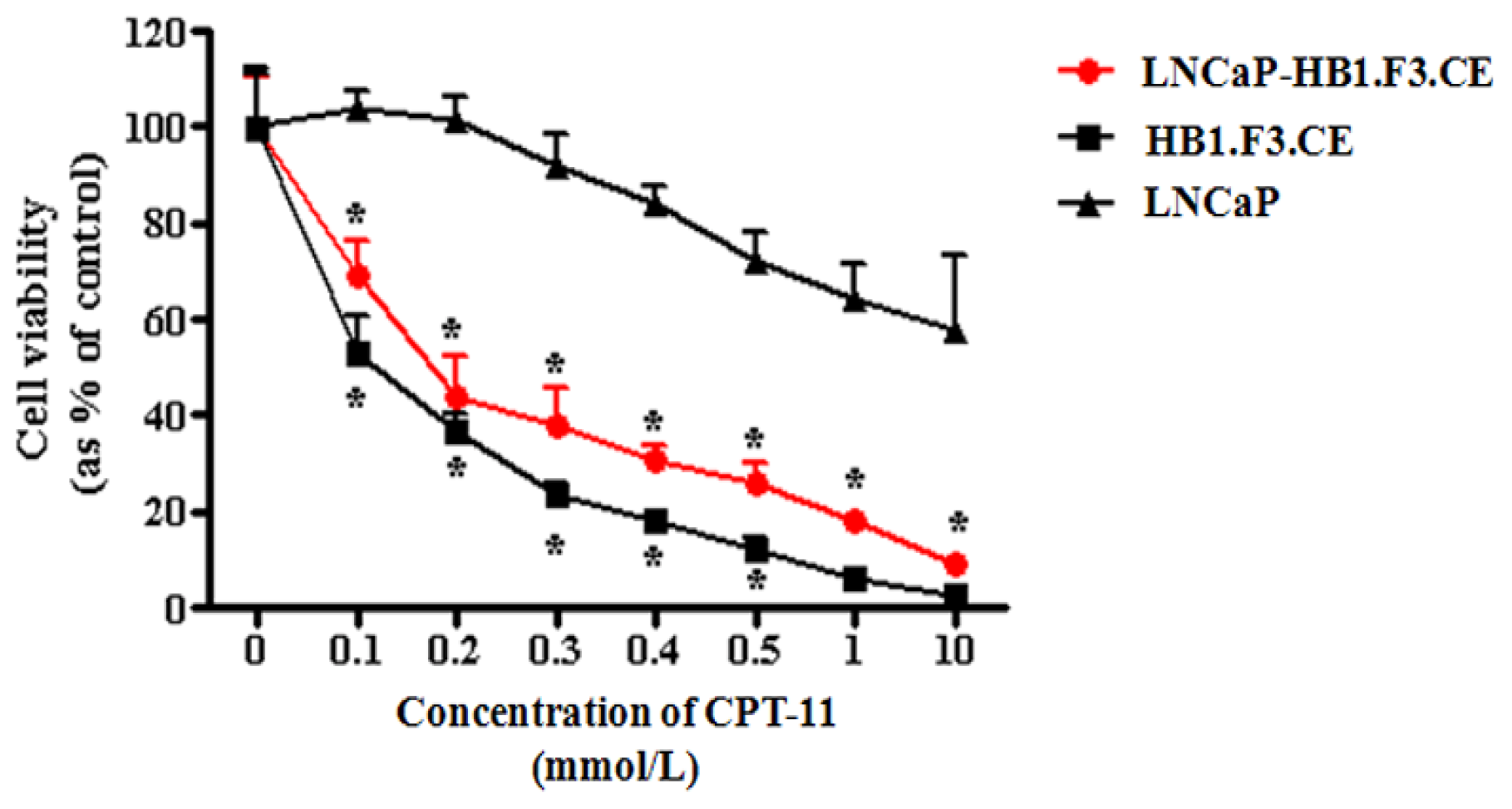
2.4. Proliferation of Prostate Cancer Cells Inhibited by CE Gene Expression in the HB1.F3.CE Cells
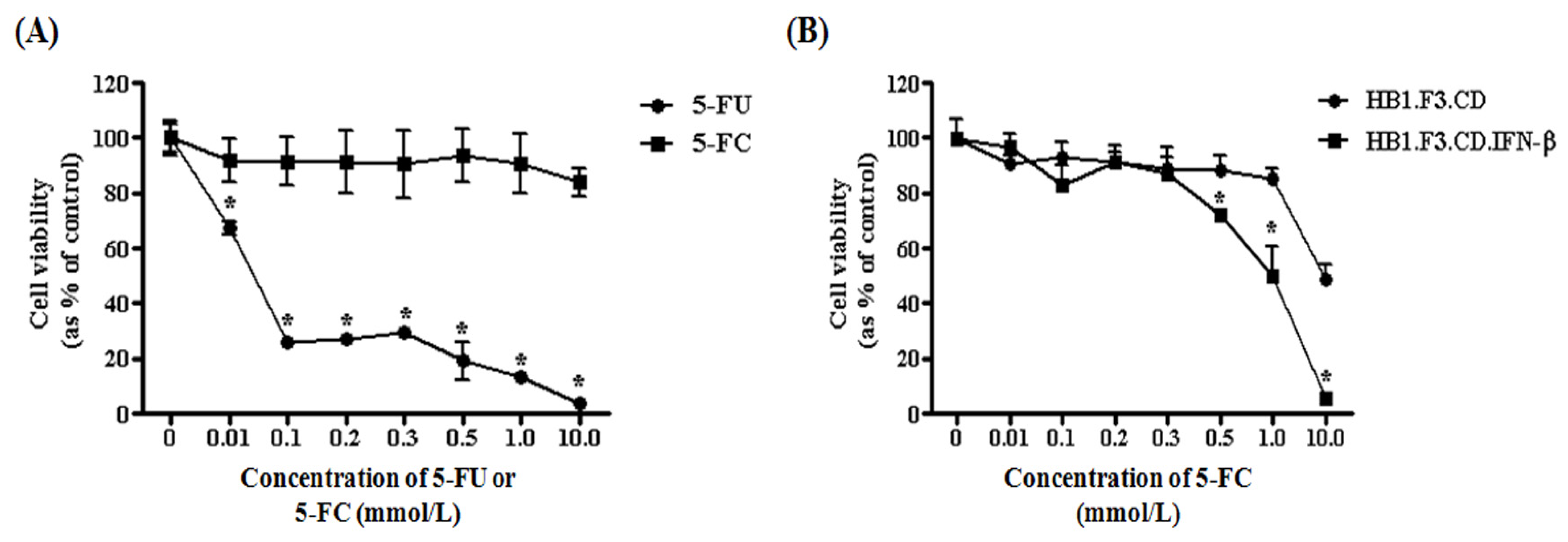
2.5. Antitumor Effects of HB1.F3.CD or HB1.F3.CD.IFN-β Cells
3. Experimental Section
3.1. Cell Lines and Cell Culture
3.2. Semi-Quantitative RT-PCR
3.3. Quantitative Real-Time PCR (qRT-PCR)
3.4. In vitro Migration Assay
3.5. Cell Proliferation Assay
3.6. Statistic Analysis
4. Discussion and Conclusions
Acknowledgments
References
- Jemal, A.; Murray, T.; Samuels, A.; Ghafoor, A.; Ward, E.; Thun, M.J. Cancer statistics, 2003. CA Cancer J. Clin 2003, 53, 5–26. [Google Scholar]
- Eberl, M.M.; Sunga, A.Y.; Farrell, C.D.; Mahoney, M.C. Patients with a family history of cancer: Identification and management. J. Am. Board Fam. Pract 2005, 18, 211–217. [Google Scholar]
- Martiniello-Wilks, R.; Dane, A.; Voeks, D.J.; Jeyakumar, G.; Mortensen, E.; Shaw, J.M.; Wang, X.Y.; Both, G.W.; Russell, P.J. Gene-directed enzyme prodrug therapy for prostate cancer in a mouse model that imitates the development of human disease. J. Gene Med 2004, 6, 43–54. [Google Scholar]
- Venara, A.; Thibaudeau, E.; Lebdai, S.; Mucci, S.; Ridereau-Zins, C.; Azzouzi, R.; Hamy, A. Rectal metastasis of prostate cancer: About a case. J. Clin. Med. Res 2010, 2, 137–139. [Google Scholar]
- Somers, K.D.; Brown, R.R.; Holterman, D.A.; Yousefieh, N.; Glass, W.F.; Wright, G.L., Jr; Schellhammer, P.F.; Qian, J.; Ciavarra, R.P. Orthotopic treatment model of prostate cancer and metastasis in the immunocompetent mouse: Efficacy of flt3 ligand immunotherapy. Int. J. Cancer 2003, 107, 773–780. [Google Scholar]
- Grubb, R.L.; Calvert, V.S.; Wulkuhle, J.D.; Paweletz, C.P.; Linehan, W.M.; Phillips, J.L.; Chuaqui, R.; Valasco, A.; Gillespie, J.; Emmert-Buck, M.; et al. Signal pathway profiling of prostate cancer using reverse phase protein arrays. Proteomics 2003, 3, 2142–2146. [Google Scholar]
- Petrylak, D.P.; Tangen, C.M.; Hussain, M.H.; Lara, P.N., Jr; Jones, J.A.; Taplin, M.E.; Burch, P.A.; Berry, D.; Moinpour, C.; Kohli, M.; et al. Docetaxel and estramustine compared with mitoxantrone and prednisone for advanced refractory prostate cancer. N. Engl. J. Med. 2004, 351, 1513–1520. [Google Scholar]
