
Modulation of Aryl Hydrocarbon Receptor Expression Alleviated Neuropathic Pain in a Chronic Constriction Nerve Injury Animal Model



Abstract
:1. Introduction
2. Results
2.1. Modulation of AhR Expression Related to Nerve Degeneration and Regeneration
2.2. Omeprazole Interacts with the Molecular Surface Representation of the AhR Binding Site (Ser36)
2.3. Knockout of AhR Aggravates Neurobehavior and Effects Attenuated by AhR Agonist
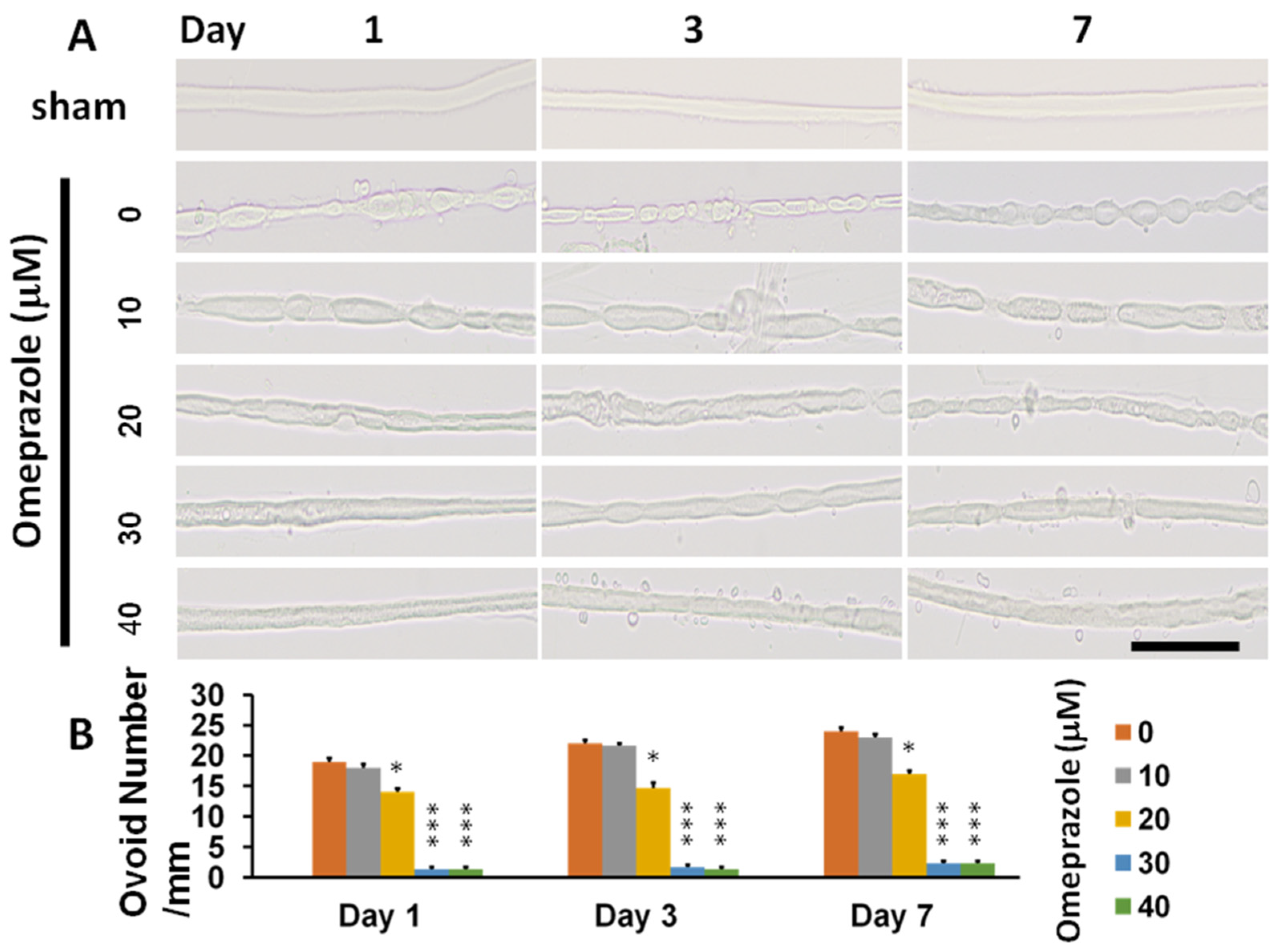
2.4. Histomorphology of Crushed Nerves from CCI and in AhR-Knockout Animals and the Attenuation Effect by AhR Agonist
2.5. Increased Pain Threshold in the Somatosensory System after CCI and in Ahr-Knockout Animals and the Attenuation Effect by AhR Agonist
3. Discussion
4. Materials and Methods
4.1. Sciatic Nerve Explants Culture from Mice
4.2. Animal Model
4.3. Mechanical Allodynia and Thermal Hyperalgesia
4.4. CatWalk Automated Quantitative Gait Analysis
4.5. Electrophysiological Study
4.6. Immunohistochemical Analyses
4.7. Molecular Docking Modeling
4.8. Statistical Analyses
5. Conclusions
6. Patents
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Baron, R.; Tölle, T.R. Assessment and diagnosis of neuropathic pain. Curr. Opin. Support. Palliat. Care 2008, 2, 1–8. [Google Scholar] [CrossRef]
- Treede, R.D.; Jensen, T.S.; Campbell, J.N.; Cruccu, G.; Dostrovsky, J.O.; Griffin, J.W.; Hansson, P.; Hughes, R.; Nurmikko, T.; Serra, J. Neuropathic pain: Redefinition and a grading system for clinical and research purposes. Neurology 2008, 70, 1630–1635. [Google Scholar] [CrossRef]
- Sacerdote, P.; Niada, S.; Franchi, S.; Arrigoni, E.; Rossi, A.; Yenagi, V.; de Girolamo, L.; Panerai, A.E.; Brini, A.T. Systemic administration of human adipose-derived stem cells reverts nociceptive hypersensitivity in an experimental model of neuropathy. Stem Cells Dev. 2013, 22, 1252–1263. [Google Scholar] [CrossRef]
