
The Relevance of Noncoding DNA Variations of Paraoxonase Gene Cluster in Atherosclerosis-Related Diseases


Abstract
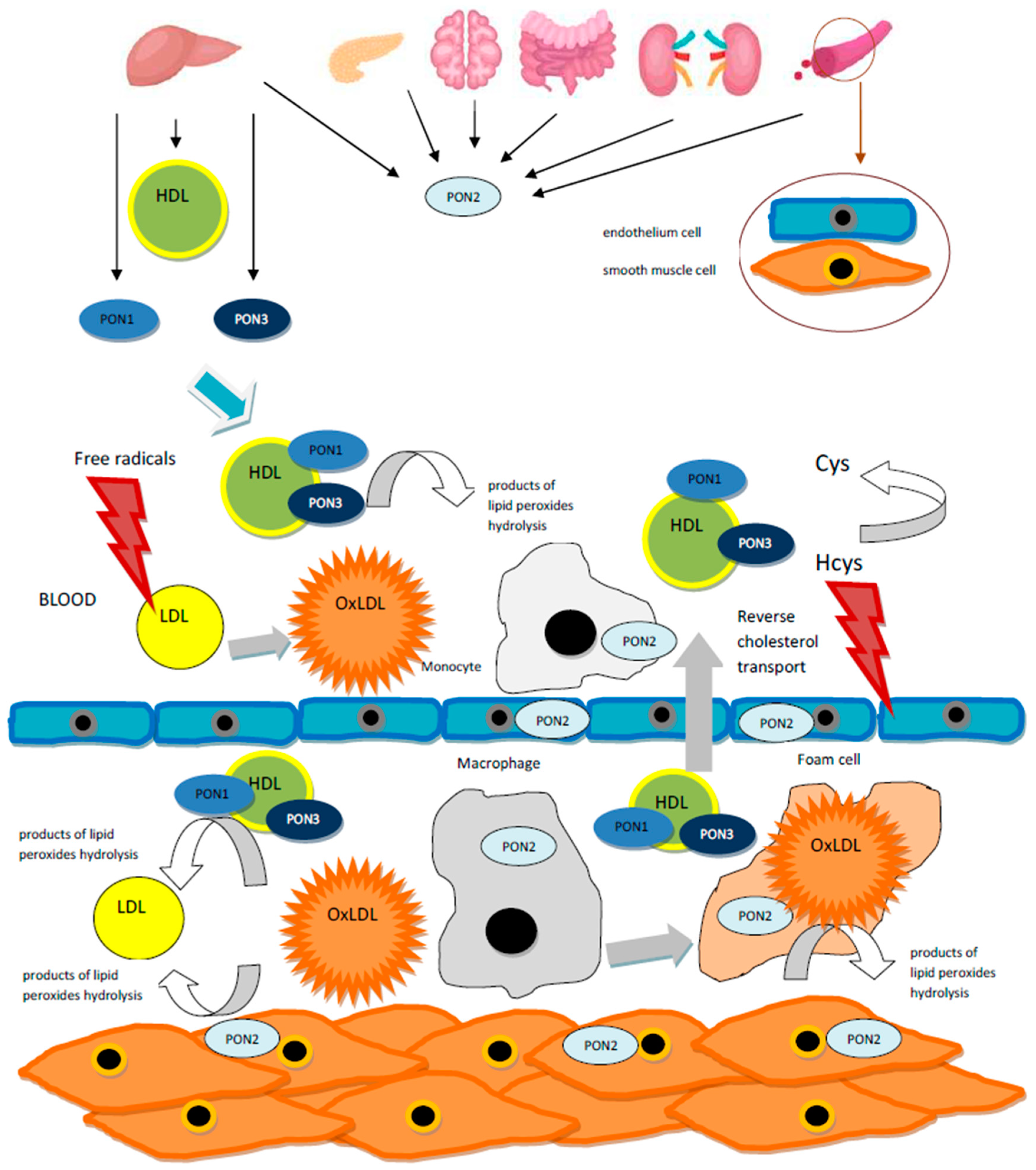
:1. Introduction
2. Search Strategy and Data Extraction
3. Effects of Genetic Polymorphism on PONs Activity and Concentration
4. PONs Polymorphism and Lipid Profile
5. Clinical Relevance of PONs Polymorphism
5.1. Atherosclerosis–Related Diseases and PONs Polymorphism
5.1.1. Coronary Heart Disease
5.1.2. Stroke
5.1.3. Diabetes
5.1.4. Peripheral Artery Disease, Dementia and Renal Disease
5.2. The Response to Drugs
5.2.1. Antiplatelet Drugs
5.2.2. Statins
6. Conclusions
- The noncoding DNA variations of PON gene cluster exert a considerable effect on the occurrence and the course of atherosclerosis-related diseases.
- The estimation of the predictive role of the PON gene polymorphism in the general population requires large prospective studies with appropriate accuracy of conduct. The data achieved until now allow us to consider PON cluster gene variations as additional atherosclerosis risk factors in selected individuals (for example in young patients without conventional risk factors who experienced cardiovascular events). Furthermore, in populations particularly prone to atherosclerosis, such as diabetic patients, these polymorphisms may serve as additional predictors of the risk of atherosclerosis.
- The issue of paramount importance that should be studied is the relationship between noncoding DNA PON genes variations and the drugs’ response, especially the clopidogrel resistance and adverse effects of statins occurrence, because the identification of compromised patients’ possible complications may enable the tailored therapy administration.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
Abbreviations
| A | adenine |
| AD | Alzheimer’s disease |
| ADP | adenosine diphosphate |
| AhR | arylhydrocarbon receptor |
| apoB | apolipoprotein B |
| AREase | arylesterase |
| ARIC | Atherosclerosis Risk in Communities |
| ARU | aspirin reaction units |
| C | cytosine |
| CABG | coronary artery bypass—grafting |
| CAD | coronary artery disease |
| CHD | coronary heart disease |
| CpG | cytosine phosphate guanine |
| del | deletion |
| DM | diabetes mellitus |
| DNA | deoxyribonucleic acid |
| ECTIM | EtudeCas – Temoins de l’InfarctusduMyocarde |
| FTO | fat mass and obesity associated |
| EDTA | ethylenediaminetetraacetic acid |
| GDM | gestational diabetes mellitus |
| GIR | glucose infusion rate |
| Hcy | homocysteine |
| HDL | high density lipoprotein |
| HIV | human immunodeficiency virus |
| IMT | intima media thickness |
| IL-6 | interleukin 6 |
| Ins | insertion |
| IS | ischemic stroke |
| ITGAL | integrin alpha L |
| KB | kilo bases |
| LD | linkage dysequilibium |
| LDL | low density protein |
| LDL maxox | LDL maximum oxidance |
| MACE | major adverse cardiac events |
| MD | mixed dementia |
| MIRAGE | Multi Institutional Research of Alzheimer Genetic Epidemiology |
| miRNA | micro ribonucleic acid |
| mRNA | messenger ribonucleic acid |
| NF | nuclear factor |
| OP | organophosphorous |
| Ox-LDL | oxidized LDL |
| PAD | peripheral artery disease |
| PCI | percutaneus coronary intervention |
| PKC | protein kinase C |
| PON | paraoxonase |
| QTL | quantitive trait loci |
| SERPINA | Serpin Family A |
| SLE | systemic lupus erytrematosus |
| SNP | single nucleotide polymorphism |
| SNV | single nucleotide variation |
| Sp1 | specific protein 1 |
| SREBP 2 | sterol regulatory binding protein 2 |
| T | tymine |
| T2DM | type 2 diabetes mellitus |
| tagSNP | tagging SNP |
| UTR | untranslated region |
| VD | vascular dementia |
| VLDL | very low density proteins |
| XRE | xenobiotic regulatory element |
References
- Humbert, R.; Adler, D.A.; Disteche, C.M.; Hassett, C.; Omiecinski, C.J.; Furlong, C.E. The molecular basis of the human serum paraoxonase activity polymorphism. Nat. Genet. 1993, 3, 73–76. [Google Scholar] [CrossRef]
- Robertson, K.S.; Hawe, E.; Miller, G.J.; Talmud, P.J.; Humphries, S.E. Human paraoxonase gene cluster polymorphisms as predictors of coronary heart disease risk in the prospective Northwick Park Heart Study II. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2003, 1639, 203–212. [Google Scholar] [CrossRef] [Green Version]
- Satoh, T.; Taylor, P.; Bosron, W.F.; Sanghani, S.P.; Hosokawa, M.; La Du, B.N. Current progress on esterases: From molecular structure to function. Drug Metab. Dispos. 2002, 30, 488–493. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, M.; Shinohara, M.; Sakoh, C.; Kataoka, M.; Shimizu, S. Lactone-ring-cleaving enzyme: Genetic analysis, novel RNA editing, and evolutionary implications. Proc. Natl. Acad. Sci. USA 1998, 95, 12787–12792. [Google Scholar] [CrossRef] [Green Version]
- Draganov, D.I.; La Du, B.N. Pharmacogenetics of paraoxonases: A brief review. NaunynSchmiedebergs Arch. Pharmacol. 2004, 369, 78–88. [Google Scholar] [CrossRef]
- Hassett, C.; Richter, R.J.; Humbert, R.; Chapline, C.; Crabb, J.W.; Omiecinski, C.J.; Furlong, C.E. Characterization of cDNA clones encoding rabbit and human serum paraoxonase: The mature protein retains its signal sequence. Biochememistry 1991, 30, 10141–10149. [Google Scholar] [CrossRef]
- Mackness, M.I.; Mackness, B.; Durrington, P.N.; Connelly, P.W.; Hegele, R.A. Paraoxonase: Biochemistry, genetics and relationship to plasma lipoproteins. Curr. Opin. Lipidol. 1996, 7, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Reddy, S.T.; Wadleigh, D.J.; Grijalva, V.; Ng, C.; Hama, S.; Gangopadhyay, A.; Shih, D.M.; Lusis, A.J.; Navab, M.; Fogelman, A.M. Human paraoxonase-3 is an HDL-associated enzyme with biological activity similar to paraoxonase-1 protein but is not regulated by oxidized lipids. Arterioscler. Thromb. Vasc. Biol. 2001, 21, 542–547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ng, C.J.; Wadleigh, D.J.; Gangopadhyay, A.; Hama, S.; Grijalva, V.R.; Navab, M.; Fogelman, A.M.; Reddy, S.T. Paraoxonase-2 Is a Ubiquitously Expressed Protein with Antioxidant Properties and Is Capable of Preventing Cell-mediated Oxidative Modification of Low Density Lipoprotein. J. Biol. Chem. 2001, 276, 44444–44449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazur, A. An enzyme in the animal organism capable of hydrolysing the phosphorus-fluorine bond of alkyl fluorophos-phates. J. Biol. Chem. 1946, 164, 271–289. [Google Scholar] [CrossRef]