- Satoh, T.; Irie, A.; Egawa, S.; Baba, S. In situ gene therapy for prostate cancer. Curr. Gene Ther 2005, 5, 111–119. [Google Scholar]
- Eder, I.E.; Haag, P.; Bartsch, G.; Klocker, H. Gene therapy strategies in prostate cancer. Curr. Gene Ther 2005, 5, 1–10. [Google Scholar]
- Dearnaley, D.P.; Hall, E.; Lawrence, D.; Huddart, R.A.; Eeles, R.; Nutting, C.M.; Gadd, J.; Warrington, A.; Bidmead, M.; Horwich, A. Phase III pilot study of dose escalation using conformal radiotherapy in prostate cancer: PSA control and side effects. Br. J. Cancer 2005, 92, 488–498. [Google Scholar]
- Iversen, P.; Tyrrell, C.J.; Kaisary, A.V.; Anderson, J.B.; van Poppel, H.; Tammela, T.L.; Chamberlain, M.; Carroll, K.; Melezinek, I. Bicalutamide monotherapy compared with castration in patients with nonmetastatic locally advanced prostate cancer: 6.3 years of followup. J. Urol 2000, 164, 1579–1582. [Google Scholar]
- Djavan, B.; Nasu, Y. Prostate cancer gene therapy-what have we learned and where are we going? Rev. Urol 2001, 3, 179–186. [Google Scholar]
- Kang, N.H.; Hwang, K.A.; Yi, B.R.; Lee, H.J.; Jeung, E.B.; Kim, S.U.; Choi, K.C. Human amniotic fluid-derived stem cells expressing cytosine deaminase and thymidine kinase inhibits the growth of breast cancer cells in cellular and xenograft mouse models. Cancer Gene Ther 2012, 19, 412–419. [Google Scholar]
- Oosterhoff, D.; Pinedo, H.M.; van der Meulen, I.H.; de Graaf, M.; Sone, T.; Kruyt, F.A.; van Beusechem, V.W.; Haisma, H.J.; Gerritsen, W.R. Secreted and tumour targeted human carboxylesterase for activation of irinotecan. Br. J. Cancer 2002, 87, 659–664. [Google Scholar]
- Gardner, T.A.; Sloan, J.; Raikwar, S.P.; Kao, C. Prostate cancer gene therapy: Past experiences and future promise. Cancer Metastasis Rev 2002, 21, 137–145. [Google Scholar]
- You, M.H.; Kim, W.J.; Shim, W.; Lee, S.R.; Lee, G.; Choi, S.; Kim, D.Y.; Kim, Y.M.; Kim, H.; Han, S.U. Cytosine deaminase-producing human mesenchymal stem cells mediate an antitumor effect in a mouse xenograft model. J. Gastroenterol. Hepatol 2009, 24, 1393–1400. [Google Scholar]
- Parker, W.B.; Cheng, Y.C. Metabolism and mechanism of action of 5-fluorouracil. Pharmacol. Ther 1990, 48, 381–395. [Google Scholar]
