An Investigation of the Effects of Self-Assembled Monolayers on Protein Crystallisation

Abstract
:1. Introduction
2. Results
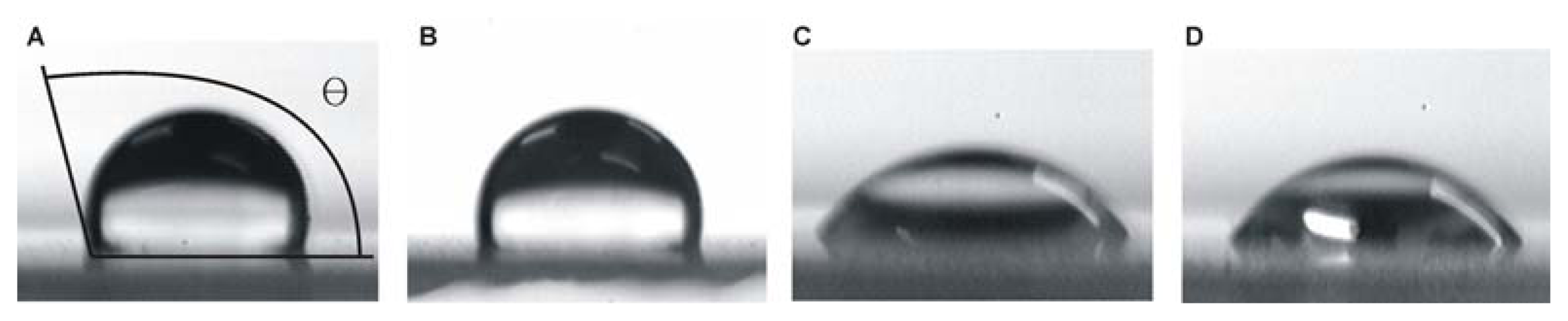
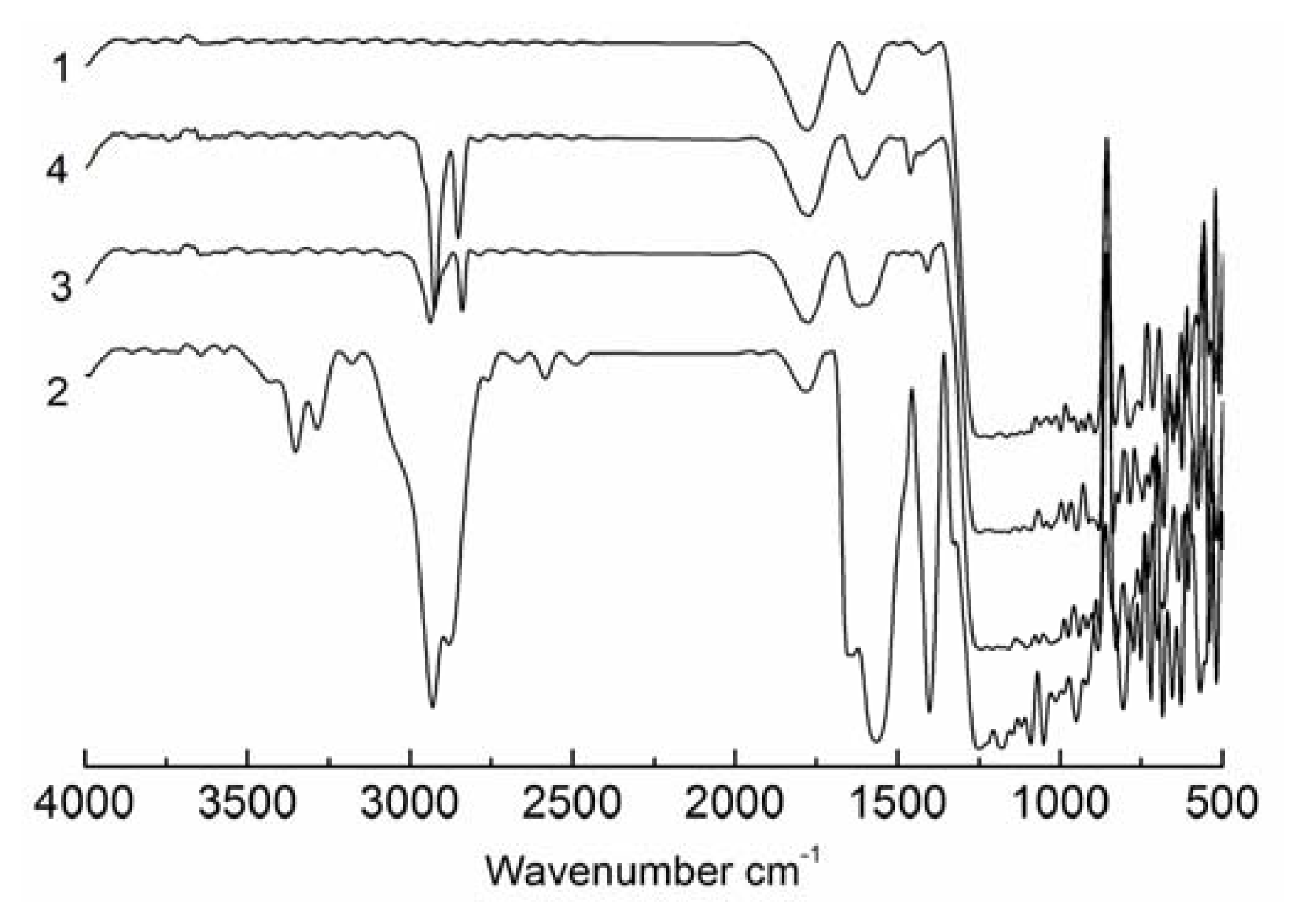
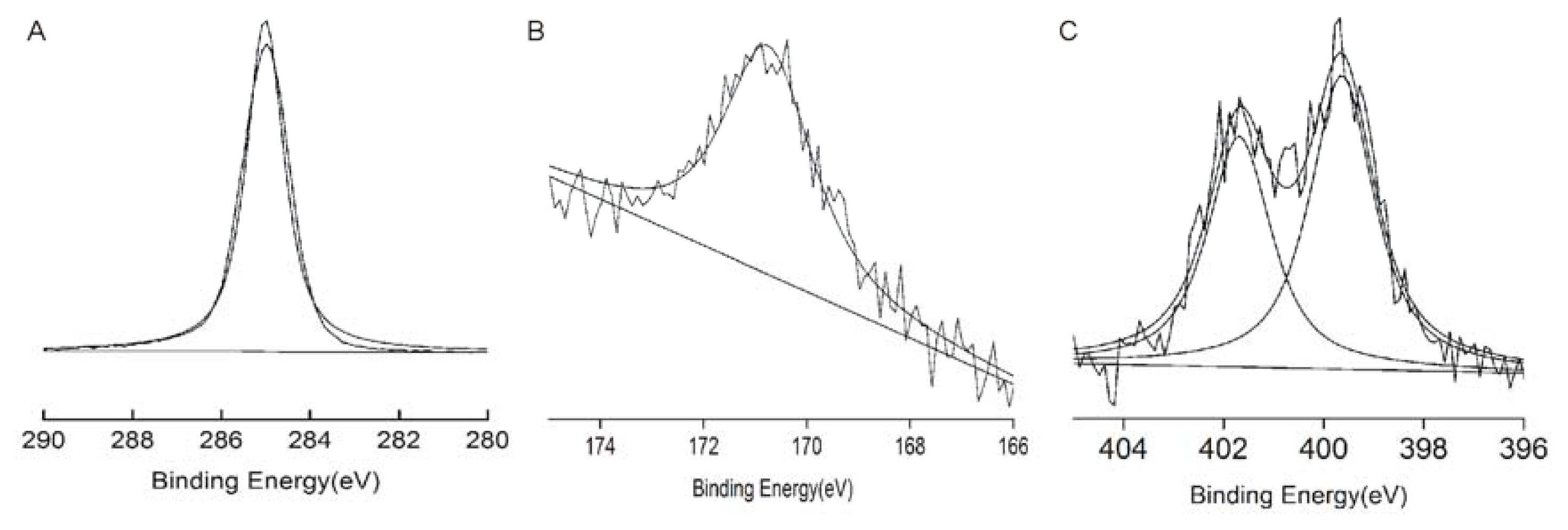
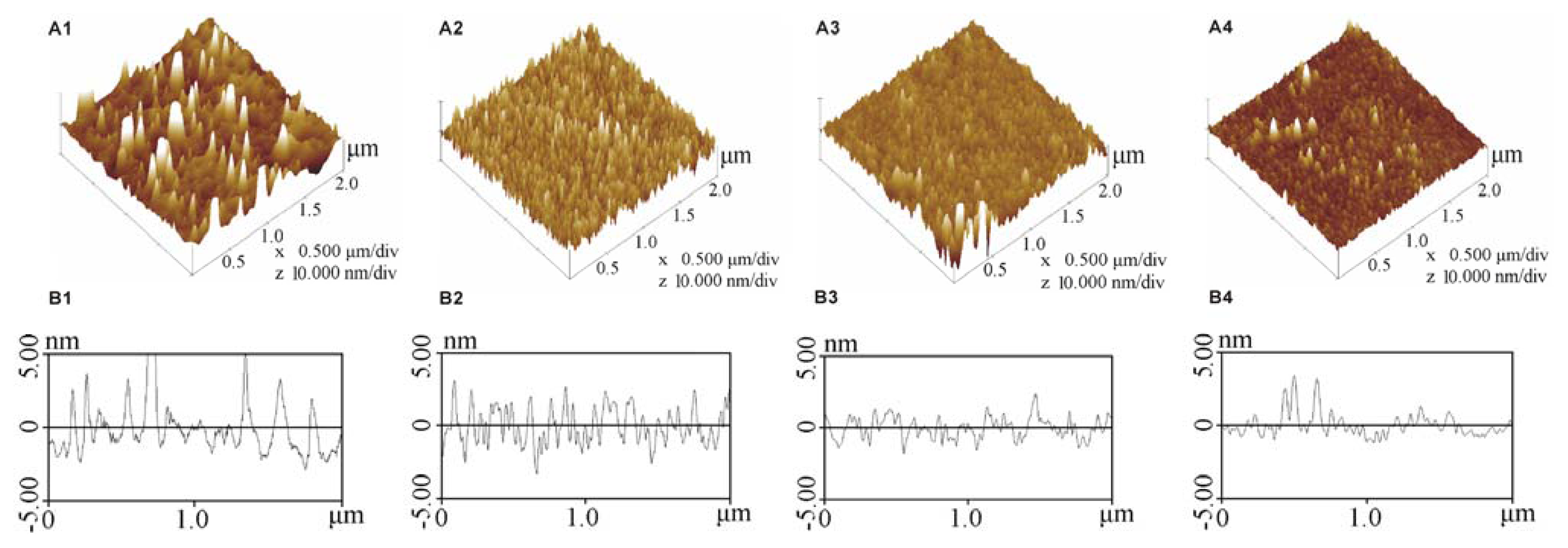
2.1. Characterisation of the Modified SAMs
2.2. Effect of the Modified SAM Cover Slips on the Reproducibility of Protein Crystallisation
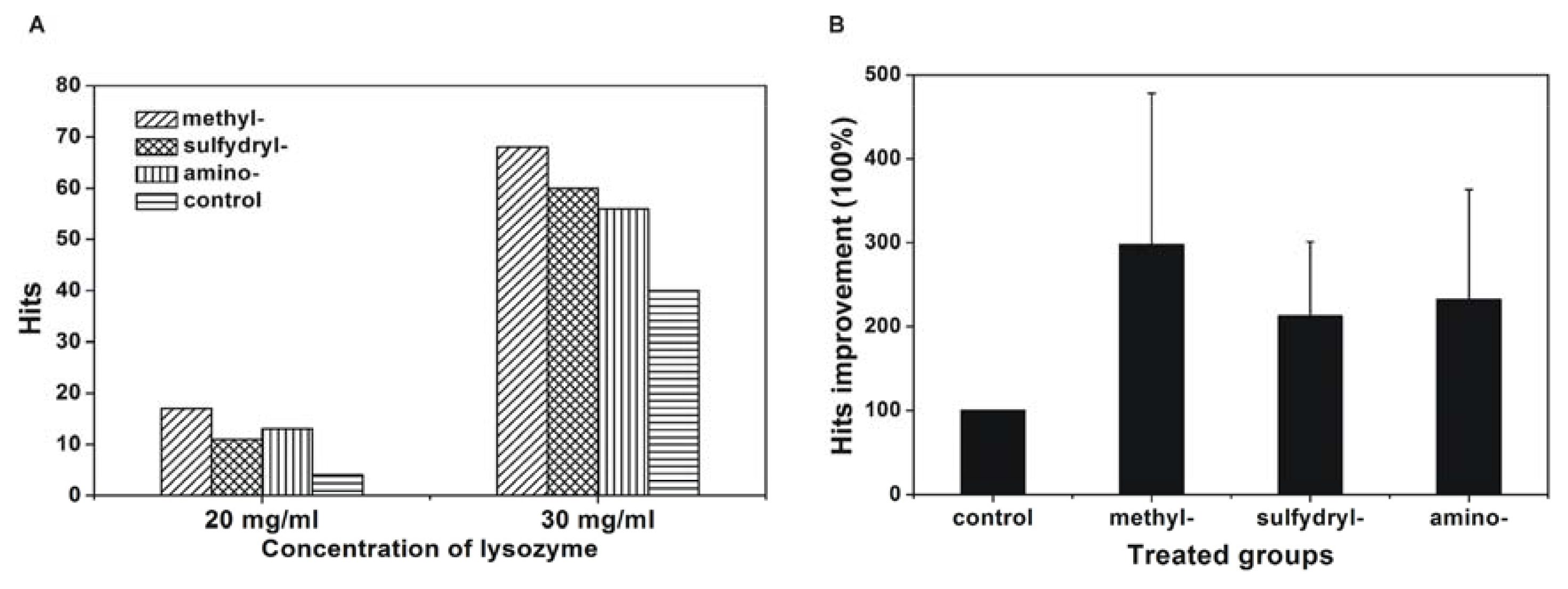
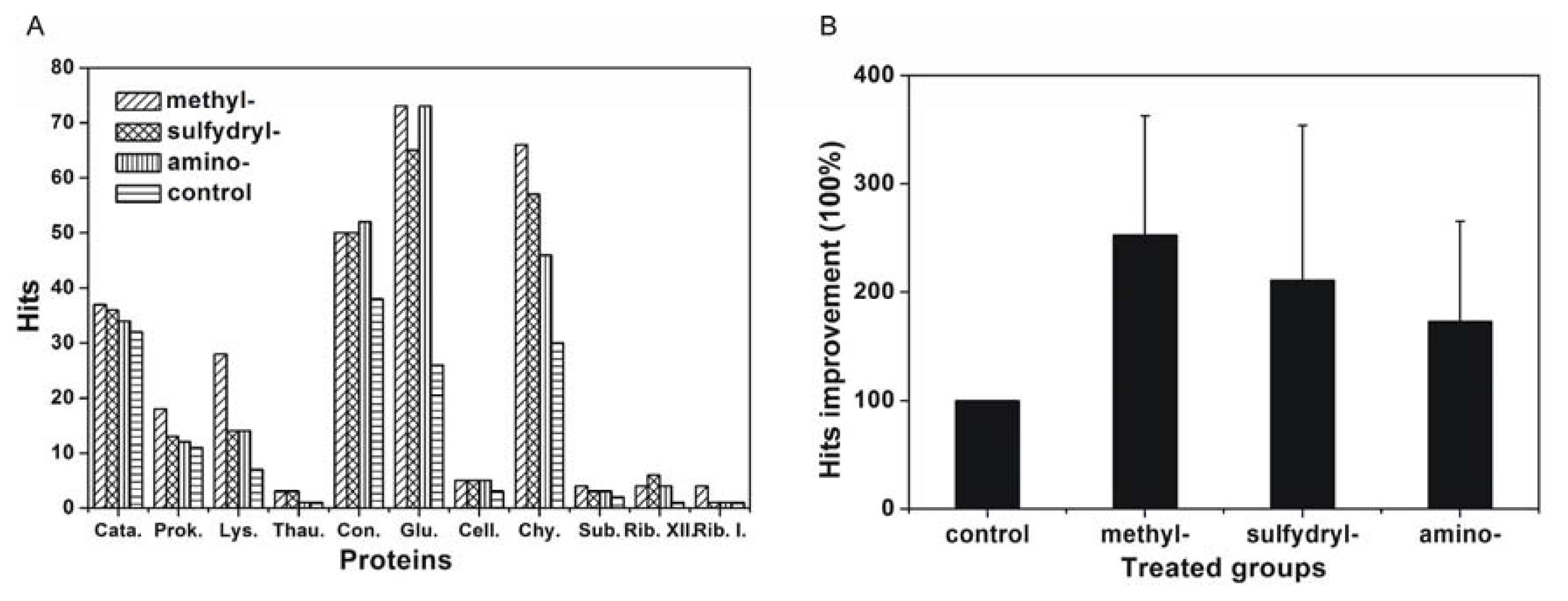
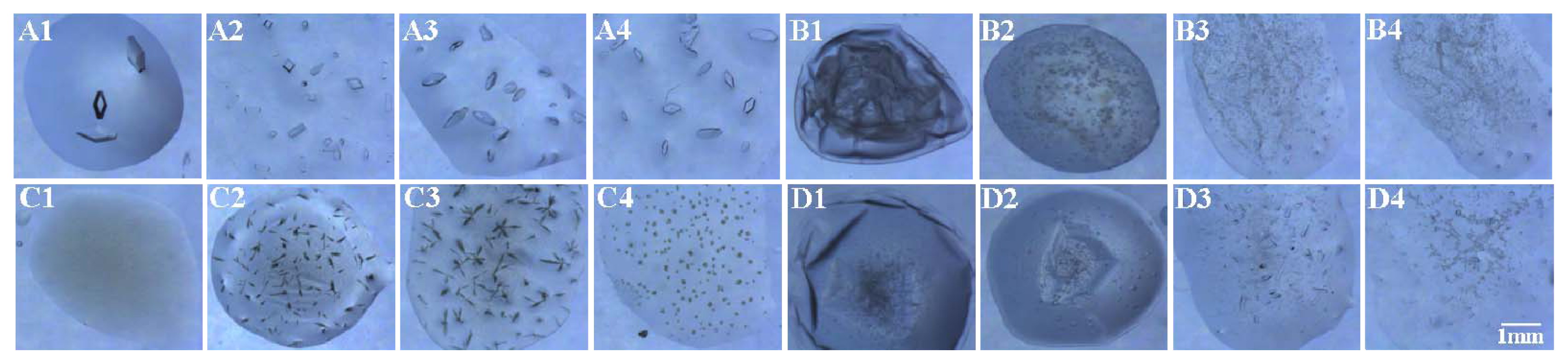
2.3. Effect of Modified SAMs on Protein Crystallisation Screening
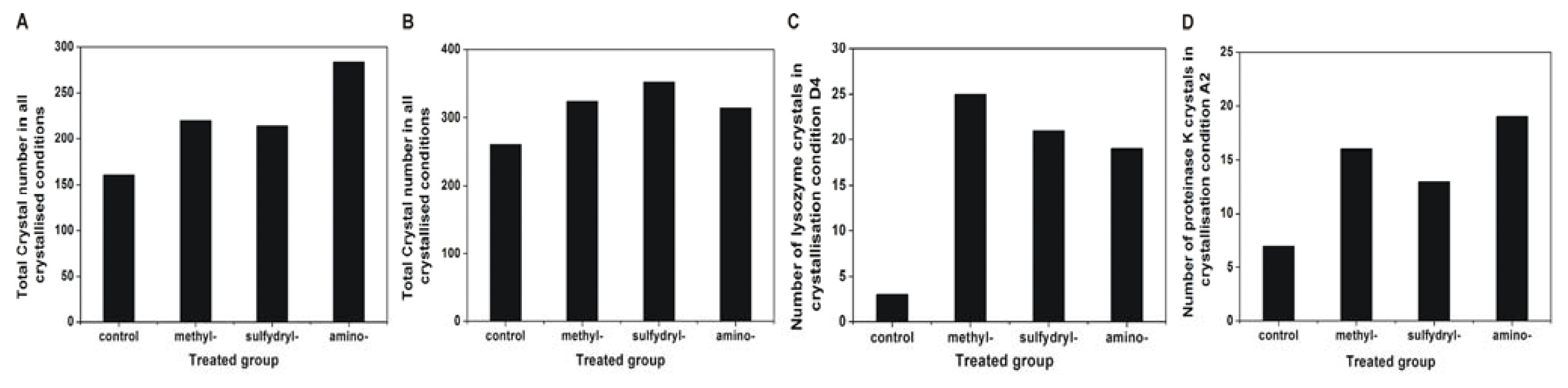
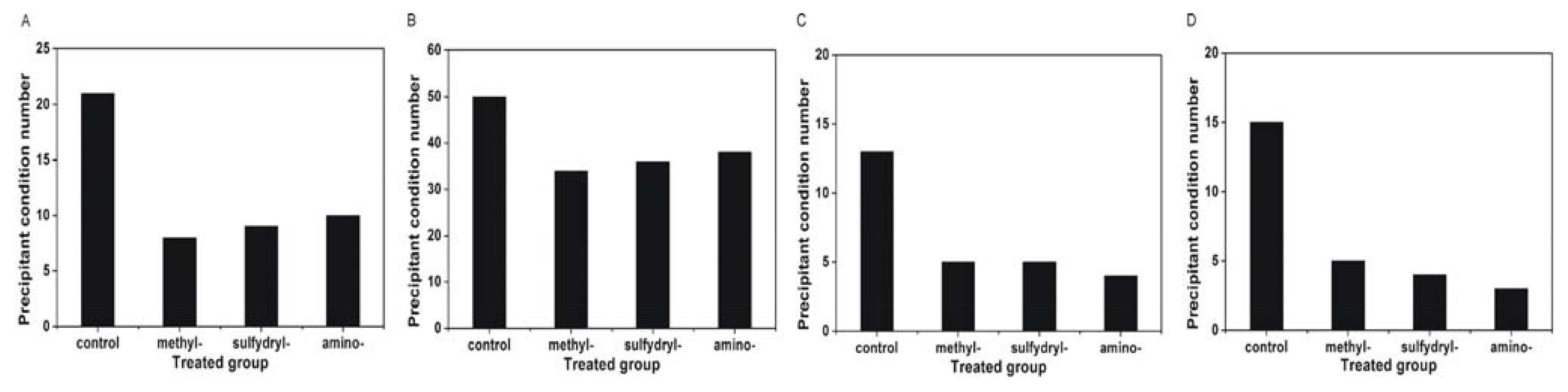
2.4. Effect of Modified SAMs on the Number of Crystals
2.5. Crystal Orientation on the Amino-Modified Cover Slip
3. Discussions
3.1. Increasing Opportunities of Heterogeneous Nucleation on Modified SAMs
3.2. Increasing the Fluctuation in Concentration on the Modified SAMs
3.3. Increase of the Fluctuation in Structure on Modified SAMs
4. Experimental Section
4.1. Materials
4.2. Experiments
4.2.1. Preparation of Modified SAMs and Silanised Cover Slips
4.2.2. Analysis of the SAMs
4.2.3. Crystallisation Experiments
5. Conclusions
Acknowledgments
Conflict of Interest
References
- Maruyama, Y.; Ebihara, T.; Nishiyama, H.; Konyuba, Y.; Senda, M.; Numaga-Tomita, T.; Senda, T.; Suga, M.; Sato, C. Direct observation of protein microcrystals in crystallization buffer by atmospheric scanning electron microscopy. Int. J. Mol. Sci 2012, 13, 10553–10567. [Google Scholar]