- Aldridge, W.N. Serum esterases. 2. An enzyme hydrolysing diethyl p-nitrophenyl phosphate (E 600) and its identity with the A-esterase of mammalian sera. Biochem. J. 1953, 53, 117–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Available online: https://www.qmul.ac.uk/sbcs/iubmb/ (accessed on 3 December 2020).
- Davies, H.G.; Richter, R.J.; Keifer, M.; Broomfield, C.A.; Sowalla, J.; Furlong, C.E. The effect of the human serum paraoxonase polymorphism is reversed with diazoxon, soman and sarin. Nat. Genet. 1996, 14, 334–336. [Google Scholar] [CrossRef]
- Mu, X.; Yi, X.; Xiao, S.; Wang, C.; Chengshan, W.; Li, Y. Substrates for Paraoxonase. Curr. Pharm. Des. 2018, 24, 615–627. [Google Scholar] [CrossRef]
- Jakubowski, H. Calcium-Dependent Human Serum Homocysteine Thiolactone Hydrolase. A protective mechanism against protein N-homocysteinylation. J. Biol. Chem. 2000, 275, 3957–3962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bedi, B.; Yuan, Z.; Joo, M.; Zughaier, S.M.; Goldberg, J.B.; Arbiser, J.L.; Hart, C.M.; Sadikot, R.T. Enhanced Clearance of Pseudomonas aeruginosa by Peroxisome Proliferator-Activated Receptor Gamma. Infect. Immun. 2016, 84, 1975–1985. [Google Scholar] [CrossRef] [Green Version]
- Aviram, M.; Rosenblat, M.; Bisgaier, C.L.; Newton, R.S.; Primo-Parmo, S.L.; La Du, B.N. Paraoxonase inhibits high-density lipoprotein oxidation and preserves its functions. A possible peroxidative role for paraoxonase. J. Clin. Investig. 1998, 101, 1581–1590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Draganov, D.I.; Teiber, J.F.; Speelman, A.; Osawa, Y.; Sunahara, R.; La Du, B.N. Human paraoxonases (PON1, PON2, and PON3) are lactonases with overlapping and distinct substrate specificities. J. Lipid Res. 2005, 46, 1239–1247. [Google Scholar] [CrossRef] [Green Version]
- Yildiz, A.; Gür, M.; Yilmaz, R.; Demirbag, R.; Polat, M.; Selek, S.; Çelik, H.; Erel, O. Association of paraoxonase activity and coronary blood flow. Atherosclerosis 2008, 197, 257–263. [Google Scholar] [CrossRef]
- Tang, W.H.W.; Hartiala, J.; Fan, Y.; Wu, Y.; Stewart, A.F.; Erdmann, J.; Kathiresan, S.; Roberts, R.; McPherson, R.; Allayee, H.; et al. Clinical and Genetic Association of Serum Paraoxonase and Arylesterase Activities with Cardiovascular Risk. Arterioscler. Thromb. Vasc. Biol. 2012, 32, 2803–2812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, F.; Ding, J.; Chen, Q.; Zhuang, X.; Feng, Z.; Xu, L. Low Paraoxonase 1 Arylesterase Activity and High von Willebrand Factor Levels are Associated with Severe Coronary Atherosclerosis in Patients with Non-Diabetic Stable Coronary Artery Disease. Med. Sci. Monit. 2014, 20, 2421–2429. [Google Scholar] [CrossRef] [Green Version]
- Wysocka, A.; Cybulski, M.; Wysokiński, A.P.; Berbeć, H.; Stążka, J.; Zapolski, T. Paraoxonase 1 Activity, Polymorphism and Atherosclerosis Risk Factors in Patients Undergoing Coronary Artery Surgery. J. Clin. Med. 2019, 8, 441. [Google Scholar] [CrossRef] [Green Version]
- Schrader, C. Determinants of Paraoxonase 1 Status: Genes, Drugs and Nutrition. Curr. Med. Chem. 2011, 18, 5624–5643. [Google Scholar] [CrossRef]
- La Du, B.N. Human serum paraoxonase/arylesterase. In Pharmacogenetics of Drug Metabolism; Kalow, W., Ed.; Pergamon Press: New York, NY, USA, 1992; pp. 51–91. [Google Scholar]
- McDaniel, C.Y.; Dail, M.B.; Wills, R.W.; Chambers, H.W.; Chambers, J.E. Paraoxonase 1 Polymorphisms Within a Mississippi USA Population as Possible Biomarkers of Enzyme Activities Associated with Disease Susceptibility. Biochem. Genet. 2014, 52, 509–523. [Google Scholar] [CrossRef]
- Phuntuwate, W.; Suthisisang, C.; Koanantakul, B.; Mackness, I.M.; Mackness, B. Paraoxonase 1 status in the Thai population. J. Hum. Genet. 2005, 50, 293–300. [Google Scholar] [CrossRef] [Green Version]
- Suehiro, T.; Nakamura, T.; Inoue, M.; Shiinoki, T.; Ikeda, Y.; Kumon, Y.; Shindo, M.; Tanaka, H.; Hashimoto, K. A polymorphism upstream from the human paraoxonase (PON1) gene and its association with PON1 expression. Atherosclerosis 2000, 150, 295–298. [Google Scholar] [CrossRef]
- Leviev, I.; James, R.W. Promoter polymorphisms of human paraoxonase PON1 gene and serum paraoxonase activities and concentrations. Arterioscler. Thromb. Vasc. Biol. 2000, 20, 516–521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brophy, V.H.; Hastings, M.D.; Clendenning, J.B.; Richter, R.J.; Jarvik, G.P.; Furlong, C.E. Polymorphisms in the human paraoxonase (PON1) promoter. Pharmacogenetics 2001, 11, 77–84. [Google Scholar] [CrossRef]
- Thameem, F.; He, X.; Voruganti, V.S.; Nath, S.D.; Fanti, P.; Blangero, J.; Maccluer, J.W.; Comuzzie, A.G.; Arar, N.H.; Abboud, H.E. Evaluation of Polymorphisms in Paraoxonase 2 (PON2) Gene and Their Association with Cardiovascular-Renal Disease Risk in Mexican Americans. Kidney Blood Press. Res. 2009, 32, 200–204. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.S.; Burt, A.A.; Ranchalis, J.E.; Richter, R.J.; Marshall, J.K.; Eintracht, J.F.; Rosenthal, E.A.; Furlong, C.E.; Jarvik, G.P. Additional Common Polymorphisms in thePONGene Cluster Predict PON1 Activity but Not Vascular Disease. J. Lipids 2012, 2012, 476316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Priyanka, K.; Singh, S.; Gill, K. Paraoxonase 3: Structure and Its Role in Pathophysiology of Coronary Artery Disease. Biomolecules 2019, 9, 817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campo, S.; Sardo, A.M.; CAmpo, G.M.; Avenoso, A.; Castaldo, M.; D’Ascola, A.; Giunta, E.; Calatroni, A.; Saitta, A. Identification of paraoxonase 3 gene (PON3) missense mutations in a population of southern Italy. Mutat. Res. Fundam. Mol. Mech. Mutagen. 2004, 546, 75–80. [Google Scholar] [CrossRef]
- Wang, X.; Fan, Z.; Huang, J.; Su, S.; Yu, Q.; Zhao, J.; Hui, R.; Yao, Z.; Shen, Y.; Qiang, B.; et al. Extensive Association Analysis Between Polymorphisms of PON Gene Cluster with Coronary Heart Disease in Chinese Han Population. Arterioscler. Thromb. Vasc. Biol. 2003, 23, 328–334. [Google Scholar] [CrossRef] [Green Version]
