Overexpression of the Vitronectin V10 Subunit in Patients with Nonalcoholic Steatohepatitis: Implications for Noninvasive Diagnosis of NASH

 ,
, 
 , ,
, ,  and
and
Abstract
:1. Introduction
2. Results
2.1. Characteristics of the Patients
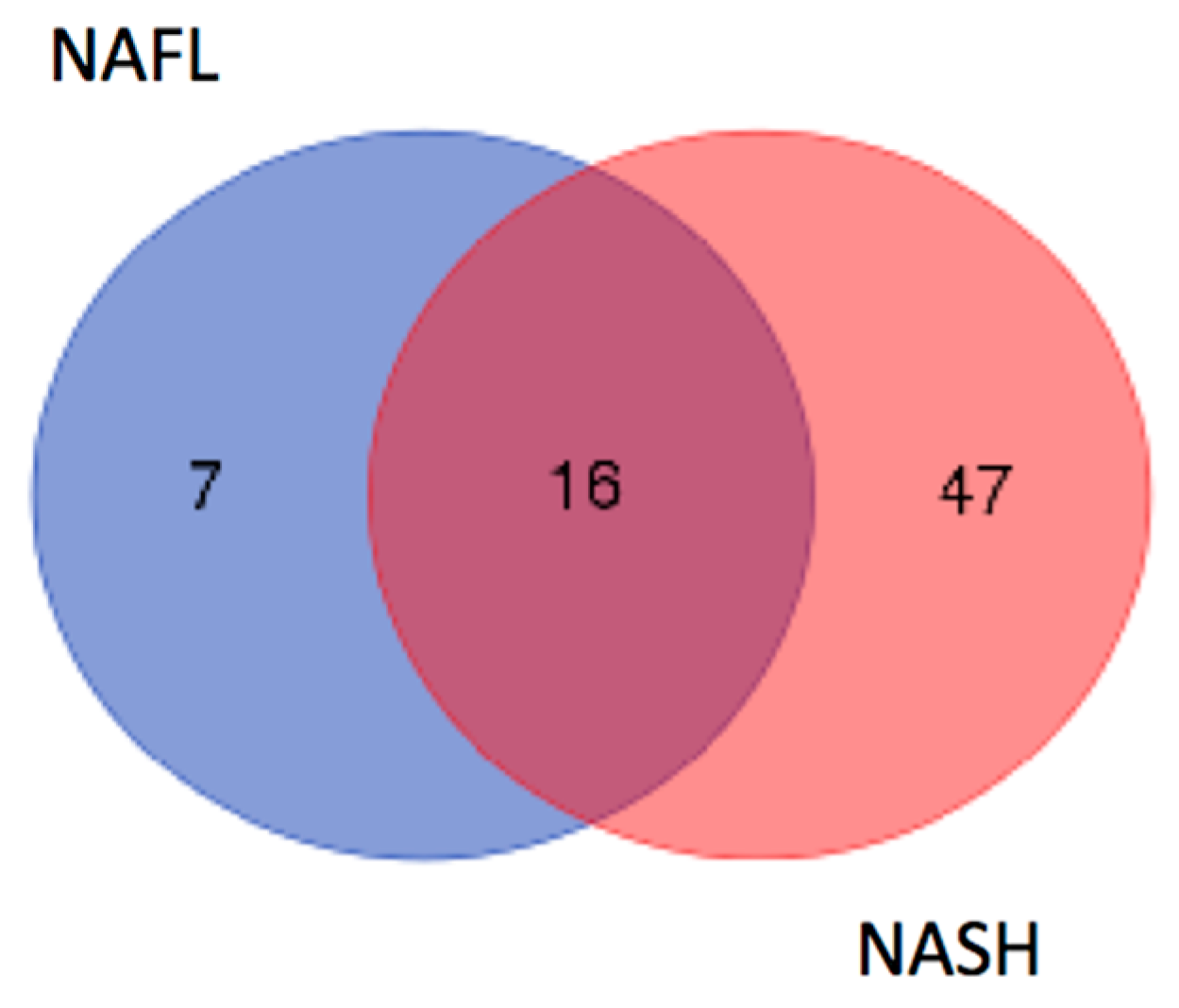
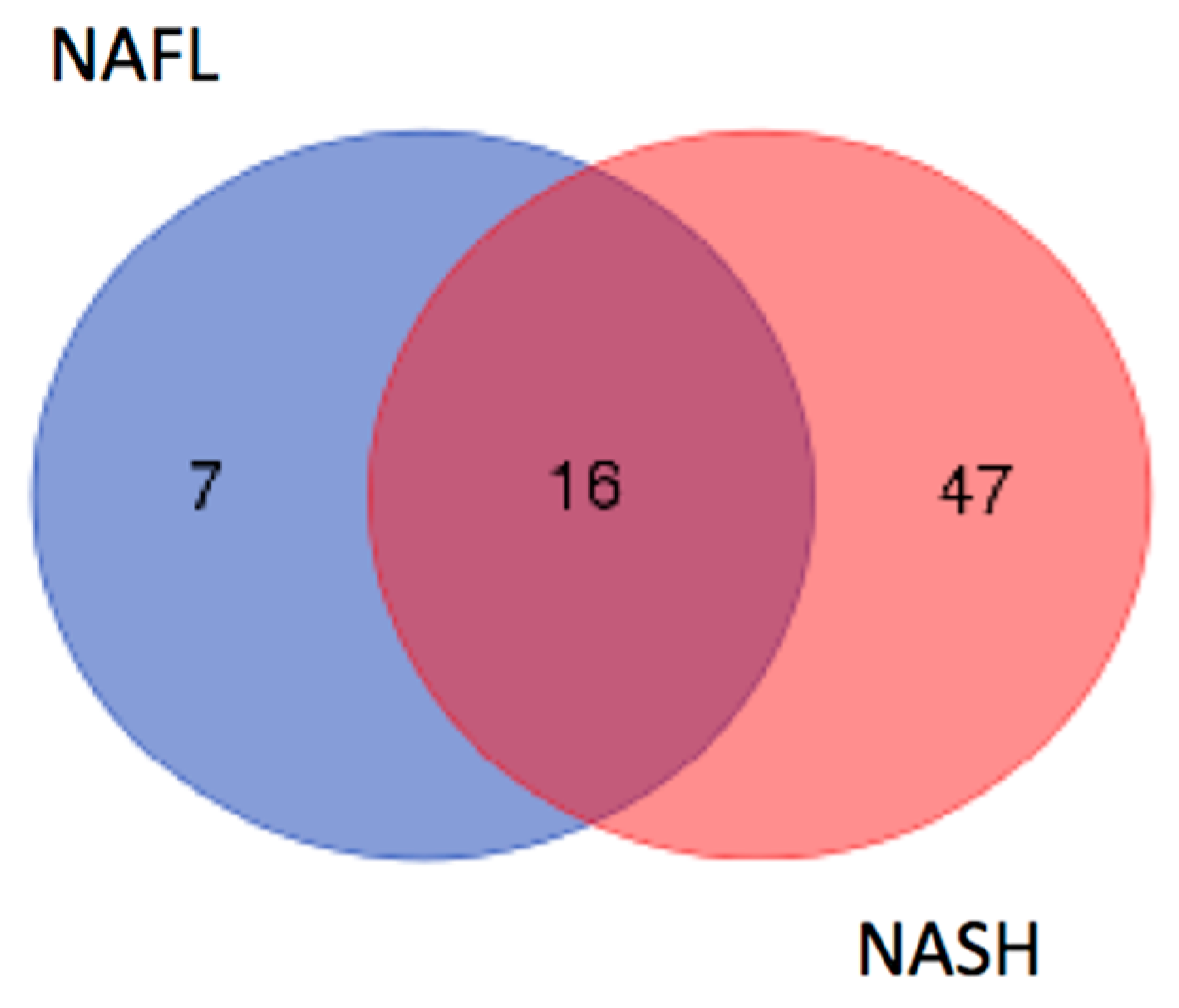
2.2. Identification of LMW Polypeptides in NAFL and NASH Samples
2.3. In Silico Analysis of NASH-Specific Fragments
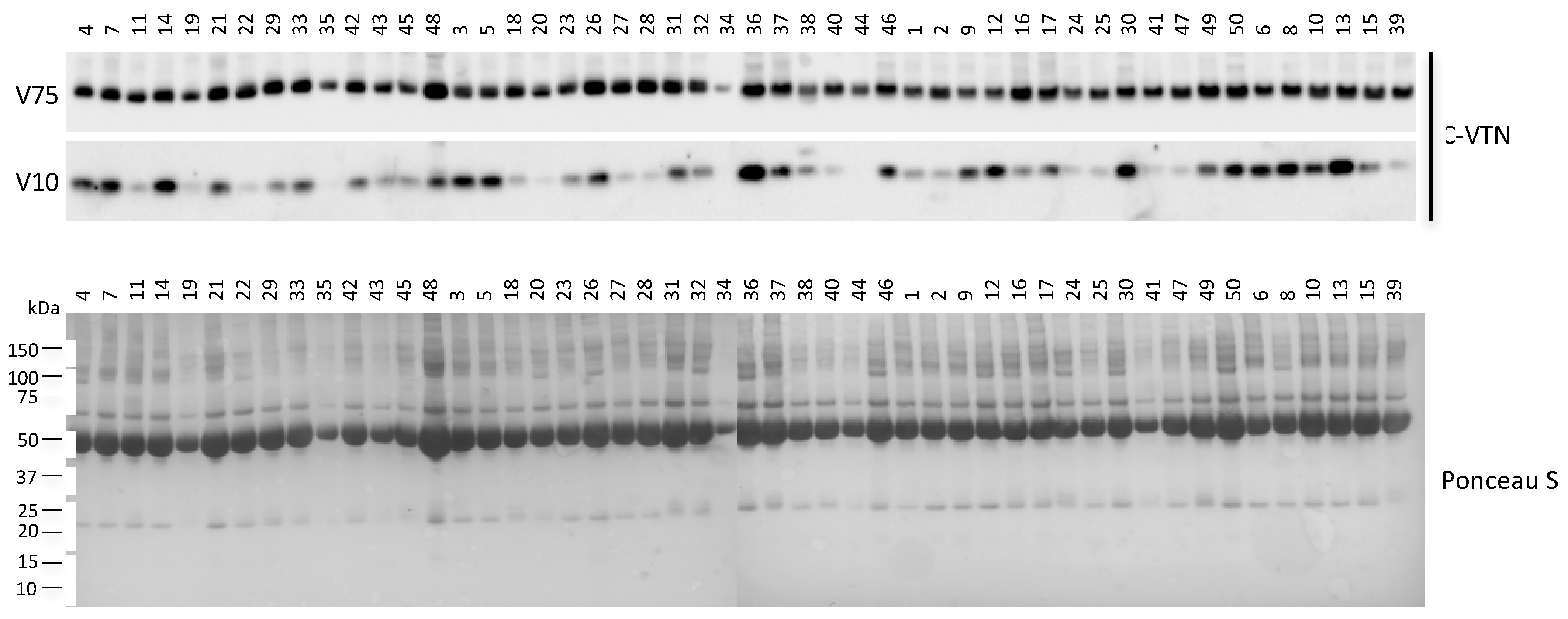
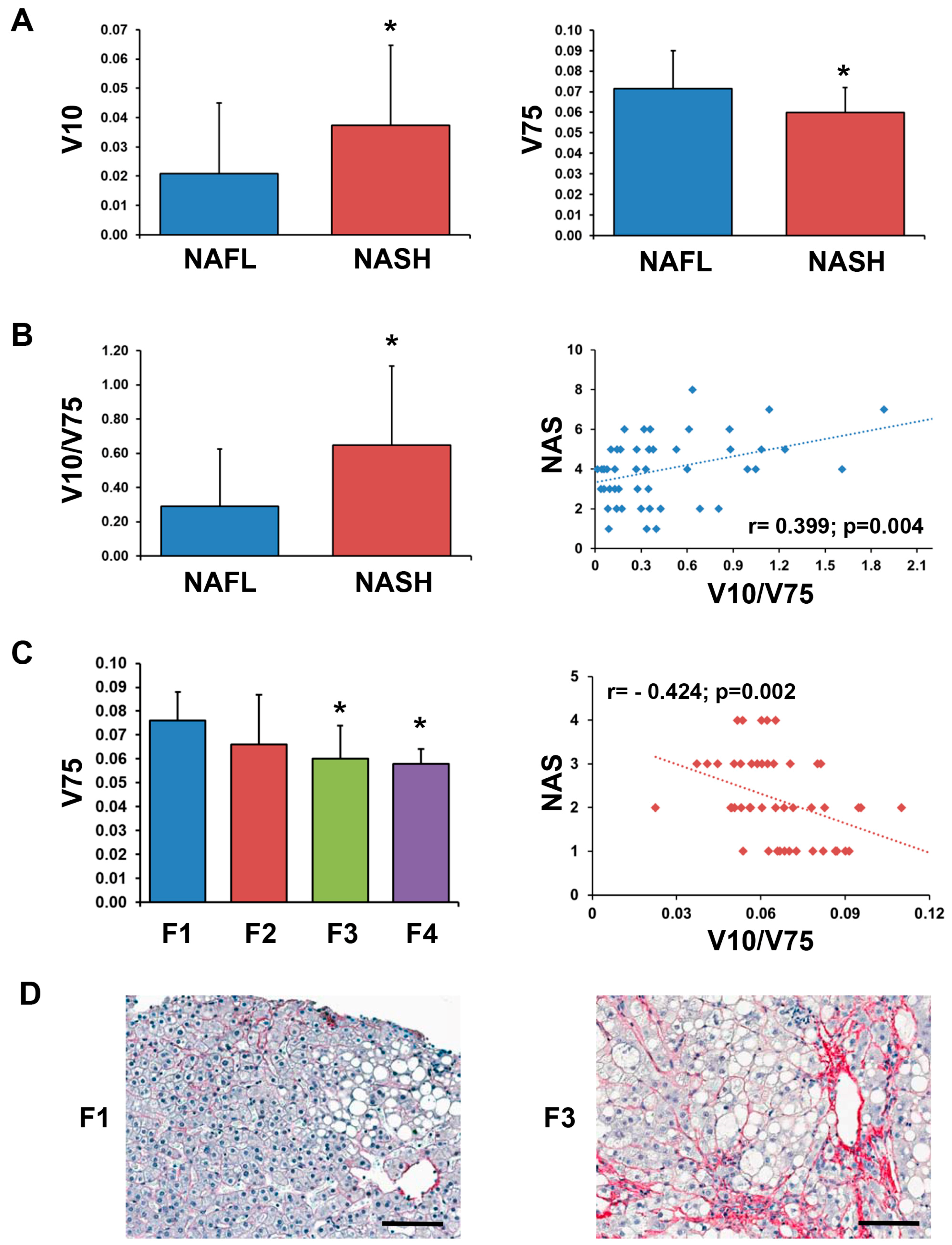
2.4. Correlation between Circulating VTN Fragments and Liver HistoMorphology
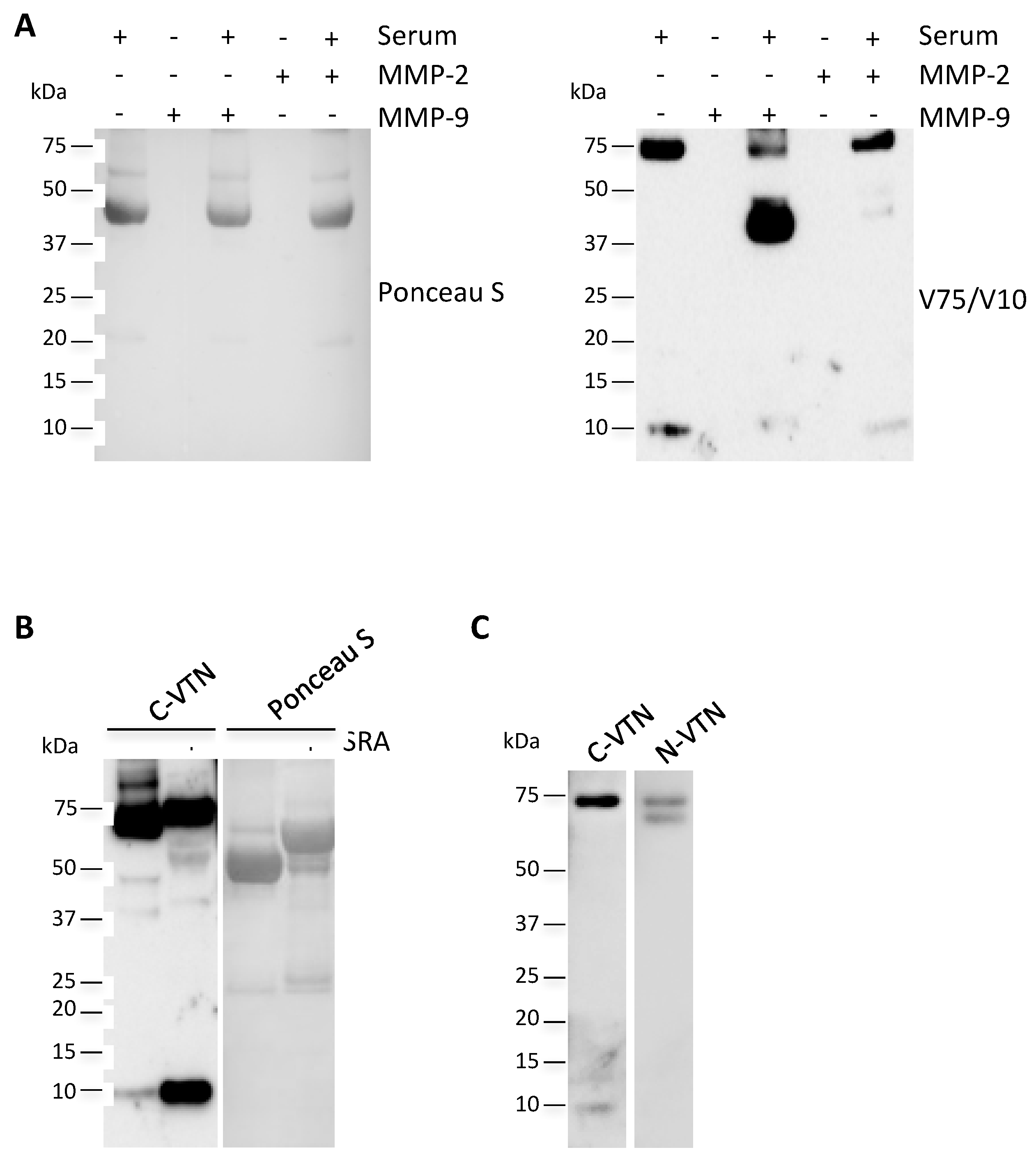
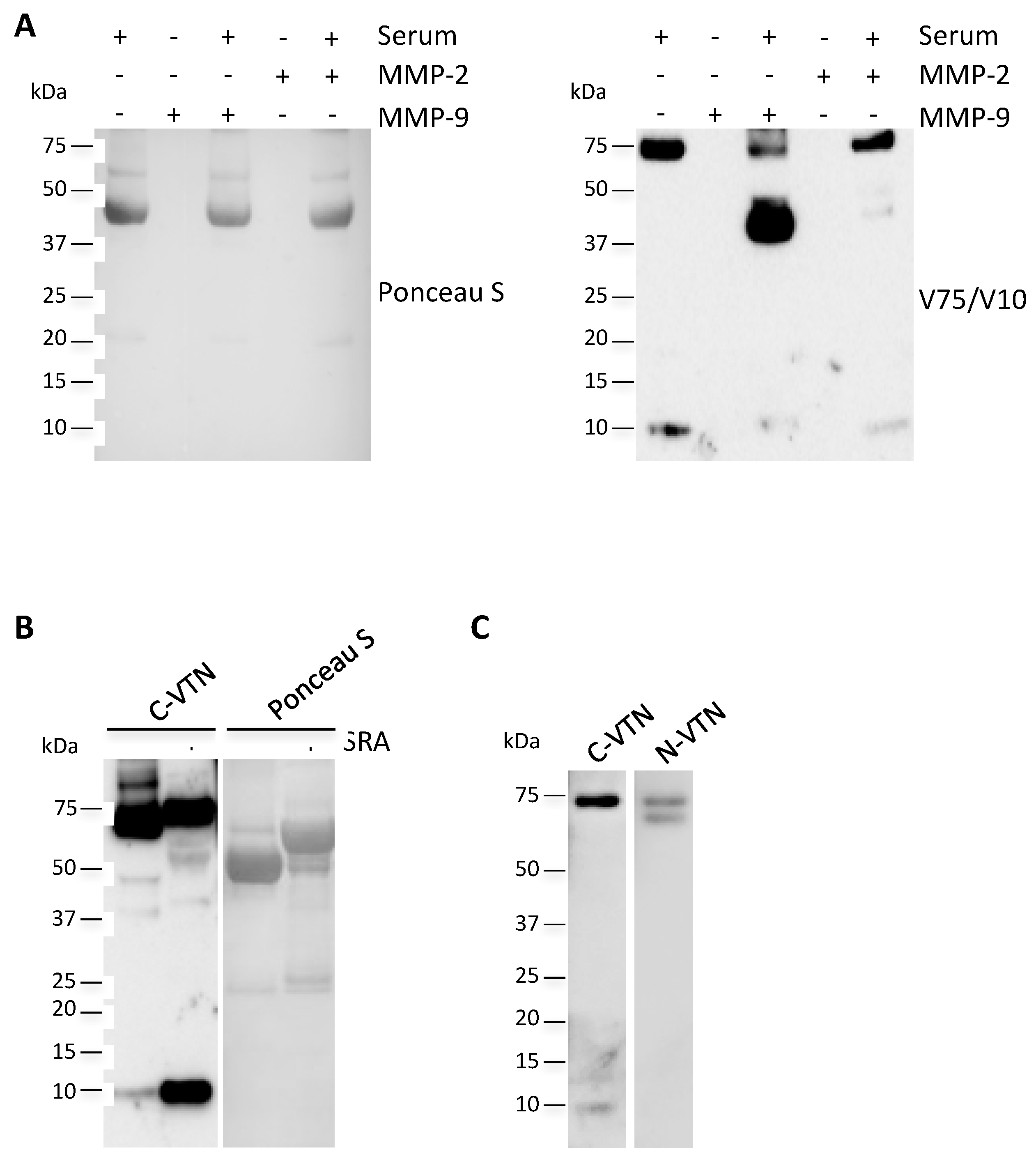
2.5. Circulating V10 Originates by the Reduction of VTN Clipped Form
3. Discussion
4. Materials and Methods
4.1. Patient Characteristics
4.2. Liver Secretome, Protein Digestion, and Peptide Purification
4.3. NanoLC Analysis and Mass Spectrometry Analysis
4.4. Immunoblotting Analysis
4.5. In Vitro Degradation of Vitronectin by Metalloproteases
4.6. Statistics
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Vernon, G.; Baranova, A.; Younossi, Z.M. Systematic review: The epidemiology and natural history of non-alcoholic fatty liver disease and non-alcoholic steatohepatitis in adults. Aliment Pharmacol. Ther. 2011, 34, 274–285. [Google Scholar] [CrossRef] [PubMed]
- Bedossa, P. Pathology of non-alcoholic fatty liver disease. Liver Int. 2017, 37, 85–89. [Google Scholar] [CrossRef] [PubMed]
- Dam-Larsen, S.; Franzmann, M.; Andersen, I.B.; Christoffersen, P.; Jensen, L.B.; Sørensen, T.I.; Becker, U.; Bendtsen, F. Long term prognosis of fatty liver: Risk of chronic liver disease and death. Gut 2004, 53, 750–755. [Google Scholar] [CrossRef] [PubMed]