- Lu, H.M.; Yin, D.C.; Liu, Y.M.; Guo, W.H.; Zhou, R.B. Correlation between protein sequence similarity and crystallization reagents in the biological macromolecule crystallization database. Int. J. Mol. Sci 2012, 13, 9514–9526. [Google Scholar]
- Bernstein, F.C.; Koetzle, T.F.; Williams, G.J.; Meyer, E.F., Jr; Brice, M.D.; Rodgers, J.R.; Kennard, O.; Shimanouchi, T.; Tasumi, M. The Protein Data Bank: a computer-based archival file for macromolecular structures. J. Mol. Biol 1977, 112, 535–542. [Google Scholar]
- Schmit, J.D.; Dill, K. Growth rates of protein crystals. J. Am. Chem. Soc 2012, 134, 3934–3937. [Google Scholar]
- Garcia-Ruiz, J.M. Nucleation of protein crystals. J. Struct. Biol 2003, 142, 22–31. [Google Scholar]
- Chayen, N.E. Methods for separating nucleation and growth in protein crystallisation. Prog. Biophys. Mol. Biol 2005, 88, 329–337. [Google Scholar]
- Eslami, F.; Elliott, J.A. Thermodynamic investigation of the barrier for heterogeneous nucleation on a fluid surface in comparison with a rigid surface. J. Phys. Chem. B 2011, 115, 10646–10653. [Google Scholar]
- Chayen, N.E.; Saridakis, E.; El-Bahar, R.; Nemirovsky, Y. Porous silicon: An effective nucleation- crystallization. J. Mol. Biol 2001, 312, 591–595. [Google Scholar]
- Cacciuto, A.; Auer, S.; Frenkel, D. Onset of heterogeneous crystal nucleation in colloidal suspensions. Nature 2004, 428, 404–406. [Google Scholar]