- Aragonès, G.; Guardiola, M.; Barreda, M.; Marsillach, J.; Beltrán-Debón, R.; Rull, A.; Mackness, B.; Mackness, M.; Joven, J.; Simó, J.M.; et al. Measurement of serum PON-3 concentration: Method evaluation, reference values, and influence of genotypes in a population-based study. J. Lipid Res. 2011, 52, 1055–1061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furlong, C.E.; Marsillach, J.; Jarvik, G.P.; Costa, L.G. Paraoxonases-1, -2 and -3: What are their functions? Chem. Interact. 2016, 259, 51–62. [Google Scholar] [CrossRef] [Green Version]
- Mackness, M.; Mackness, B. Human paraoxonase-1 (PON1): Gene structure and expression, promiscuous activities and multiple physiological roles. Gene 2015, 567, 12–21. [Google Scholar] [CrossRef] [Green Version]
- Shunmoogam, N.; Naidoo, P.; Chilton, R. Paraoxonase (PON)-1: A brief overview on genetics, structure, polymorphisms and clinical relevance. Vasc. Health Risk Manag. 2018, 14, 137–143. [Google Scholar] [CrossRef] [Green Version]
- She, Z.-G.; Chen, H.-Z.; Yan, Y.; Li, H.; Liu, D.-P. The Human Paraoxonase Gene Cluster As a Target in the Treatment of Atherosclerosis. Antioxid. Redox Signal. 2012, 16, 597–632. [Google Scholar] [CrossRef]
- Mackness, B.; Davies, G.K.; Turkie, W.; Lee, E.; Roberts, D.H.; Hill, E.; Roberts, C.; Durrington, P.N.; Mackness, M.I. Paraoxonase Status in Coronary Heart Disease: Are activity and concentration more important than genotype? Arterioscler. Thromb. Vasc. Biol. 2001, 21, 1451–1457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Primo-Parmo, S.L.; Sorenson, R.C.; Teiber, J.; Du, B.N. The Human Serum Paraoxonase/Arylesterase Gene (PON1) Is One Member of a Multigene Family. Genomics 1996, 33, 498–507. [Google Scholar] [CrossRef] [PubMed]
- Clendenning, J.B.; Humbert, R.; Green, E.D.; Wood, C.; Traver, D.; Furlong, C.E. Structural Organization of the HumanPON1Gene. Genomics 1996, 35, 586–589. [Google Scholar] [CrossRef]
- Deakin, S.; Leviev, I.; Brulhart-Meynet, M.-C.; James, R.W. Paraoxonase-1 promoter haplotypes and serum paraoxonase: A predominant role for polymorphic position −107, implicating the Sp1 transcription factor. Biochem. J. 2003, 372, 643–649. [Google Scholar] [CrossRef] [Green Version]
- Kumon, Y.; Suehiro, T.; Ikeda, Y.; Hashimoto, K. Human paraoxonase-1 gene expression by HepG2 cells is downregulated by interleukin-1β and tumor necrosis factor-α, but is upregulated by interleukin-6. Life Sci. 2003, 73, 2807–2815. [Google Scholar] [CrossRef]
- Deakin, S.; Leviev, I.; Guernier, S.; James, R.W. Simvastatin Modulates Expression of the PON1 Gene and Increases Serum Paraoxonase: A role for sterol regulatory element-binding protein-2. Arterioscler. Thromb. Vasc. Biol. 2003, 23, 2083–2089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osaki, F.; Ikeda, Y.; Suehiro, T.; Ota, K.; Tsuzura, S.; Arii, K.; Kumon, Y.; Hashimoto, K. Roles of Sp1 and protein kinase C in regulation of human serum paraoxonase 1 (PON1) gene transcription in HepG2 cells. Atherosclerosis 2004, 176, 279–287. [Google Scholar] [CrossRef]
- Gouédard, C.; Barouki, R.; Morel, Y. Dietary Polyphenols Increase Paraoxonase 1 Gene Expression by an Aryl Hydrocarbon Receptor-Dependent Mechanism. Mol. Cell. Biol. 2004, 24, 5209–5222. [Google Scholar] [CrossRef] [Green Version]
- Li, W.-F.; Costa, L.G.; Richter, R.J.; Hagen, T.; Shih, D.M.; Tward, A.; Lusis, A.J.; Furlong, C.E. Catalytic efficiency determines the in-vivo efficacy of PON1 for detoxifying organophosphorus compounds. Pharmacogenetics 2000, 10, 767–779. [Google Scholar] [CrossRef] [PubMed]
- Mackness, B.I.; Mackness, M.; Arrol, S.; Turkie, W.; Durrington, P.N. Effect of the human serum paraoxonase 55 and 192 genetic polymorphisms on the protection by high density lipoprotein against low density lipoprotein oxidative modification. FEBS Lett. 1998, 423, 57–60. [Google Scholar] [CrossRef] [Green Version]
- Aviram, M.; Billecke, S.; Sorenson, R.; Bisgaier, C.; Newton, R.; Rosenblat, M.; Erogul, J.; Hsu, C.; Dunlop, C.; La Du, B. Paraoxonase Active Site Required for Protection Against LDL Oxidation Involves Its Free Sulfhydryl Group and Is Different from That Required for Its Arylesterase/Paraoxonase Activities: Selective action of human paraoxonaseallozymes Q and R. Arterioscler. Thromb. Vasc. Biol. 1998, 18, 1617–1624. [Google Scholar] [CrossRef] [Green Version]
- Brophy, V.H.; Jarvik, G.P.; Richter, R.J.; Rozek, L.S.; Schellenberg, G.D.; Furlong, C.E. Analysis of paraoxonase (PON1) L55M status requires both genotype and phenotype. Pharmacogenetics 2000, 10, 453–460. [Google Scholar] [CrossRef]
- Garin, M.C.; James, R.W.; Dussoix, P.; Blanché, H.; Passa, P.; Froguel, P.; Ruiz, J. Paraoxonase polymorphism Met-Leu54 is associated with modified serum concentrations of the enzyme. A possible link between the paraoxonase gene and increased risk of cardiovascular disease in diabetes. J. Clin. Investig. 1997, 99, 62–66. [Google Scholar] [CrossRef] [Green Version]
- Leviev, I.; Negro, F.; James, R.W. Two Alleles of the Human Paraoxonase Gene Produce Different Amounts of mRNA.An explanation for differences in serum concentrations of paraoxonase associated with the (Leu-Met54) polymorphism. Arterioscler. Thromb. Vasc. Biol. 1997, 17, 2935–2939. [Google Scholar] [CrossRef]
- Brophy, V.H.; Jampsa, R.L.; Clendenning, J.B.; McKinstry, L.A.; Jarvik, G.P.; Furlong, C.E. Effects of 5′ Regulatory-Region Polymorphisms on Paraoxonase-Gene (PON1) Expression. Am. J. Hum. Genet. 2001, 68, 1428–1436. [Google Scholar] [CrossRef] [Green Version]
- Ray, A. A SAF Binding Site in the Promoter Region of Human γ-Fibrinogen Gene Functions as an IL-6 Response Element. J. Immunol. 2000, 165, 3411–3417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furlong, C.; Richter, R.; Li, W.-F.; Brophy, V.; Carlson, C.; Rieder, M.; Nickerson, D.; Costa, L.; Ranchalis, J.; Lusis, A.; et al. The Functional Consequences of Polymorphisms in the Human PON1 Gene. In The Paraoxonases: Their Role in Disease Development and Xenobiotic Metabolism; Mackness, M.M.B., Aviram, M., Paragh, G., Eds.; Springer: Dordrecht, The Netherlands, 2008; pp. 267–281. [Google Scholar]
- Carlson, C.S.; Heagerty, P.J.; Hatsukami, T.S.; Richter, R.J.; Ranchalis, J.; Lewis, J.; Bacus, T.J.; McKinstry, L.A.; Schellenberg, G.D.; Rieder, M.; et al. TagSNP analyses of the PON gene cluster: Effects on PON1 activity, LDL oxidative susceptibility, and vascular disease. J. Lipid Res. 2006, 47, 1014–1024. [Google Scholar] [CrossRef] [Green Version]