- Chalasani, N.; Younossi, Z.; Lavine, J.E.; Diehl, A.M.; Brunt, E.M.; Cusi, K.; Charlton, M.; Sanyal, A.J. The diagnosis and management of non-alcoholic fatty liver disease: Practice Guideline by the American Association for the Study of Liver Diseases, American College of Gastroenterology, and the American Gastroenterological Association. Hepatology 2012, 55, 2005–2023. [Google Scholar] [CrossRef] [PubMed]
- Kleiner, D.E.; Brunt, E.M.; van Natta, M.; Behling, C.; Contos, M.J.; Cummings, O.W.; Ferrell, L.D.; Liu, Y.C.; Torbenson, M.S.; Unalp-Arida, A.; et al. Design and validation of a histological scoring system for nonalcoholic fatty liver disease. Hepatology 2005, 41, 1313–1321. [Google Scholar] [CrossRef] [PubMed]
- Sanyal, A.J.; Brunt, E.M.; Kleiner, D.E.; Kowdley, K.V.; Chalasani, N.; Lavine, J.E.; Ratziu, V.; McCullough, A. Endpoints and clinical trial design for nonalcoholic steatohepatitis. Hepatology 2011, 54, 344–353. [Google Scholar] [CrossRef] [PubMed]
- Ratziu, V.; Charlotte, F.; Heurtier, A.; Gombert, S.; Giral, P.; Bruckert, E.; Grimaldi, A.; Capron, F.; Poynard, T.; LIDO Study Group. Sampling variability of liver biopsy in nonalcoholic fatty liver disease. Gastroenterology 2005, 128, 1898–1906. [Google Scholar] [CrossRef] [PubMed]
- Feldstein, A.E.; Wieckowska, A.; Lopez, A.R.; Liu, Y.C.; Zein, N.N.; McCullough, A.J. Cytokeratin-18 fragment levels as noninvasive biomarkers for nonalcoholic steatohepatitis: A multicenter validation study. Hepatology 2009, 50, 1072–1078. [Google Scholar] [CrossRef] [PubMed]