- Wierdl, M.; Morton, C.L.; Danks, M.K.; Potter, P.M. Isolation and characterization of a cDNA encoding a horse liver butyrylcholinesterase: Evidence for CPT-11 drug activation. Biochem. Pharmacol 2000, 59, 773–781. [Google Scholar]
- Wierdl, M.; Tsurkan, L.; Hyatt, J.L.; Edwards, C.C.; Hatfield, M.J.; Morton, C.L.; Houghton, P.J.; Danks, M.K.; Redinbo, M.R.; Potter, P.M. An improved human carboxylesterase for enzyme/prodrug therapy with CPT-11. Cancer Gene Ther 2008, 15, 183–192. [Google Scholar]
- Yi, B.R.; Kang, N.H.; Hwang, K.A.; Kim, S.U.; Jeung, E.B.; Choi, K.C. Antitumor therapeutic effects of cytosine deaminase and interferon-beta against endometrial cancer cells using genetically engineered stem cells in vitro. Anticancer Res 2011, 31, 2853–2861. [Google Scholar]
- Aboody, K.S.; Najbauer, J.; Danks, M.K. Stem and progenitor cell-mediated tumor selective gene therapy. Gene Ther 2008, 15, 739–752. [Google Scholar]
- Bobis, S.; Jarocha, D.; Majka, M. Mesenchymal stem cells: Characteristics and clinical applications. Folia Histochem. Cytobiol 2006, 44, 215–230. [Google Scholar]
- Studeny, M.; Marini, F.C.; Dembinski, J.L.; Zompetta, C.; Cabreira-Hansen, M.; Bekele, B.N.; Champlin, R.E.; Andreeff, M. Mesenchymal stem cells: Potential precursors for tumor stroma and targeted-delivery vehicles for anticancer agents. J. Natl. Cancer Inst 2004, 96, 1593–1603. [Google Scholar]
- Yi, B.R.; Kim, S.U.; Kim, Y.B.; Lee, H.J.; Cho, M.H.; Choi, K.C. Antitumor effects of genetically engineered stem cells expressing yeast cytosine deaminase in lung cancer brain metastases via their tumor-tropic properties. Oncol. Rep 2012, 27, 1823–1828. [Google Scholar]
- Barkho, B.Z.; Munoz, A.E.; Li, X.; Li, L.; Cunningham, L.A.; Zhao, X. Endogenous matrix metalloproteinase (MMP)-3 and MMP-9 promote the differentiation and migration of adult neural progenitor cells in response to chemokines. Stem Cells 2008, 26, 3139–3149. [Google Scholar]
- Kim, K.Y.; Yi, B.R.; Lee, H.R.; Kang, N.H.; Jeung, E.B.; Kim, S.U.; Choi, K.C. Stem cells with fused gene expression of cytosine deaminase and interferon-beta migrate to human gastric cancer cells and result in synergistic growth inhibition for potential therapeutic use. Int. J. Oncol 2011, 40, 1097–1104. [Google Scholar]