- Eom, S.-Y.; Kim, Y.-S.; Lee, C.-J.; Lee, C.-H.; Kim, Y.-D.; Kim, H. Effects of Intronic and Exonic Polymorphisms of Paraoxonase 1 (PON1) Gene on Serum PON1 Activity in a Korean Population. J. Korean Med. Sci. 2011, 26, 720–725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa, L.G.; Vitalone, A.; Cole, T.B.; Furlong, C.E. Modulation of paraoxonase (PON1) activity. Biochem. Pharmacol. 2005, 69, 541–550. [Google Scholar] [CrossRef]
- Liu, M.-E.; Liao, Y.-C.; Lin, R.-T.; Wang, Y.-S.; Hsi, E.; Lin, H.-F.; Chen, K.-C.; Juo, S.-H.H. A functional polymorphism of PON1 interferes with microRNA binding to increase the risk of ischemic stroke and carotid atherosclerosis. Atherosclerosis 2013, 228, 161–167. [Google Scholar] [CrossRef] [PubMed]
- Winnier, D.A.; Rainwater, D.L.; Cole, S.A.; Dyer, T.D.; Blangero, J.; Maccluer, J.W.; Mahaney, M.C. Multiple QTLs Influence Variation in Paraoxonase 1 Activity in Mexican Americans. Hum. Biol. 2006, 78, 341–352. [Google Scholar] [CrossRef] [PubMed]
- Winnier, D.A.; Rainwater, D.L.; Cole, S.A.; Williams, J.T.; Dyer, T.D.; Blangero, J.; Maccluer, J.W.; Mahaney, M.C. Sex-specific QTL effects on variation in paraoxonase 1 (PON1) activity in Mexican Americans. Genet. Epidemiol. 2006, 31, 66–74. [Google Scholar] [CrossRef]
- Nolan, D.; Kraus, W.E.; Hauser, E.; Li, Y.-J.; Thompson, D.K.; Johnson, J.; Chen, H.-C.; Nelson, S.; Haynes, C.; Gregory, S.G.; et al. Genome-Wide Linkage Analysis of Cardiovascular Disease Biomarkers in a Large, Multigenerational Family. PLoS ONE 2013, 8, e71779. [Google Scholar] [CrossRef]
- Koda, Y.; Tachida, H.; Soejima, M.; Takenaka, O.; Kimura, H. Population differences in DNA sequence variation and linkage disequilibrium at the PON1 gene. Ann. Hum. Genet. 2004, 68, 110–119. [Google Scholar] [CrossRef]
- Jarvik, G.P.; Jampsa, R.; Richter, R.J.; Carlson, C.S.; Rieder, M.J.; Nickerson, D.A.; Furlong, C.E. Novel paraoxonase (PON1) nonsense and missense mutations predicted by functional genomic assay of PON1 status. Pharmacogenetics 2003, 13, 291–295. [Google Scholar] [CrossRef]
- Kim, D.S.; Burt, A.A.; Crosslin, D.R.; Robertson, P.D.; Ranchalis, J.E.; Boyko, E.J.; Nickerson, D.A.; Furlong, C.E.; Jarvik, G.P. Novel common and rare genetic determinants of paraoxonase activity: FTO, SERPINA12, and ITGAL. J. Lipid Res. 2013, 54, 552–560. [Google Scholar] [CrossRef] [Green Version]
- Morita, S.; Takahashi, R.-U.; Yamashita, R.; Toyoda, A.; Horii, T.; Kimura, M.; Fujiyama, A.; Nakai, K.; Tajima, S.; Matoba, R.; et al. Genome-Wide Analysis of DNA Methylation and Expression of MicroRNAs in Breast Cancer Cells. Int. J. Mol. Sci. 2012, 13, 8259–8272. [Google Scholar] [CrossRef]
- Su, J.; Li, J.; Yu, Q.; Xu, X.; Wang, J.; Yang, J.; Li, X.; Chen, X. Association of PON1 gene promoter DNA methylation with the risk of Clopidogrel resistance in patients with coronary artery disease. J. Clin. Lab. Anal. 2019, 33, e22867. [Google Scholar] [CrossRef]
- Stoltz, D.A.; Ozer, E.A.; Recker, T.J.; Estin, M.; Yang, X.; Shih, D.M.; Lusis, A.J.; Zabner, J. A Common Mutation in Paraoxonase-2 Results in Impaired Lactonase Activity. J. Biol. Chem. 2009, 284, 35564–35571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mochizuki, H.; Scherer, S.W.; Xi, T.; Nickle, D.C.; Majer, M.; Huizenga, J.J.; Tsui, L.-C.; Prochazka, M. Human PON2 gene at 7q21.3: Cloning, multiple mRNA forms, and missense polymorphisms in the coding sequence. Gene 1998, 213, 149–157. [Google Scholar] [CrossRef]
- Aragonès, G.; García-Heredia, A.; Guardiola, M.; Rull, A.; Beltrán-Debón, R.; Marsillach, J.; Alonso-Villaverde, C.; Mackness, B.; Mackness, M.; Pedro-Botet, J.; et al. Serum paraoxonase-3 concentration in HIV-infected patients. Evidence for a protective role against oxidation. J. Lipid Res. 2012, 53, 168–174. [Google Scholar] [CrossRef] [Green Version]
- Rull, A.; García, R.; Fernández-Sender, L.; García-Heredia, A.; Aragonès, G.; Beltrán-Debón, R.; Marsillach, J.; Alegret, J.M.; Martín-Paredero, V.; Mackness, B.; et al. Serum paraoxonase-3 concentration is associated with insulin sensitivity in peripheral artery disease and with inflammation in coronary artery disease. Atherosclerosis 2012, 220, 545–551. [Google Scholar] [CrossRef] [PubMed]
- Riedmaier, S.; Klein, K.; Winter, S.; Hofmann, U.; Schwab, M.; Zanger, U.M. Paraoxonase (PON1 and PON3) Polymorphisms: Impact on Liver Expression and Atorvastatin-Lactone Hydrolysis. Front. Pharmacol. 2011, 2, 41. [Google Scholar] [CrossRef] [Green Version]
- Childs, B.G.; Baker, D.J.; Wijshake, T.; Conover, C.A.; Campisi, J.; Van Deursen, J.M. Senescent intimal foam cells are deleterious at all stages of atherosclerosis. Science 2016, 354, 472–477. [Google Scholar] [CrossRef]
- März, W.; Kleber, M.E.; Scharnagl, H.; Speer, T.; Zewinger, S.; Ritsch, A.; Parhofer, K.G.; Von Eckardstein, A.; Landmesser, U.; Laufs, U. HDL cholesterol: Reappraisal of its clinical relevance. Clin. Res. Cardiol. 2017, 106, 663–675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garin, M.-C.B.; Moren, X.; James, R.W. Paraoxonase-1 and serum concentrations of HDL-cholesterol and apoA-I. J. Lipid Res. 2006, 47, 515–520. [Google Scholar] [CrossRef] [Green Version]
- Campo, S.; Sardo, M.A.; Trimarchi, G.; Bonaiuto, M.; Fontana, L.; Castaldo, M.; Bonaiuto, A.; Saitta, C.; Bitto, A.; Manduca, B.; et al. Association between serum paraoxonase (PON1) gene promoter T(-107)C polymorphism, PON1 activity and HDL levels in healthy Sicilian octogenarians. Exp. Gerontol. 2004, 39, 1089–1094. [Google Scholar] [CrossRef] [PubMed]
- Tsuzura, S.; Ikeda, Y.; Suehiro, T.; Ota, K.; Osaki, F.; Arii, K.; Kumon, Y.; Hashimoto, K. Correlation of plasma oxidized low-density lipoprotein levels to vascular complications and human serum paraoxonase in patients with type 2 diabetes. Metabolism 2004, 53, 297–302. [Google Scholar] [CrossRef]
- She, Z.-G.; Zheng, W.; Wei, Y.-S.; Chen, H.-Z.; Wang, A.-B.; Li, H.-L.; Liu, G.; Zhang, R.; Liu, J.-J.; Stallcup, W.B.; et al. Human Paraoxonase Gene Cluster Transgenic Overexpression Represses Atherogenesis and Promotes Atherosclerotic Plaque Stability in ApoE-Null Mice. Circ. Res. 2009, 104, 1160–1168. [Google Scholar] [CrossRef] [Green Version]