- Goh, G.B.; Issa, D.; Lopez, R.; Dasarathy, S.; Sargent, R.; Hawkins, C.; Pai, R.K.; Yerian, L.; Khiyami, A.; Pagadala, M.R.; et al. The development of a non-invasive model to predict the presence of non-alcoholic steatohepatitis in patients with non-alcoholic fatty liver disease. J Gastroenterol. Hepatol. 2016, 31, 995–1000. [Google Scholar] [CrossRef] [PubMed]
- Dong, S.; Zhan, Z.Y.; Cao, H.Y.; Wu, C.; Bian, Y.Q.; Li, J.Y.; Cheng, G.H.; Liu, P.; Sun, M.Y. Urinary metabolomics analysis identifies key biomarkers of different stages of nonalcoholic fatty liver disease. World J. Gastroenterol. 2017, 23, 2771–2784. [Google Scholar] [CrossRef]
- Yilmaz, Y.; Eren, F.; Ayyildia, T.; Colak, Y.; Kurt, R.; Senates, E.; Tuncer, I.; Dolar, E.; Imeryuz, N. Serum pigment epithelium-derived factor levels are increased in patients with biopsy-proven nonalcoholic fatty liver disease and independently associated with liver steatosis. Clin. Chim. Acta 2011, 412, 2296–2299. [Google Scholar] [CrossRef]
- Drake, R.R.; Cazares, L.; Semmes, O.J. Mining the low molecular weight proteome of blood. Proteomics Clin. Appl. 2007, 1, 758–768. [Google Scholar] [CrossRef] [PubMed]
- Karsdal, M.A.; Manon-Jensen, T.; Genovese, F.; Kristensen, J.H.; Nielsen, M.J.; Sand, J.M.; Hansen, N.U.; Bay-Jensen, AC.; Bager, C.L.; Krag, A.; et al. Novel insights into the function and dynamics of extracellular matrix in liver fibrosis. Am. J. Physiol. Gastrointest. Liver Physiol. 2015, 308, G807–G830. [Google Scholar] [CrossRef] [PubMed]
- Naba, A.; Clauser, K.R.; Ding, H.; Whittaker, C.A.; Carr, S.A.; Hynes, R.O. The Extracellular Matrix: Tools and Insights for the "Omics" Era. Matrix Biol. 2016, 49, 10–24. [Google Scholar] [CrossRef] [PubMed]
