3.1. Genetic Diversity and Differentiation
C. japonicum possessed considerably low genetic diversity both at the species (
PPL = 38.53%,
H = 0.1273 and
I = 0.1928) and population levels (
PPL = 11.19%,
H = 0.0416 and
I = 0.0613) compared to other orchid species (average value:
PPLs = 64.36%,
Hs = 0.2788 and
Is = 0.3997;
PPLp = 50.51%,
Hp= 0.1812 and
Ip= 0.3084) using the same molecular makers (ISSR) (
Table 4). The high
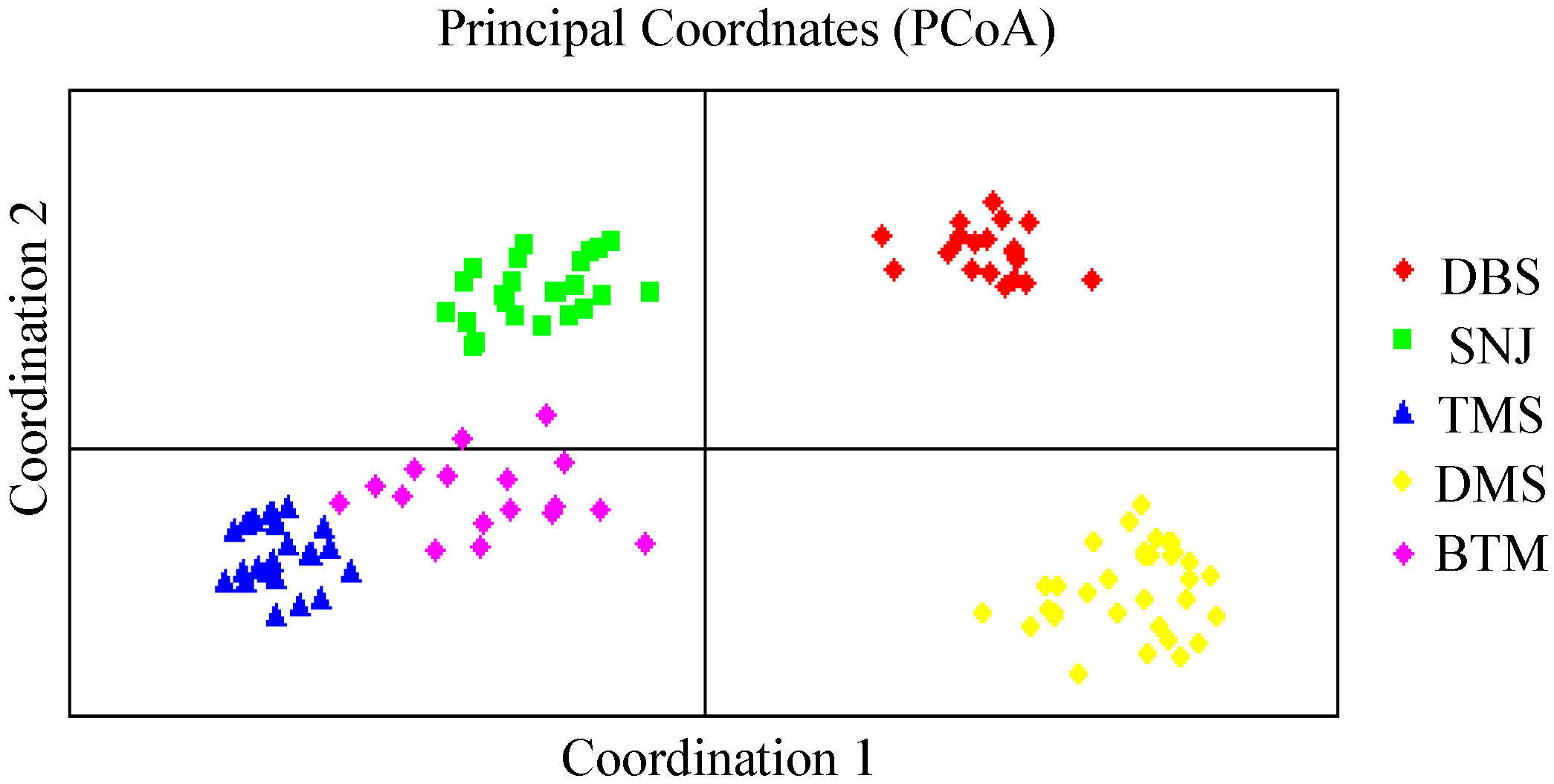
Gst values (0.6712) and the UPGMA and PCoA results suggested limited gene flow, even between the less distant populations (TMS and DMS populations), as well as marked differentiation between populations, which was supported by the AMOVA results (
Φpt = 0.698), indicating that the majority of genetic diversity was explained by variation among populations. The species’ ISSR genetic diversity of
C. japonicum was also lower than some other self-compatible plants (such as
Camellia japonica [
32],
Lilium tsingtauense [
33] and
Viola pubescens [
34]) and asexually reproducing plants (such as
Carex moorcroftii [
35],
Mentha cervina [
16] and
Psammochloa villosa [
36]).