- Thakur, A.S.; Robin, G.; Guncar, G.; Saunders, N.F.W.; Newman, J.; Martin, J.L.; Kobe, B. Improved success of sparse matrix protein crystallization screening with heterogeneous nucleating agents. PLoS One 2007, 2, e1091. [Google Scholar]
- McPherson, A.; Shlichta, P. Heterogeneous and epitaxial nucleation of protein crystals on mineral surfaces. Science 1988, 239, 385–387. [Google Scholar]
- Tang, L.; Huang, Y.B.; Liu, D.Q.; Li, J.L.; Mao, K.; Liu, L.; Cheng, Z.J.; Gong, W.M.; Hu, J.; He, J.H. Effects of the silanized mica surface on protein crystallization. Acta Crystallogr. D Biol. Crystallogr 2005, 61, 53–59. [Google Scholar]
- Hemming, S.A.; Bochkarev, A.; Darst, S.A.; Kornberg, R.D.; Ala, P.; Yang, D.S.; Edwards, A.M. The mechanism of protein crystal growth from lipid layers. J. Mol. Biol 1995, 246, 308–316. [Google Scholar]
- Wallace, E.; Dranow, D.; Laible, P.D.; Christensen, J.; Nollert, P. Monoolein lipid phases as incorporation and enrichment materials for membrane protein crystallization. PLoS One 2011, 6, e24488. [Google Scholar]
- Fermani, S.; Falini, G.; Minnucci, M.; Ripamonti, A. Protein crystallization on polymeric film surfaces. J. Cryst. Growth 2001, 224, 327–334. [Google Scholar]
- Liu, Y.X.; Wang, X.J.; Lu, J.; Ching, C.B. Influence of the roughness, topography, and physicochemical properties of chemically modified surfaces on the heterogeneous nucleation of protein crystals. J. Phys. Chem. B 2007, 111, 13971–13978. [Google Scholar]