- Koukoulis, G.K.; Shen, J.; Virtanen, I.; Gould, V.E. Vitronectin in the cirrhotic liver: An immunomarker of mature fibrosis. Hum. Pathol. 2001, 32, 1356–1362. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, J.; Yamada, S.; Kawasaki, H. Distribution of vitronectin in plasma and liver tissue: Relationship to chronic liver disease. Hepatology 1994, 20, 1412–1417. [Google Scholar] [CrossRef] [PubMed]
- Inuzuka, S.; Ueno, T.; Torimura, T.; Tamaki, S.; Sakata, R.; Sata, M.; Yoshida, H.; Tanikawa, K. Vitronectin in liver disorders: Biochemical and immunohistochemical studies. Hepatology 1992, 15, 629–636. [Google Scholar] [CrossRef] [PubMed]
- Baiocchini, A.; Montaldo, C.; Conigliaro, A.; Grimaldi, A.; Correani, V.; Mura, F.; Ciccosanti, F.; Rotiroti, N.; Brenna, A.; Montalbano, M.; et al. Extracellular Matrix Molecular Remodeling in Human Liver Fibrosis Evolution. PLoS ONE 2016, 11, e0151736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schvartz, I.; Seger, D.; Shaltiel, S. Vitronectin. Int. J. Biochem. Cell. Biol. 1999, 31, 539–544. [Google Scholar] [CrossRef]
- Imai, K.; Shikata, H.; Okada, Y. Degradation of vitronectin by matrix metalloproteinases-1, -2, -3, -7 and -9. FEBS Lett. 1995, 369, 249–251. [Google Scholar] [CrossRef]
- Ljumovic, D.; Diamantis, I.; Alegakis, A.K.; Kouroumalis, E.A. Differential expression of matrix metalloproteinases in viral and non-viral chronic liver diseases. Clin. Chim. Acta 2004, 349, 203–211. [Google Scholar] [CrossRef] [PubMed]
- Finoulst, I.; Pinkse, M.; van Dongen, W.; Verhaert, P. Sample preparation techniques for the untargeted LC-MS-based discovery of peptides in complex biological matrices. J. Biomed. Biotechnol. 2011, 2011, 245291. [Google Scholar] [CrossRef] [PubMed]
- Anderson, N.L.; Anderson, N.G. The human plasma proteome: History, character, and diagnostic prospects. Mol. Cell. Proteom. 2002, 1, 845–867. [Google Scholar] [CrossRef] [PubMed]