Table 4.
Comparisons in genetic diversity and differentiation in orchid species based on inter-simple sequence repeat (ISSR) markers.
Table 4.
Comparisons in genetic diversity and differentiation in orchid species based on inter-simple sequence repeat (ISSR) markers.
| Orchid Species | Hp | Ip | PPLp (%) | Hs | Is | PPLs (%) | Gst (Fst) | SR |
|---|
| Amitostigma hemipilioides | 0.603 | 0.2949 | 50.9 | 0.686 | 0.3873 | 64.7 | 0.367 | [26] |
| Brassavola tuberculata | - | - | - | 0.216 | 0.314 | 52.41 | - | [37] |
| Calanthe tsoongiana | 0.183 | 0.271 | 50.0 | 0.398 | 0.576 | 96.80 | 0.55 | [27] |
| Cattleya bicolor | - | - | - | 0.219 | 0.323 | 56.63 | - | [37] |
| Cattleya elongata | - | - | 56.8 | - | - | - | 0.18 | [28] |
| Cattleya granulosa | - | - | - | 0.163 | 0.237 | 30.72 | - | [37] |
| Cattleya labiata | - | - | - | 0.132 | 0.193 | 30.12 | - | [37] |
| Cattleya schofieldiana | - | - | - | 0.213 | 0.314 | 56.02 | - | [37] |
| Cymbidium goeringii | 0.1945 | 0.2958 | 63.06 | 0.2628 | 0.4037 | 88.19 | 0.244 | [29] |
| Dendrobium fimbriatum | 0.0871 | 0.1290 | 23.93 | 0.3227 | 0.4779 | 89.74 | 0.7443 | [38] |
| Gastrodia elata | 0.176 | 0.270 | 59.09 | 0.236 | 0.367 | 81.82 | 0.2725 | [39] |
| Octomeria crassifolia | 0.2648 | 0.4005 | 91.57 | 0.352 | 0.530 | - | 0.76 | [40] |
| Octomeria grandiflora | 0.2578 | 0.382 | 82.4 | 0.338 | 0.508 | - | 0.12 | [40] |
| Paphiopedilum micranthum | 0.2847 | 0.4236 | 80.28 | 0.3839 | 0.5646 | 91.66 | 0.2577 | [41] |
| Piperia yadonii (2006) | 0.062 | - | - | - | - | - | 0.424 | [30] |
| Piperia yadonii (2007) | 0.059 | - | - | - | - | - | 0.394 | [30] |
| Platanthera aquilonis | 0.084 | - | 22.61 | 0.184 | - | 61.7 | 0.70 | [42] |
| Platanthera dilatata | 0.1312 | - | 35.35 | 0.182 | - | 57.5 | 0.49 | [42] |
| Platanthera huronensis | 0.119 | - | 32.64 | 0.172 | - | 43.0 | 0.36 | [42] |
| Tipularia discolor | 0.0309 | - | 7.95 | - | - | - | 0.415 | [43] |
| Average | 0.1812 | 0.3084 | 50.51 | 0.2788 | 0.3997 | 64.36 | 0.4186 | |
Much of our knowledge of genetic diversity and population structure in
Cypripedium orchids has come from allozyme markers. Chung
et al. [
14] summarized the allozyme diversity levels of 10
Cypripedium species and concluded that genetic diversity within the
Cypripedium genera can vary substantially, and those with traits, such as wide distribution and broad habitat preference, harbored much greater genetic diversity than rare species with narrower habitat amplitudes. Therefore, commonness and habitat diversity appear to be appropriate predictors for genetic variation in
Cypripedium. From an ecological perspective,
C. japonicum is a habitat specialist that is only found on rich humus soils under old, shady, wet deciduous forests on mountainous hillsides, partly due to the narrow mycorrhizal specificity for seed germination and seedling development [
44] under local microsite conditions with special soil characteristics.