- Aviram, M.; Hardak, E.; Vaya, J.; Mahmood, S.; Milo, S.; Hoffman, A.; Billicke, S.; Draganov, D.; Rosenblat, M. Human Serum Paraoxonases (PON1) Q and R Selectively Decrease Lipid Peroxides in Human Coronary and Carotid Atherosclerotic Lesions: PON1 esterase and peroxidase-like activities. Circulation 2000, 101, 2510–2517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanghera, D.K.; Aston, C.E.; Saha, N.; Kamboh, M.I. DNA Polymorphisms in Two Paraoxonase Genes (PON1 and PON2) Are Associated with the Risk of Coronary Heart Disease. Am. J. Hum. Genet. 1998, 62, 36–44. [Google Scholar] [CrossRef] [Green Version]
- Sanghera, D.K.; Saha, N.; Aston, C.E.; Kamboh, M.I. Genetic Polymorphism of Paraoxonase and the Risk of Coronary Heart Disease. Arterioscler. Thromb. Vasc. Biol. 1997, 17, 1067–1073. [Google Scholar] [CrossRef]
- Godbole, C.; Thaker, S.; Kerkar, P.; Nadkar, M.; Gogtay, N.; Thatte, U. Association of PON1 gene polymorphisms and enzymatic activity with risk of coronary artery disease. Future Cardiol. 2021, 17, 119–126. [Google Scholar] [CrossRef]
- Shahsavari, G.; Nouryazdan, N.; Adibhesami, G.; Birjandi, M. Genetic associations and serum paraoxonase levels with atherosclerosis in western Iranian patients. Mol. Biol. Rep. 2020, 47, 5137–5144. [Google Scholar] [CrossRef]
- Sanghera, D.K.; Saha, N.; Kamboh, M.I. The codon 55 polymorphism in the paraoxonase 1 gene is not associated with the risk of coronary heart disease in Asian Indians and Chinese. Atherosclerosis 1998, 136, 217–223. [Google Scholar] [CrossRef]
- Zeng, Q.; Zeng, J. A meta-analysis on relationship between paraoxonase 1 polymorphisms and atherosclerotic cardiovascular diseases. Life Sci. 2019, 232, 116646. [Google Scholar] [CrossRef]
- James, R.W.; Leviev, I.; Ruiz, J.; Passa, P.; Froguel, P.; Garin, M.C. Promoter polymorphism T(-107)C of the paraoxonase PON1 gene is a risk factor for coronary heart disease in type 2 diabetic patients. Diabetes 2000, 49, 1390–1393. [Google Scholar] [CrossRef] [Green Version]
- Leviev, I.; Righetti, A.; James, R.W. Paraoxonase promoter polymorphism T(-107)C and relative paraoxonase deficiency as determinants of risk of coronary artery disease. J. Mol. Med. 2001, 79, 457–463. [Google Scholar] [CrossRef] [PubMed]
- Leviev, I.; Poirier, O.; Nicaud, V.; Evans, A.; Kee, F.; Arveiler, D.; Morrisson, C.; Cambien, F.; James, R.W. High expressor paraoxonase PON1 gene promoter polymorphisms are associated with reduced risk of vascular disease in younger coronary patients. Atherosclerosis 2002, 161, 463–467. [Google Scholar] [CrossRef]
- Su, S.-Y.; Chen, J.-H.; Huang, J.-F.; Wang, X.-L.; Zhao, J.-G.; Shen, Y.; Qiang, B.-Q.; Gu, D.-F. Paraoxonase gene cluster variations associated with coronary heart disease in Chinese Han women. Chin. Med. J. 2005, 118, 1167–1174. [Google Scholar] [PubMed]
- Najafi, M.; Gohari, L.H.; Firoozrai, M. Paraoxonase 1 gene promoter polymorphisms are associated with the extent of stenosis in coronary arteries. Thromb. Res. 2009, 123, 503–510. [Google Scholar] [CrossRef]
- Zapolski, T.; Wysocka, A.; Cybulski, M.; Berbeć, H.; Wysokiński, A.; Stążka, J. Prognostic value of paraoxonase 1 in patients undergoing coronary artery bypass grafting surgery. Med. Sci. Monit. 2014, 20, 594–600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, N.; Singh, S.; Maturu, V.N.; Sharma, Y.P.; Gill, K.D. Paraoxonase 1 (PON1) Polymorphisms, Haplotypes and Activity in Predicting CAD Risk in North-West Indian Punjabis. PLoS ONE 2011, 6, e17805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grubisa, I.; Otasevic, P.; Dimkovic, N.; Nedeljkovic, I.; Toljic, B.; Vucinic, N. Genetic polymorphisms of paraoxonase 1 and susceptibility to atherogenesis. Srp. Arh. Celok. Lek. 2013, 141, 629–633. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, J.G.; Keavney, B.D.; Watkins, H.; Collins, R.; Danesh, J. Four paraoxonase gene polymorphisms in 11 212 cases of coronary heart disease and 12 786 controls: Meta-analysis of 43 studies. Lancet 2004, 363, 689–695. [Google Scholar] [CrossRef]
- Wang, M.; Lang, X.; Zou, L.; Huang, S.; Xu, Z. Four genetic polymorphisms of paraoxonase gene and risk of coronary heart disease: A meta-analysis based on 88 case-control studies. Atherosclerosis 2011, 214, 377–385. [Google Scholar] [CrossRef]
- Mackness, B.; Turkie, W.; Mackness, M. Paraoxonase-1 (PON1) promoter region polymorphisms, serum PON1 status and coronary heart disease. Arch. Med. Sci. 2013, 1, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Franey, E.G.; Kritz-Silverstein, D.; Richard, E.L.; Alcaraz, J.E.; Nievergelt, C.M.; Shaffer, R.A.; Bhatnagar, V. Association of Race and Major Adverse Cardiac Events (MACE): The Atherosclerosis Risk in Communities (ARIC) Cohort. J. Aging Res. 2020, 2020, 7417242. [Google Scholar] [CrossRef] [Green Version]
- Demirdöğen, B.C.; Türkanoğlu, A.; Bek, S.; Sanisoglu, Y.; Demirkaya, Ş.; Vural, O.; Arınç, E.; Adali, O. Paraoxonase/arylesterase ratio, PON1 192Q/R polymorphism and PON1 status are associated with increased risk of ischemic stroke. Clin. Biochem. 2008, 41, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Liu, Y.-H.; Yang, Q.-D.; Xiao, B.; Ge, L.; Zhang, N.; Xia, J.; Zhang, L.; Liu, Z.-J. Human serum paraoxonase gene polymorphisms, Q192R and L55M, are not associated with the risk of cerebral infarction in Chinese Han population. Neurol. Res. 2006, 28, 549–554. [Google Scholar] [CrossRef]
- Martínez-Salazar, M.F.; Soriano-Martínez, M.D.L.L.; Juantorena-Ugas, A.; Almenares-López, D.; Yescas, P.; Boll, M.-C.; Monroy-Noyola, A. Paraoxonase-1 polymorphisms and cerebral ischemic stroke: A pilot study in mexican patients. Colomb. Med. 2018, 49, 223–227. [Google Scholar] [CrossRef]
- Dahabreh, I.J.; Kitsios, G.D.; Kent, D.M.; Trikalinos, T.A. Paraoxonase 1 polymorphisms and ischemic stroke risk: A systematic review and meta-analysis. Genet. Med. 2010, 12, 606–615. [Google Scholar] [CrossRef] [Green Version]
- Voetsch, B.; Benke, K.S.; Panhuysen, C.I.; Damasceno, B.P.; Loscalzo, J. The Combined Effect of Paraoxonase Promoter and Coding Region Polymorphisms on the Risk of Arterial Ischemic Stroke Among Young Adults. Arch. Neurol. 2004, 61, 351–356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demirdöğen, B.C.; Demirkaya, S.; Türkanoğlu, A.; Bek, S.; Arinç, E.; Adali, O. Analysis of paraoxonase 1 (PON1) genetic polymorphisms and activities as risk factors for ischemic stroke in Turkish population. Cell Biochem. Funct. 2009, 27, 558–567. [Google Scholar] [CrossRef]