- Guo, Y.Z.; Yin, D.C.; Lu, Q.Q.; Wang, X.K.; Liu, J. Enhancement of nucleation during hanging drop protein crystallization using HF treatment of cover glasses. Cryst. Res. Tech 2010, 45, 158–166. [Google Scholar]
- Ulman, A. Formation and structure of self-assembled monolayers. Chem. Rev 1996, 96, 1533–1554. [Google Scholar]
- Kidoaki, S.; Matsuda, T. Adhesion forces of the blood plasma proteins on self-assembled monolayer surfaces of alkanethiolates with different functional groups measured by an atomic force microscope. Langmuir 1999, 15, 7639–7646. [Google Scholar]
- Martins, M.C.; Ratner, B.D.; Barbosa, M.A. Protein adsorption on mixtures of hydroxyl- and methyl-terminated alkanethiols self-assembled monolayers. J. Biomed. Mater. Res. A 2003, 67, 158–171. [Google Scholar]
- Rixman, M.A.; Dean, D.; Macias, C.E.; Ortiz, C. Nanoscale intermolecular interactions between human serum albumin and alkanethiol self-assembled monolayers. Langmuir 2003, 19, 6202–6218. [Google Scholar]
- Sperling, C.; Schweiss, R.B.; Streller, U.; Werner, C. In vitro hemocompatibility of self-assembled monolayers displaying various functional groups. Biomaterials 2005, 26, 6547–6557. [Google Scholar]
- Barbosa, J.N.; Barbosa, M.A.; Aguas, A.P. Adhesion of human leukocytes to biomaterials: An in vitro study using alkanethiolate monolayers with different chemically functionalized surfaces. J. Biomed. Mater. Res. A 2003, 65, 429–434. [Google Scholar]