The biological properties of species, such as the types of reproduction and breeding systems, are stressed as one of the most important factors shaping the genetic diversity and extent of plant population differentiation [
8]. Rewarding animal-pollinated species favors near-neighbor mating, which restricts pollen dispersal distances and increases genetic differentiation. [
29]. Conversely, a deceptive species generally disperses its pollen over long distances, which results in a high outcrossing rate [
1].
C. japonicum does not produce food rewards (nectar or edible pollen); rather, it has evolved a food-deceptive pollination system, like most other members of the
Cypripedium genus [
13].
C. japonicum probably attracts pollinators visiting the flowers through false nectar guides; The pollinator enters the labellum via a front orifice. To escape, the insect should squeezes out of the rear exit hole, forcing it to pass under the receptive stigma and then pick up a new mass of pollen. Suetsugu
et al. [
13] found that
Bombus ardens and
Bombus diversus diversus were the effective pollinators in the Japanese
C. japonicum population, whereas two bumblebee species,
Bombus remotus and
Bombus imitator, have been shown to be the effective primary pollinators of
C. japonicum in the SNJ population. However, the remarkable genetic differentiation among populations in studied
C. japonicum populations does not match what is expected, indicating that some other more important factors shape their current genetic pattern. The same situation was observed in
Changnienia amoena, another non-rewarding orchid [
1]. We hold that pollen transfer among the studied populations would be very difficult due to large geographic distance and a limited pollinator flying range. Jacquemyn
et al. [
44] suggested that nectar less pollination results in a fitness cost, with declining fruit set and seedling recruitment rates when the population size reduces. Sun
et al. [
11] found a very low fruit set (5.2%–7.7%) of
C. japonicum individuals under natural conditions. One reason for the low genetic diversity in
C. japonicum populations can also be explained by the less successful fruit production. The mobility of bumblebees and the highly self-compatibility of
C. japonicum [
13] did not exclude the possibility that they could transport pollinia between flowers within the same genet or between closely-related individuals, which could raise the proportion of recessive deleterious alleles.
Most orchid species have experienced acute declines in abundance due to a wide range of human activities that have resulted in habitat fragmentation, and the remaining populations tend to be small and isolated [
45]. Izawa
et al. [
46] pointed out that the habitat range and population size of Lady’s slipper have rapidly declined, especially since the 1950s, mainly because of illegal collecting for the purposes of horticultural use and sale. Small populations of
C. japonicum were also observed in severely fragmented habitats, which makes this species very vulnerable to habitat disturbance and climate change. Population genetics theory predicts that rare orchid species occurring in small, spatially isolated populations will lead to the random fixation of alleles within populations and subsequent low levels of genetic variation; these populations will exhibit a high degree of population genetic divergence, primarily as a result of stochastic events, genetic drift and inbreeding [
14]. Higher levels of genetic drift and inbreeding promote genetic erosion at both the population and species levels. Moreover, fragmented populations, owing to the long distances between
C. japonicum localities, may be exposed to reduced gene flow, which is also considered to be an important factor increasing genetic divergence between populations. The lack of correlation between geographic and genetic distances of the studied
C. japonicum, revealed by the Mantel test, UPGMA clustering and PCoA, might be also caused by population fragmentation, genetic drift and restricted gene flow.
Generally, gene flow between populations occurs by one of only two methods: (1) pollen movement by pollinators; and (2) seed dispersal by the wind. Considering the former method, we expect that pollen transfer by insects is not likely when there are large geographical distances between different populations. Long-distance gene flow by pollen is not possible given the limited flight ranges of pollinators. Although orchids are commonly known to show wind seed dispersal over a wide range, our field observations showed that the dense forest structure surrounding the
C. japonicum populations and the low fruit set likely reduced the probability of seeds being dispersed over long distances between populations. Limited seed dispersal distances have also been reported for other
Cypripedium orchids. For example, Chung
et al. [
14] stated that most
Cypripedium macranthos seeds fall close to the maternal plants on the basis of the significant fine-scale spatial genetic structure, and Brzosko
et al. [
47] confirmed that
Cypripedium calceolus seed dispersal occurred mostly at short distances within or around the fruiting ramets. Infrequent long-distance seed dispersal combined with a paucity of suitable habitats suggests that the natural founding of new
C. japonicum populations is a rare event. However, we cannot completely exclude great distance gene transfer via seeds through intensive tourism, forest management or some other random event. If long-distance seed dispersal occurs, founder effects are also likely to take place when populations are established by a small number of seed sources, resulting in a loss of genetic diversity during colonization [
48]. Therefore, we cannot rule out the possibility that the low levels of genetic diversity, at least in some of the studied populations, may be due to founder effects.