- Kawashima, Y.; Fukutomi, T.; Tomonaga, T.; Takahashi, H.; Nomura, F.; Maeda, T.; Kodera, Y. High-yield peptide-extraction method for the discovery of subnanomolar biomarkers from small serum samples. J. Proteome Res. 2010, 9, 1694–1705. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Zhai, L.; Li, Y.; Li, N.; Zhang, C.; Ping, L.; Chang, L.; Wu, J.; Li, X.; Shi, D.; Xu, P. Development of gel-filter method for high enrichment of low-molecular weight proteins from serum. PLoS ONE 2015, 10, e0115862. [Google Scholar] [CrossRef] [PubMed]
- Paradis, V.; Degos, F.; Dargère, D.; Pham, N.; Belghiti, J.; Degott, C.; Janeau, J.L.; Bezeaud, A.; Delforge, D.; Cubizolles, M.; et al. Identification of a new marker of hepatocellular carcinoma by serum protein profiling of patients with chronic liver diseases. Hepatology 2005, 41, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Tariq, Z.; Green, C.J.; Hodson, L. Are oxidative stress mechanisms the common denominator in the progression from hepatic steatosis towards non-alcoholic steatohepatitis (NASH)? Liver Int. 2014, 34, e180–e190; [Google Scholar] [CrossRef] [PubMed]
- Polimeni, L.; del Ben, M.; Baratta, F.; Perri, L.; Albanese, F.; Pastori, D.; Violi, F.; Angelico, F. Oxidative stress: New insights on the association of non-alcoholic fatty liver disease and atherosclerosis. World J. Hepatol. 2015, 7, 1325–1336. [Google Scholar] [CrossRef] [PubMed]
- Del Ben, M.; Polimeni, L.; Baratta, F.; Bartimoccia, S.; Carnevale, R.; Loffredo, L.; Pignatelli, P.; Violi, F.; Angelico, F. Serum Cytokeratin-18 Is Associated with NOX2-Generated Oxidative Stress in Patients with Nonalcoholic Fatty Liver. Int. J. Hepatol. 2014, 2014, 784985. [Google Scholar] [CrossRef] [PubMed]