- Roest, M.; Jansen, A.C.; Barendrecht, A.; Leus, F.R.; Kastelein, J.J.; Voorbij, H.A. Variation at the paraoxonase gene locus contributes to carotid arterial wall thickness in subjects with familial hypercholesterolemia. Clin. Biochem. 2005, 38, 123–127. [Google Scholar] [CrossRef]
- Kim, N.S.; Kang, B.K.; Cha, M.H.; Oh, S.-M.; Ko, M.M.; Bang, O.-S. Association between PON1 5′-regulatory region polymorphisms, PON1 activity and ischemic stroke. Clin. Biochem. 2009, 42, 857–863. [Google Scholar] [CrossRef]
- Zhang, G.; Li, W.; Li, Z.; Lv, H.; Ren, Y.; Ma, R.; Li, X.; Kang, X.; Shi, Y.; Sun, Y. Association between paraoxonase gene and stroke in the Han Chinese population. BMC Med. Genet. 2013, 14, 16. [Google Scholar] [CrossRef] [Green Version]
- Pan, Y.; He, B.; Sun, H.; Xu, T.; Pan, B.; Wang, S.; Mei, Y. Susceptibility of PON1/PON2 Genetic Variations to Ischemic Stroke Risk in a Chinese Han Population. Pharmacogenom. Pers. Med. 2020, 13, 563–570. [Google Scholar] [CrossRef]
- Xiao, J.; Li, X.; Yuan, Q.; Zhang, S.; Qu, K.; Wu, B.; Wang, Y.; Duan, S. PON1 Hypermethylation and PON3 Hypomethylation are Associated with Risk of Cerebral Infarction. Curr. Neurovasc. Res. 2019, 16, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Hu, F.B.; Stampfer, M.J.; Haffner, S.M.; Solomon, C.G.; Willett, W.C.; Manson, J.E. Elevated Risk of Cardiovascular Disease Prior to Clinical Diagnosis of Type 2 Diabetes. Diabetes Care 2002, 25, 1129–1134. [Google Scholar] [CrossRef] [Green Version]
- Luo, J.-Q.; Ren, H.; Liu, M.-Z.; Fang, P.-F.; Xiang, D.-X. European versus Asian differences for the associations between paraoxonase-1 genetic polymorphisms and susceptibility to type 2 diabetes mellitus. J. Cell. Mol. Med. 2018, 22, 1720–1732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shakeri, R.; Khajeniazi, S.; Marjani, A. Association between promoter polymorphism (−108C > T) of paraoxonase1 gene and it’s paraoxonase activity in patients with Type2 diabetes in northern Iran. Clin. Chim. Acta 2017, 474, 34–37. [Google Scholar] [CrossRef] [PubMed]
- Gupta, N.; Binukumar, B.; Singh, S.; Sunkaria, A.; Kandimalla, R.; Bhansali, A.; Gill, K.D. Serum paraoxonase-1 (PON1) activities (PONase/AREase) and polymorphisms in patients with type 2 diabetes mellitus in a North-West Indian population. Gene 2011, 487, 88–95. [Google Scholar] [CrossRef]
- Flekac, M.; Skrha, J.; Zídková, K.; Lacinová, Z.; Hilgertová, J. Paraoxonase 1 gene polymorphisms and enzyme activities in di-abetes mellitus. Physiol. Res. 2008, 57, 717–726. [Google Scholar]
- Ikeda, Y.; Suehiro, T.; Arii, K.; Kumon, Y.; Hashimoto, K. High glucose induces transactivation of the human paraoxonase 1 gene in hepatocytes. Metabolism 2008, 57, 1725–1732. [Google Scholar] [CrossRef]
- Zhou, M.; Liu, X.-H.; Liu, Q.-Q.; Chen, M.; Bai, H.; Jiang, C.-Y.; Guan, L.-B.; Fan, P. Lactonase activity and status of paraoxonase 1 and oxidative stress in neonates of women with gestational diabetes mellitus. Pediatr. Res. 2020, 22, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Leviev, I.; Kalix, B.; Meynet, M.-C.B.; James, R.W. The paraoxonase PON1 promoter polymorphism C(-107)T is associated with increased serum glucose concentrations in non-diabetic patients. Diabetologia 2001, 44, 1177–1183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ikeda, Y.; Suehiro, T.; Ohsaki, F.; Arii, K.; Kumon, Y.; Hashimoto, K. Relationships between polymorphisms of the human serum paraoxonase gene and insulin sensitivity in Japanese patients with Type 2 diabetes. Diabetes Res. Clin. Pract. 2003, 60, 79–85. [Google Scholar] [CrossRef]
- Hofer, S.E.; Bennetts, B.; Chan, A.K.; Holloway, B.; Karschimkus, C.; Jenkins, A.J.; Silink, M.; Donaghue, K.C. Association between PON 1 polymorphisms, PON activity and diabetes complications. J. Diabetes Complicat. 2006, 20, 322–328. [Google Scholar] [CrossRef]
- Araki, S.; Makita, Y.; Canani, L.; Ng, D.; Warram, J.H.; Krolewski, A.S. Polymorphisms of human paraoxonase 1 gene (PON1) and susceptibility to diabetic nephropathy in Type I diabetes mellitus. Diabetologia 2000, 43, 1540–1543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernández-Aguilera, A.; Sepúlveda, J.; Rodríguez-Gallego, E.; Guirro, M.; García-Heredia, A.; Cabré, N.; Luciano-Mateo, F.; Fort-Gallifa, I.; Martín-Paredero, V.; Joven, J.; et al. Immunohistochemical Analysis of Paraoxonases and Chemokines in Arteries of Patients with Peripheral Artery Disease. Int. J. Mol. Sci. 2015, 16, 11323–11338. [Google Scholar] [CrossRef] [Green Version]
- Pasqualini, L.; Cortese, C.; Marchesi, S.; Siepi, D.; Pirro, M.; Vaudo, G.; Liberatoscioli, L.; Gnasso, A.; Schillaci, G.; Mannarino, E. Paraoxonase-1 activity modulates endothelial function in patients with peripheral arterial disease. Atherosclerosis 2005, 183, 349–354. [Google Scholar] [CrossRef]
- Rajković, M.G.; Barišić, K.; Juretić, D.; Grubišić, T.Ž.; Flegar-Meštrić, Z.; Rumora, L. Polymorphisms of pon1 and pon2 genes in hemodialyzed patients. Clin. Biochem. 2011, 44, 964–968. [Google Scholar] [CrossRef] [PubMed]
- Tripi, L.M.; Manzi, S.; Chen, Q.; Kenney, M.; Shaw, P.; Kao, A.; Bontempo, F.; Kammerer, C.; Kamboh, M.I. Relationship of serum paraoxonase 1 activity and paraoxonase 1 genotype to risk of systemic lupus erythematosus. Arthritis Rheum. 2006, 54, 1928–1939. [Google Scholar] [CrossRef]
- Kovács, T.J.; Harris, S.; Vas, T.K.; Seres, I.; Short, C.D.; Wittmann, I.K.; Paragh, G.; Mackness, M.I.; Mackness, B.; Durrington, P.N.; et al. Paraoxonase gene polymorphism and serum activity in progressive IgA nephropathy. J. Nephrol. 2006, 19, 732–738. [Google Scholar] [PubMed]
- Bednarska-Makaruk, M.E.; Krzywkowski, T.; Graban, A.; Lipczyńska-Łojkowska, W.; Bochyńska, A.; Rodo, M.; Wehr, H.; Ryglewicz, D.K. Original article Paraoxonase 1 (PON1) gene −108C>T and p.Q192R polymorphisms and arylesterase activity of the enzyme in patients with dementia. Folia Neuropathol. 2013, 2, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Cagnin, A.; Leon, A.; Vianello, D.; Colavito, D.; Favaretto, S.; Zarantonello, G.; Stecca, A.; Ermani, M.; Zambon, A. LDL Density and Oxidation are Modulated by PON1 Promoter Genotype in Patients with Alzheimer’s Disease. J. Alzheimers Dis. 2013, 34, 377–385. [Google Scholar] [CrossRef] [Green Version]