- Pham, T.; Lai, D.; Ji, D.; Tuntiwechapikul, W.; Friedman, J.M.; Lee, T.R. Well-ordered self-assembled monolayer surfaces can be used to enhance the growth of protein crystals. Colloids Surf. B Biointerfaces 2004, 34, 191–196. [Google Scholar]
- Ji, D.; Arnold, C.M.; Graupe, M.; Beadle, E.; Dunn, R.V.; Phan, M.N.; Villazana, R.J.; Benson, R.; Colorado, R., Jr; Lee, T.R.; et al. Improved protein crystallization by vapor diffusion from drops in contact with transparent, self-assembled monolayers on gold-coated glass coverslips. Growth 2000, 218, 390–398. [Google Scholar]
- Chayen, N.E.; Saridakis, E. Protein crystallization: From purified protein to diffraction-quality crystal. Nat. Methods 2008, 5, 147–153. [Google Scholar]
- Vekilov, P.G. Nucleation. Cryst. Growth Des 2010, 10, 5007–5019. [Google Scholar]
- De Yoreo, J.J.; Vekilov, P.G. Principles of Crystal Nucleation and Growth. In Biomineralization; Mineralogical Society of America: Washington, DC, USA, 2003; pp. 57–93. [Google Scholar]
- Yin, D.C.; Lu, H.M.; Geng, L.Q.; Shi, Z.H.; Luo, H.M.; Li, H.S.; Ye, Y.J.; Guo, W.H.; Shang, P.; Wakayama, N.I. Growing and dissolving protein crystals in a levitated and containerless droplet. J. Crystal Growth 2008, 310, 1206–1212. [Google Scholar]
- Steer, B.; Gorbunov, B.; Rowles, J.; Green, D. Surface area controlled heterogeneous nucleation. J. Chem. Phys 2012, 136, 054704. [Google Scholar]