- Rolo, A.P.; Teodoro, J.S.; Palmeira, C.M. Role of oxidative stress in the pathogenesis of nonalcoholic steatohepatitis. Free Radic. Biol. Med. 2012, 52, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.S.; Buck, M.; Houglum, K.; Chojkier, M. Activation of hepatic stellate cells by TGFα and collagen type I is mediated by oxidative stress through c-myb expression. J. Clin. Invest. 1995, 96, 2461–2468. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Kim, S.M.; Lee, R.T. Thioredoxin and thioredoxin target proteins: From molecular mechanisms to functional significance. Antioxid. Redox Signal. 2013, 18, 1165–1207. [Google Scholar] [CrossRef] [PubMed]
- Sumida, Y.; Nakashima, T.; Yoh, T.; Furutani, M.; Hirohama, A.; Kakisaka, Y.; Nakajima, Y.; Ishikawa, H.; Mitsuyoshi, H.; Okanoue, T.; et al. Serum thioredoxin levels as a predictor of steatohepatitis in patients with nonalcoholic fatty liver disease. J. Hepatol. 2003, 38, 32–38. [Google Scholar] [CrossRef]
- Edwards, S.; Lalor, P.F.; Tuncer, C.; Adams, D.H. Vitronectin in human hepatic tumours contributes to the recruitment of lymphocytes in an alpha v beta3-independent manner. Br. J. Cancer 2006, 95, 1545–1554. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Lloyd, D.J.; Hale, C.; Stanislaus, S.; Chen, M.; Sivits, G.; Vonderfecht, S.; Hecht, R.; Li, Y.S.; Lindberg, R.A.; et al. Fibroblast growth factor 21 reverses hepatic steatosis, increases energy expenditure, and improves insulin sensitivity in diet-induced obese mice. Diabetes 2009, 58, 250–259. [Google Scholar] [CrossRef] [PubMed]