- Saeidi, M.; Shakeri, R.; Marjani, A.; Khajeniazi, S. Alzheimer’s Disease and Paraoxonase 1 (PON1) Gene Polymorphisms. Open Biochem. J. 2017, 11, 47–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erlich, P.M.; Lunetta, K.L.; Cupples, L.A.; Huyck, M.; Green, R.C.; Baldwin, C.T.; Farrer, L.A.; Auerbach, S.; Akomolafe, A.; Griffith, P.; et al. Polymorphisms in the PON gene cluster are associated with Alzheimer disease. Hum. Mol. Genet. 2006, 15, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Chapuis, J.; Boscher, M.; Bensemain, F.; Cottel, D.; Amouyel, P.; Lambert, J.-C. Association study of the paraoxonase 1 gene with the risk of developing Alzheimer’s disease. Neurobiol. Aging 2009, 30, 152–156. [Google Scholar] [CrossRef] [PubMed]
- Erlich, P.M.; Lunetta, K.L.; Cupples, L.A.; Abraham, C.R.; Green, R.C.; Baldwin, C.T.; Farrer, L.A. Serum paraoxonase activity is associated with variants in the PON gene cluster and risk of Alzheimer disease. Neurobiol. Aging 2012, 33, 1015.e7–1015.e23. [Google Scholar] [CrossRef] [Green Version]
- Cellini, E.; Tedde, A.; Bagnoli, S.; Nacmias, B.; Piacentini, S.; Bessi, V.; Bracco, L.; Sorbi, S. Association analysis of the paraoxonase-1 gene with Alzheimer’s disease. Neurosci. Lett. 2006, 408, 199–202. [Google Scholar] [CrossRef]
- Bednarska-Makaruk, M.; Graban, A.; Sobczyńska-Malefora, A.; Harrington, D.J.; Mitchell, M.; Voong, K.; Dai, L.; Łojkowska, W.; Bochyńska, A.; Ryglewicz, D.; et al. Homocysteine metabolism and the associations of global DNA methylation with selected gene polymorphisms and nutritional factors in patients with dementia. Exp. Gerontol. 2016, 81, 83–91. [Google Scholar] [CrossRef]
- Nie, Y.; Luo, D.; Yang, M.; Wang, Y.; Xiong, L.; Gao, L.; Liu, Y.; Liu, H. A Meta-Analysis on the Relationship of the PON Genes and Alzheimer Disease. J. Geriatr. Psychiatry Neurol. 2017, 30, 303–310. [Google Scholar] [CrossRef] [PubMed]
- Trenk, D.; Hochholzer, W. Genetics of platelet inhibitor treatment. Br. J. Clin. Pharmacol. 2014, 77, 642–653. [Google Scholar] [CrossRef] [Green Version]
- Bouman, H.J.; Schoemig, E.; Van Werkum, J.W.; Velder, J.; Hackeng, C.M.; Hirschhaeuser, C.; Waldmann, C.; Schmalz, H.-G.; Berg, J.M.T.; Taubert, D. Paraoxonase-1 is a major determinant of clopidogrel efficacy. Nat. Med. 2010, 17, 110–116. [Google Scholar] [CrossRef] [Green Version]
- Rideg, O.; Komócsi, A.; Magyarlaki, T.; Tőkés-Füzesi, M.; Miseta, A.; Kovács, G.L.; Aradi, D. Impact of genetic variants on post-clopidogrel platelet reactivity in patients after elective percutaneous coronary intervention. Pharmacogenomics 2011, 12, 1269–1280. [Google Scholar] [CrossRef] [PubMed]
- Fontana, P.; James, R.; Barazer, I.; Berdagué, P.; Schved, J.-F.; Rebsamen, M.; Vuilleumier, N.; Reny, J.-L. Relationship between paraoxonase-1 activity, its Q192R genetic variant and clopidogrel responsiveness in the ADRIE study. J. Thromb. Haemost. 2011, 9, 1664–1666. [Google Scholar] [CrossRef] [PubMed]
- Simon, T.; Steg, P.G.; Becquemont, L.; Verstuyft, C.; Kotti, S.; Schiele, F.; Ferrari, E.; Drouet, E.; Grollier, G.; Danchin, N. Effect of Paraoxonase-1 Polymorphism on Clinical Outcomes in Patients Treated with Clopidogrel After an Acute Myocardial Infarction. Clin. Pharmacol. Ther. 2011, 90, 561–567. [Google Scholar] [CrossRef]
- Reny, J.-L.; Combescure, C.; Daali, Y.; Fontana, P.; PON1 Meta-Analysis Group. Influence of the paraoxonase-1 Q192R genetic variant on clopidogrel responsiveness and recurrent cardiovascular events: A systematic review and meta-analysis. J. Thromb. Haemost. 2012, 10, 1242–1251. [Google Scholar] [CrossRef]
- Dansette, P.M.; Rosi, J.; Bertho, G.; Mansuy, D. Paraoxonase-1 and clopidogrel efficacy. Nat. Med. 2011, 17, 1040–1041. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Chen, M.; Zhang, L.; Zhao, Q. The impact of cytochrome 450 and Paraoxonase polymorphisms on clopidogrel resistance and major adverse cardiac events in coronary heart disease patients after percutaneous coronary intervention. BMC Pharmacol. Toxicol. 2020, 21, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, W.; Shi, X.; Xu, X.; Lin, Y. Both CYP2C19 and PON1 Q192R Genotypes Influence Platelet Response to Clopidogrel by Thrombelastography in Patients with Acute Coronary Syndrome. Cardiovasc. Ther. 2019, 2019, 3470145. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.-H.; Lao, H.-Y.; Wu, H.; Lai, W.-H.; Li, X.-X.; Yu, X.-Y.; Chen, J.-Y.; Zhong, S.-L. Association of PON1 genotype and haplotype with susceptibility to coronary artery disease and clinical outcomes in dual antiplatelet-treated Han Chinese patients. Eur. J. Clin. Pharmacol. 2013, 69, 1511–1519. [Google Scholar] [CrossRef]
- Lei, H.-P.; Yu, X.-Y.; Wu, H.; Kang, Y.-H.; Zhong, W.-P.; Cai, L.-Y.; Zhang, M.-Z.; Chen, J.-Y.; Mai, L.-P.; Ding, Q.-S.; et al. Effects of PON1 Gene Promoter DNA Methylation and Genetic Variations on the Clinical Outcomes of Dual Antiplatelet Therapy for Patients Undergoing Percutaneous Coronary Intervention. Clin. Pharmacokinet. 2017, 57, 817–829. [Google Scholar] [CrossRef] [PubMed]
- Deakin, S.; Guernier, S.; James, R.W. Pharmacogenetic interaction between paraoxonase-1 gene promoter polymorphism C-107T and statin. Pharmacogenet. Genom. 2007, 17, 451–457. [Google Scholar] [CrossRef] [PubMed]
- Arii, K.; Suehiro, T.; Ota, K.; Ikeda, Y.; Kumon, Y.; Osaki, F.; Inoue, M.; Inada, S.; Ogami, N.; Takata, H.; et al. Pitavastatin induces PON1 expression through p44/42 mitogen-activated protein kinase signaling cascade in Huh7 cells. Atherosclerosis 2009, 202, 439–445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arii, K.; Suehiro, T.; Ikeda, Y.; Kumon, Y.; Inoue, M.; Inada, S.; Takata, H.; Ishibashi, A.; Hashimoto, K.; Terada, Y. Role of protein kinase C in pitavastatin-induced human paraoxonase I expression in Huh7 cells. Metabolism 2010, 59, 1287–1293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ota, K.; Suehiro, T.; Arii, K.; Ikeda, Y.; Kumon, Y.; Osaki, F.; Hashimoto, K. Effect of pitavastatin on transactivation of human serum paraoxonase 1 gene. Metabolism 2005, 54, 142–150. [Google Scholar] [CrossRef]