- Borgstahl, G.E.; Vahedi-Faridi, A.; Lovelace, J.; Bellamy, H.D.; Snell, E.H. A test of macromolecular crystallization in microgravity: Large well ordered insulin crystals. Acta Crystallogr. D Biol. Crystallogr 2001, 57, 1204–1207. [Google Scholar]
- Xie, Y.; Liu, M.F.; Zhou, J. Molecular dynamics simulations of peptide adsorption on self-assembled monolayer. Appl. Surf. Sci 2012, 258, 8153–8159. [Google Scholar]
- Sivaraman, B.; Fears, K.P.; Latour, R.A. Investigation of the effects of surface chemistry and solution concentration on the conformation of adsorbed proteins using an improved circular dichroism method. Langmuir 2009, 25, 3050–3056. [Google Scholar]
- Fears, K.P.; Latour, R.A. Assessing the influence of adsorbed-state conformation on the bioactivity of adsorbed enzyme layers. Langmuir 2009, 25, 13926–13933. [Google Scholar]
- Bonanni, B.; Bizzarri, A.R.; Cannistraro, S. Optimized biorecognition of cytochrome c 551 and azurin immobilized on thiol-terminated monolayers assembled on Au(111) substrates. J. Phys. Chem. B 2006, 110, 14574–14580. [Google Scholar]
- Dierks, K.; Meyer, A.; Oberthür, D.; Rapp, G.; Einspahr, H.; Betzel, C. Efficient UV detection of protein crystals enabled by fluorescence excitation at wavelengths longer than 300 nm. Acta Crystallogr. F Struct. Biol. Cryst. Commun 2010, 66, 478–484. [Google Scholar]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | CA a | Si (%) | O (%) | C (%) | N (%) | S (%) | Cl (%) |
|---|---|---|---|---|---|---|---|