- Yi, B.R.; O, S.N.; Kang, N.H.; Hwang, K.A.; Kim, S.U.; Jeung, E.B.; Kim, Y.B.; Heo, G.J.; Choi, K.C. Genetically engineered stem cells expressing cytosine deaminase and interferon-beta migrate to human lung cancer cells and have potentially therapeutic anti-tumor effects. Int. J. Oncol 2011, 39, 833–839. [Google Scholar]
- Kim, K.Y.; Kim, S.U.; Leung, P.C.; Jeung, E.B.; Choi, K.C. Influence of the prodrugs 5-fluorocytosine and CPT-11 on ovarian cancer cells using genetically engineered stem cells: Tumor-tropic potential and inhibition of ovarian cancer cell growth. Cancer Sci 2010, 101, 955–962. [Google Scholar]
- Dhanasekaran, S.M.; Barrette, T.R.; Ghosh, D.; Shah, R.; Varambally, S.; Kurachi, K.; Pienta, K.J.; Rubin, M.A.; Chinnaiyan, A.M. Delineation of prognostic biomarkers in prostate cancer. Nature 2001, 412, 822–826. [Google Scholar]
- Imitola, J.; Raddassi, K.; Park, K.I.; Mueller, F.J.; Nieto, M.; Teng, Y.D.; Frenkel, D.; Li, J.; Sidman, R.L.; Walsh, C.A.; et al. Directed migration of neural stem cells to sites of CNS injury by the stromal cell-derived factor 1alpha/CXC chemokine receptor 4 pathway. Proc. Natl. Acad. Sci. USA 2004, 101, 18117–18122. [Google Scholar]
- Kang, N.H.; Yi, B.R.; Lim, S.Y.; Hwang, K.A.; Baek, Y.S.; Kang, K.S.; Choi, K.C. Human amniotic membrane-derived epithelial stem cells display anticancer activity in BALB/c female nude mice bearing disseminated breast cancer xenografts. Int. J. Oncol 2012, 40, 2022–2028. [Google Scholar]
- Choi, E.A.; Lei, H.; Maron, D.J.; Mick, R.; Barsoum, J.; Yu, Q.C.; Fraker, D.L.; Wilson, J.M.; Spitz, F.R. Combined 5-fluorouracil/systemic interferon-beta gene therapy results in long-term survival in mice with established colorectal liver metastases. Clin. Cancer Res 2004, 10, 1535–1544. [Google Scholar]
- Zhang, Y.; Chirmule, N.; Gao, G.P.; Qian, R.; Croyle, M.; Joshi, B.; Tazelaar, J.; Wilson, J.M. Acute cytokine response to systemic adenoviral vectors in mice is mediated by dendritic cells and macrophages. Mol. Ther 2001, 3, 697–707. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequence | Predicted product size (bp) | |
|---|---|---|---|
| Rabbit CE | reverse | CTCCAGCATCTCTGTGGTGA | 237 |