- Petrasek, J.; Bala, S.; Csak, T.; Lippai, D.; Kodys, K.; Menashy, V.; Barrieau, M.; Min, S.Y.; Kurt-Jones, E.A.; Szabo, G. IL-1 receptor antagonist ameliorates inflammasome-dependent alcoholic steatohepatitis in mice. J. Clin. Invest. 2012, 122, 3476–3489. [Google Scholar] [CrossRef] [PubMed]
- An, J.J.; Han, D.H.; Kim, D.M.; Kim, S.H.; Rhee, Y.; Lee, E.J.; Lim, S.K. Expression and regulation of osteoprotegerin in adipose tissue. Yonsei Med. J. 2007, 48, 765–772. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Xu, D.; Liu, Y.; Guo, X.; Li, W.; Guo, C.; Zhang, H.; Gao, Y.; Mao, Y.; Zhao, J. Combined Serum Biomarkers in Non-Invasive Diagnosis of Non-Alcoholic Steatohepatitis. PLoS ONE 2015, 10, e0131664. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NAFL | NASH | p Value | |
|---|---|---|---|
| All | 27 | 23 | |
| Gender (male/female) | 14/13 | 10/13 | 0.55 |
| Nas | 2.81 ± 1.04 | 5.43 ± 0.99 | <0.001 |
| Fibrosis 1/2/3/4 | 11/12/4/0 | 3/5/9/6 | <0.001 |
| AGE at enrolment (years) | 52.3 ± 14.4 | 51.7 ± .9.8 | 0.44 |
| Body mass index (kg/m2) | 28.7 ± 4.1 | 30.5 ± 3.9 | 0.067 |
| AST (U/L) | 37.6 ± 17.8 | 61.9 ± 40.9 | 0.005 |
| ALT (U/L) | 69.3 ± 36.1 | 97.1 ± 56.1 | 0.024 |
| GGT (U/L) | 62.4 ± 41.5 | 80.7 ± 86.7 | 0.18 |
| AST/ALT ratio | 0.56 ± 0.18 | 0.68 ± 0.31 | 0.07 |
| Gene Symbol | Name | Matrisome Division | Category |
|---|---|---|---|
| FGA | Fibrinogen alpha chain | Core matrisome | ECM Glycoproteins |
| VTN | Vitronectin | Core matrisome | ECM Glycoproteins |
| LGALS3 | Galectin-3 | Matrisome-associated | ECM-affiliated Proteins |
| LGALS4 | Galectin-4 | Matrisome-associated | ECM-affiliated Proteins |
| CTSB | Cathepsin B | Matrisome-associated | ECM Regulators |