| untreated | 18.4 | 61.9 | 38.1 | - | - | - | - |
| silanised | 111.0 | 51.0 | 34.9 | 14.1 | - | - | 0.1 |
| methyl-modified | 104.0 | 45.1 | 33.2 | 21.8 | - | - | - |
| sulfydryl-modified | 49.1 | 58.0 | 35.2 | 6.8 | - | 0.1 | - |
| amino-modified | 41.9 | 57.7 | 34.7 | 6.4 | 1.2 | - | - |
| Protein | pI | Protein charge | Hits | Ratio of hits between methyl- and amino-modified SAMs | |
|---|---|---|---|---|---|
| methyl-modified SAMs | amino-modified SAMs | ||||
| lysozyme | 11.3 | positive | 28 | 14 | 0.50 |
| subtilisin A type VII | 9.4 | positive | 4 | 3 | 0.75 |
| thaumatin | 12.0 | positive | 3 | 1 | 0.33 |
| ribonuclease A type I | 9.6 | positive | 4 | 1 | 0.25 |
| ribonuclease A type XII | 9.6 | positive | 4 | 4 | 1.00 |
| α-chymotrypsinogen A type II | 9.0 | positive | 66 | 30 | 0.70 |
| proteinase K | 9.0 | positive | 18 | 12 | 0.67 |
| catalase | 5.0 | negative | 37 | 34 | 0.92 |
| concanavalin A type VI | 5.5 | negative | 50 | 52 | 1.04 |
| glucose Isomerase | 3.0 | negative | 73 | 73 | 1.00 |
| cellulase | 4.8 | negative | 5 | 5 | 1.00 |
| No. | Protein | Catalogue No. | Supplier | Buffer |
|---|---|---|---|---|
| 1 | lysozyme | E05801 | Seikagaku | 100 mM sodium acetate, pH 4.6 |
| 2 | catalase | C40 | Sigma-Aldrich | 25 mM HEPES sodium, pH 7.0 |
| 3 | subtilisin A type VII | P5380 | Sigma-Aldrich | 25 mM HEPES sodium, pH 7.0 |
| 4 | thaumatin | T7638 | Sigma-Aldrich | 25 mM HEPES sodium, pH 7.0 |
| 5 | concanavalin A type VI | L7647 | Sigma-Aldrich | 25 mM HEPES sodium, pH 7.0 |