| forward | TGCTGGGCTATCCACTCTCT | ||
| Human CD | reverse | GCGCGAGTCACCGCCAGCCACCACGGC | 559 |
| forward | GTTTGTAATCGATGGCTTCTGGCTGC | ||
| Human IFN-β | reverse | AAAGAAGCAGCAATTTTCAG | 296 |
| forward | TTTCTCCAGTTTTTCTTCCA | ||
| Human GAPDH | reverse | ATGTTCGTCATGGGTGTGAACCA | 351 |
| forward | TGGCAGGTTTTTCTAGACGGCAG |
| Gene | Sequence | |
|---|---|---|
| uPA | reverse | GGCAGGCAGATGGTCTGTAT |
| forward | TTGCTCACCACAACGACATT | |
| uPAR | reverse | TCCCCTTGCAGCTGTAACACT |
| forward | GCCCAATCCTGGAGCTTGA | |
| SCF | reverse | GCCTTCAGAAATATTTGAAAACTTG |
| forward | GGCAAATCTTCCAAAAGACTACA | |
| c-Kit | reverse | TCACAGATGGTTGAGAAGAGCCT |
| forward | CGCCTGGGATTTTCTCTGC | |
| SDF-1 | reverse | TCCCATCCCACAGAGAGAAG |
| forward | GTGTCACTGGCGACACGTAG | |
| CXCR4 | reverse | GAGGGCCTTGCGCTTCTGGTG |
| forward | ATCCCTGCCCTCCTGCTGACTATTC | |
| VEGF | reverse | TCTTTCTTTGGTCTGCATTCACAT |
| forward | CCAGCACATAGGAGAGATGAGCTT | |
| VEGFR2 | reverse | AGCATGGAAGAGGATTCTGGACT |
| forward | CGGCTCTTTCGCTTACTGTTCT | |
| MCP-1 | reverse | TCTTCGGAGTTTGGGTTTGC |
| forward | CAAGCAGAAGTGGGTTCAGGA | |
| CCR-2 | reverse | ACATTTACAAGTTGCAGTTTTCAGC |
| forward | CTACCTTCCAGTTCCTCATTTTT |
© 2012 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Yi, B.-R.; Hwang, K.-A.; Kim, Y.-B.; Kim, S.U.; Choi, K.-C. Effects of Genetically Engineered Stem Cells Expressing Cytosine Deaminase and Interferon-Beta or Carboxyl Esterase on the Growth of LNCaP Prostate Cancer Cells. Int. J. Mol. Sci. 2012, 13, 12519-12532. https://doi.org/10.3390/ijms131012519
Yi B-R, Hwang K-A, Kim Y-B, Kim SU, Choi K-C. Effects of Genetically Engineered Stem Cells Expressing Cytosine Deaminase and Interferon-Beta or Carboxyl Esterase on the Growth of LNCaP Prostate Cancer Cells. International Journal of Molecular Sciences. 2012; 13(10):12519-12532. https://doi.org/10.3390/ijms131012519
Chicago/Turabian StyleYi, Bo-Rim, Kyung-A. Hwang, Yun-Bae Kim, Seung U. Kim, and Kyung-Chul Choi. 2012. "Effects of Genetically Engineered Stem Cells Expressing Cytosine Deaminase and Interferon-Beta or Carboxyl Esterase on the Growth of LNCaP Prostate Cancer Cells" International Journal of Molecular Sciences 13, no. 10: 12519-12532. https://doi.org/10.3390/ijms131012519