| SERPINB1 | Leukocyte elastase inhibitor | Matrisome-associated | ECM Regulators |
| SERPINC1 | Antithrombin-III | Matrisome-associated | ECM Regulators |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Del Ben, M.; Overi, D.; Polimeni, L.; Carpino, G.; Labbadia, G.; Baratta, F.; Pastori, D.; Noce, V.; Gaudio, E.; Angelico, F.; et al. Overexpression of the Vitronectin V10 Subunit in Patients with Nonalcoholic Steatohepatitis: Implications for Noninvasive Diagnosis of NASH. Int. J. Mol. Sci. 2018, 19, 603. https://doi.org/10.3390/ijms19020603
Del Ben M, Overi D, Polimeni L, Carpino G, Labbadia G, Baratta F, Pastori D, Noce V, Gaudio E, Angelico F, et al. Overexpression of the Vitronectin V10 Subunit in Patients with Nonalcoholic Steatohepatitis: Implications for Noninvasive Diagnosis of NASH. International Journal of Molecular Sciences. 2018; 19(2):603. https://doi.org/10.3390/ijms19020603
Chicago/Turabian StyleDel Ben, Maria, Diletta Overi, Licia Polimeni, Guido Carpino, Giancarlo Labbadia, Francesco Baratta, Daniele Pastori, Valeria Noce, Eugenio Gaudio, Francesco Angelico, and et al. 2018. "Overexpression of the Vitronectin V10 Subunit in Patients with Nonalcoholic Steatohepatitis: Implications for Noninvasive Diagnosis of NASH" International Journal of Molecular Sciences 19, no. 2: 603. https://doi.org/10.3390/ijms19020603
APA StyleDel Ben, M., Overi, D., Polimeni, L., Carpino, G., Labbadia, G., Baratta, F., Pastori, D., Noce, V., Gaudio, E., Angelico, F., & Mancone, C. (2018). Overexpression of the Vitronectin V10 Subunit in Patients with Nonalcoholic Steatohepatitis: Implications for Noninvasive Diagnosis of NASH. International Journal of Molecular Sciences, 19(2), 603. https://doi.org/10.3390/ijms19020603