| 6 | ribonuclease A type I | R4875 | Sigma-Aldrich | 25 mM HEPES sodium, pH 7.0 |
| 7 | ribonuclease A type XII | R5500 | Sigma-Aldrich | 25 mM HEPES sodium, pH 7.0 |
| 8 | α-chymotrypinoyen A type II | C4879 | Sigma-Aldrich | 25 mM HEPES sodium, pH 7.0 |
| 9 | proteinase K | P6556 | Sigma-Aldrich | 25 mM HEPES sodium, pH 7.0 |
| 10 | glucose isomerase | HR7-100 | Hampton Research | 25 mM HEPES sodium, pH 7.0 |
| 11 | cellulase | C0615 | Sigma-Aldrich | 25 mM HEPES sodium, pH 7.0 |
© 2013 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Zhang, C.-Y.; Shen, H.-F.; Wang, Q.-J.; Guo, Y.-Z.; He, J.; Cao, H.-L.; Liu, Y.-M.; Shang, P.; Yin, D.-C. An Investigation of the Effects of Self-Assembled Monolayers on Protein Crystallisation. Int. J. Mol. Sci. 2013, 14, 12329-12345. https://doi.org/10.3390/ijms140612329
Zhang C-Y, Shen H-F, Wang Q-J, Guo Y-Z, He J, Cao H-L, Liu Y-M, Shang P, Yin D-C. An Investigation of the Effects of Self-Assembled Monolayers on Protein Crystallisation. International Journal of Molecular Sciences. 2013; 14(6):12329-12345. https://doi.org/10.3390/ijms140612329
Chicago/Turabian StyleZhang, Chen-Yan, He-Fang Shen, Qian-Jin Wang, Yun-Zhu Guo, Jin He, Hui-Ling Cao, Yong-Ming Liu, Peng Shang, and Da-Chuan Yin. 2013. "An Investigation of the Effects of Self-Assembled Monolayers on Protein Crystallisation" International Journal of Molecular Sciences 14, no. 6: 12329-12345. https://doi.org/10.3390/ijms140612329