- Sardo, M.A.; Campo, S.; Bonaiuto, M.; Bonaiuto, A.; Saitta, C.; Trimarchi, G.; Castaldo, M.; Bitto, A.; Cinquegrani, M.; Saitta, A. Antioxidant effect of atorvastatin is independent of PON1 gene T(–107)C, Q192R and L55M polymorphisms in hypercholesterolaemic patients. Curr. Med. Res. Opin. 2005, 21, 777–784. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PON Activity | Polymorphic Site (Functional Alias) | DbSNP Rs Number | References |
|---|---|---|---|
| Responsible for 22.8% variation in arylesterase activity; C allele associated with higher PON1 activity towards paraoxon, phenyl acetate and PON1 concentration; the highest PON1 activity and concentration is seen in CC genotype, medium in CT and the lowest in low-expressor genotype (TT) | -107/108 C/T; PON1 promoter | rs705379 | [31,43,54,57] |
| Influence on PON1 arylesterase activity | -162 A/G PON1 promoter | rs705381 | [54] |
| Influence on PON1 arylesterase activity | -907/909 C/G PON1 promoter | rs854572 | [54] |
| Influence on PON1 arylesterase and paraoxonase activity | PON1 introns | rs854567 | [31] |
| rs2299257 | |||
| rs3917486 | |||
| PON2 intron | rs2375005 | ||
| PON1 activity towards paraoxon higher in the presence 17899insTT/insTT and 17974 CT or TT PON1 activity towards phenyl acetate higher for 1799delTT/del TT | 17899insdelC/T PON1 intron 5 17974 C/T PON1 intron 5 | [58] | |
| Associated with PON3 concentration | -567 C/T PON3 promoter -665 A/G PON3 promoter -746 C/T PON3 promoter | [35] | |
| Differences in PON1 efficiency in atorvastatin δ -lactone hydrolysis | -4984 G/A PON3 promoter -4105 G/A PON3 promoter -1091 A/G PON3 promoter -746 C/T PON3 promoter | [73] |
| Clinical Relevance | Polymorphic Site (Functional Alias | DbSNP Rs Number | References |
|---|---|---|---|
| TT genotype more frequent in diabetic patients with CHD; CC genotype associated with lower risk CHD < 60 years old; T allele more frequent in women with CHD; C allele associated with less severe coronary artery stenoses; shorter cardiac events free survival after artery bypass—grafting (CABG) in TT and CT genotypes | -107/108 C/T; PON1 promoter | rs705379 | [87,88,90,91,92] |
| GG genotype independently associated with coronary artery disease (CAD) in northwest Indian Punjabi population | -162 A/G PON1 promoter | rs705381 | [93] |
| GG genotype associated with decreased risk of myocardial infarction 60 years old; and less severe coronary artery stenoses | -907/909 C/G PON1 promoter | rs854572 | [89,91] |
| G allele more frequent in women with CHD | -831 G/A PON1 promoter | rs854571 | [90] |
| Clinical Relevance | Polymorphic Site (Functional Alias) | DbSNP Rs Number | References |
|---|---|---|---|
| T allele increased the overall risk of arterial ischemic stroke in young adults (age <45); TT genotype two times increased risk of acute hemispheric stroke in the elderly (age >56) | -107/108 C/T; PON1 promoter | rs705379 | [103,104] |
| A/G polymorphism associated with risk of total and ischemic stroke | -162 A/G PON1 promoter | rs705381 | [106] |
| G/A variations were independent predictor of carotid IMT; A allele identified as protective factor | -824/-832 G/A PON1 promoter | rs854571 | [105,106] |
| CT or TT genotype via miR 616 associated with lower risk of ischemic stroke and lesser extent of carotid atherosclerosis | 3′UTR PON1 | rs3735590 | [60] |
| Clinical Relevance | Polymorphic Site (Functional Alias) | DbSNP Rs Number | References |
|---|---|---|---|
| TT genotype more frequent in diabetic patients with CHD; T allele associated with lower PON1 lactonase activity in neonates of gestational diabetes mellitus(GDM )mothers; TT and CT genotypes more frequent in nondiabetic patients with abnormal glucose control; C/T variations associated with insulin sensitivity; TT genotype associated with proatherogenic lipid profile, macrovascular disease and peripheral neuropathy in diabetics | -107/108 C/T; PON1 promoter | rs705379 | [87,88,112,116,117,118,78] |
| A allele associated with higher urinary albumin loss in diabetics | -162 A/G PON1 promoter | rs705381 | [119] |
| GG and GC genotypes associated with diabetes in northwest Indian Punjabi population; GG genotype was protective factor against diabetic retinopathy | -907/909 C/G PON1 promoter | rs854572 | [113] |
| A allele associated with higher urinary albumin loss in diabetics | -1074 A/G PON1 promoter | [119] |
| Clinical Relevance | Polymorphic Site (Functional Alias) | DbSNP Rs Number | References |
|---|---|---|---|
| Higher frequency of C allele in lupus erythematosus (LE) renal disease; allele C frequency higher in patients with non- progressive IgA nephropathy Allele T more frequent in patients with AD, TT genotype associated with unfavorable lipid profile in patients with AD of late onset; in meta – analysis allele A associated with higher risk of AD, protective role of GG genotype entry 1 | -107/108 C/T; PON1 promoter | rs705379 | [124,125,126,127,128,134] |
| Higher frequency of A allele in LE renal disease; Higher albumin loss in diabetic patients with allele A | -160/-162 A/G PON1 promoter | rs705381 | [124,119] |
| Higher frequency of G allele in LE renal disease; Higher albumin loss in diabetic patients with allele G | -1074 A/G | [124,119] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wysocka, A.; Zwolak, A. The Relevance of Noncoding DNA Variations of Paraoxonase Gene Cluster in Atherosclerosis-Related Diseases. Int. J. Mol. Sci. 2021, 22, 2137. https://doi.org/10.3390/ijms22042137
Wysocka A, Zwolak A. The Relevance of Noncoding DNA Variations of Paraoxonase Gene Cluster in Atherosclerosis-Related Diseases. International Journal of Molecular Sciences. 2021; 22(4):2137. https://doi.org/10.3390/ijms22042137
Chicago/Turabian StyleWysocka, Anna, and Agnieszka Zwolak. 2021. "The Relevance of Noncoding DNA Variations of Paraoxonase Gene Cluster in Atherosclerosis-Related Diseases" International Journal of Molecular Sciences 22, no. 4: 2137. https://doi.org/10.3390/ijms22042137
APA StyleWysocka, A., & Zwolak, A. (2021). The Relevance of Noncoding DNA Variations of Paraoxonase Gene Cluster in Atherosclerosis-Related Diseases. International Journal of Molecular Sciences, 22(4), 2137. https://doi.org/10.3390/ijms22042137