Much literature documenting the influence of the Last Glacial Maximum on population genetic structure describe high levels of genetic diversity in previously unglaciated ranges and relatively low genetic variation in formerly glaciated areas. The regions of sampled
C. japonicum in this study belong to the “Sino-Japanese Floristic Region”, which was not affected by the major quaternary glaciations [
49]. Therefore, the deficiency of genetic diversity in this orchid species contradicts what would be expected. The opposite pattern has also been found in two other congeneric orchids:
Cypripedium parviflorum [
50] and
Cypripedium reginae [
51]. Chung
et al. [
14] attributed the lack of genetic variation both within and among conspecific populations of
C. japonicum in South Korea to their origination from the same genetically depauperate glacial refugium. However, our data are not sufficient to assess the influence of the Last Glacial Maximum on
C. japonicum genetic diversity. Further studies involving a wider spectrum of populations from the geographic range of this orchid and the use of more genetic markers, including some codominant DNA markers, could help precisely determine how the glaciations shaped the genetic diversity in its distribution.
3.2. Conservation Implication
Knowledge about population genetic structure is considered an important element of conservation biology, so answers to these questions could help develop effective conservation management measures. The loss of genetic diversity will decrease adaptability to environmental changes, which reduces evolutionary potential. In addition, C. japonicum has been collected by plant sellers, herbalists and orchid hobbyists in recent decades, due to its horticultural and medicinal potential. This species has suffered from habitat loss and anthropogenic threats, including deforestation, building, agriculture and tourism. Many previously recorded populations have disappeared. Consequently, C. japonicum is at risk of extinction in China. Our genetic data and field observations suggest that the main explanatory factors for the low levels of genetic diversity and the shaping of the population genetic structure of C. japonicum are narrow habitat preference, genetic drift due to a small and isolated population size, restricted gene flow, historical events and human activity. Considering the current genetic diversity and structure of this orchid species, several conservation guidelines are suggested for the studied C. japonicum population.
We recommend that all of the studied populations should be incorporated into conservation plans to maintain their total genetic diversity and, thus, prevent a further decline in the effective population size. Because each individual population comprises a unique genetic pool given the high genetic differentiation among the studied
C. japonicum populations, the disappearance of each population would remove any unique biological characteristics that it may possess, and this could reduce the overall species biodiversity. Protection of habitats of standing populations
in situ and prohibition of
C. japonicum by law should be stressed to prevent habitat damage and harvest pressure on wild populations. Because the fruit set of
C. japonicum in natural habitats is very low, nearby pollinator (bumblebees) populations and other plant species with floral rewards, if any, should also be safeguarded to ensure reproductive success. Furthermore, the method of artificial gene flow via seeds and hand-mediated artificial pollination suggested for
Cypripedium macranthos var.
rebunense [
46] may be also fit for
C. japonicum to rapidly increase the gene flow and relieve the effects of genetic drift. However, careful attention should be paid when implementing artificial gene flow. Concerning
ex situ conservation measures, seed asymbiotic germination and tissue cultures would be helpful for the success of further reinforcement or reintroduction. Sampling for
ex situ conservation should be taken from as many genets as possible in each population considering the current genetic structure of the studied
C. japonicum.
To design and implement comprehensive and effective conservation actions for this threatened orchid species, more research is needed to investigate life-history traits, demographic dynamics, germination biology, seedling establishment and other population ecology parameters. In addition, as C. japonicum spreads by long, creeping rhizomes, we cannot rule out the possibility that vegetative reproduction could also have a pronounced effect on the genetic structure of populations. Therefore, further study of clonal diversity and structure is also necessary in this endangered orchid.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}