- Siniscalco, D.; Rossi, F.; Maione, S. Molecular approaches for neuropathic pain treatment. Curr. Med. Chem. 2007, 14, 1783–1787. [Google Scholar] [CrossRef]
- Guyot, E.; Chevallier, A.; Barouki, R.; Coumoul, X. The AhR twist: Ligand-dependent AhR signaling and pharmaco-toxicological implications. Drug Discov. Today 2013, 18, 479–486. [Google Scholar] [CrossRef]
- Juricek, L.; Coumoul, X. The Aryl Hydrocarbon Receptor and the Nervous System. Int. J. Mol. Sci. 2018, 19, 2504. [Google Scholar] [CrossRef]
- Jain, S.; Maltepe, E.; Lu, M.M.; Simon, C.; Bradfield, C.A. Expression of ARNT, ARNT2, HIF1 alpha, HIF2 alpha and Ah receptor mRNAs in the developing mouse. Mech. Dev. 1998, 73, 117–123. [Google Scholar] [CrossRef]
- Kimura, E.; Tohyama, C. Embryonic and Postnatal Expression of Aryl Hydrocarbon Receptor mRNA in Mouse Brain. Front. Neuroanat. 2017, 11, 4. [Google Scholar] [CrossRef]
- Moran, T.B.; Brannick, K.E.; Raetzman, L.T. Aryl-hydrocarbon receptor activity modulates prolactin expression in the pituitary. Toxicol. Appl. Pharmacol. 2012, 265, 139–145. [Google Scholar] [CrossRef]
- Filbrandt, C.R.; Wu, Z.; Zlokovic, B.; Opanashuk, L.; Gasiewicz, T.A. Presence and functional activity of the aryl hydrocarbon receptor in isolated murine cerebral vascular endothelial cells and astrocytes. Neurotoxicology 2004, 25, 605–616. [Google Scholar] [CrossRef]
- Kubota, A.; Stegeman, J.J.; Woodin, B.R.; Iwanaga, T.; Harano, R.; Peterson, R.E.; Hiraga, T.; Teraoka, H. Role of zebrafish cytochrome P450 CYP1C genes in the reduced mesencephalic vein blood flow caused by activation of AHR2. Toxicol. Appl. Pharmacol. 2011, 253, 244–252. [Google Scholar] [CrossRef] [PubMed]
- Pravettoni, A.; Colciago, A.; Negri-Cesi, P.; Villa, S.; Celotti, F. Ontogenetic development, sexual differentiation, and effects of Aroclor 1254 exposure on expression of the arylhydrocarbon receptor and of the arylhydrocarbon receptor nuclear translocator in the rat hypothalamus. Reprod. Toxicol. 2005, 20, 521–530. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Chen, G.; Zhao, J.; Nie, X.; Wan, C.; Liu, J.; Duan, Z.; Xu, G. 2,3,7,8-Tetrachlorodibenzo-p-dioxin (TCDD) induces microglial nitric oxide production and subsequent rat primary cortical neuron apoptosis through p38/JNK MAPK pathway. Toxicology 2013, 312, 132–141. [Google Scholar] [CrossRef] [PubMed]
- Mukai, M.; Lin, T.M.; Peterson, R.E.; Cooke, P.S.; Tischkau, S.A. Behavioral rhythmicity of mice lacking AhR and attenuation of light-induced phase shift by 2,3,7,8-tetrachlorodibenzo-p-dioxin. J. Biol. Rhythm. 2008, 23, 200–210. [Google Scholar] [CrossRef]
- Petersen, S.L.; Curran, M.A.; Marconi, S.A.; Carpenter, C.D.; Lubbers, L.S.; McAbee, M.D. Distribution of mRNAs encoding the arylhydrocarbon receptor, arylhydrocarbon receptor nuclear translocator, and arylhydrocarbon receptor nuclear translocator-2 in the rat brain and brainstem. J. Comp. Neurol. 2000, 427, 428–439. [Google Scholar] [CrossRef]
- Cuartero, M.I.; Ballesteros, I.; de la Parra, J.; Harkin, A.L.; Abautret-Daly, A.; Sherwin, E.; Fernández-Salguero, P.; Corbí, A.L.; Lizasoain, I.; Moro, M.A. L-kynurenine/aryl hydrocarbon receptor pathway mediates brain damage after experimental stroke. Circulation 2014, 130, 2040–2051. [Google Scholar] [CrossRef]
- Wójtowicz, A.K.; Szychowski, K.A.; Wnuk, A.; Kajta, M. Dibutyl Phthalate (DBP)-Induced Apoptosis and Neurotoxicity are Mediated via the Aryl Hydrocarbon Receptor (AhR) but not by Estrogen Receptor Alpha (ERα), Estrogen Receptor Beta (ERβ), or Peroxisome Proliferator-Activated Receptor Gamma (PPARγ) in Mouse Cortical Neurons. Neurotoxic. Res. 2017, 31, 77–89. [Google Scholar]
- Xu, K.; Yang, Z.; Shi, R.; Luo, C.; Zhang, Z. Expression of aryl hydrocarbon receptor in rat brain lesions following traumatic brain injury. Diagn. Pathol. 2016, 11, 72. [Google Scholar] [CrossRef]
- Scholz, J.; Woolf, C.J. The neuropathic pain triad: Neurons, immune cells and glia. Nat. Neurosci. 2007, 10, 1361–1368. [Google Scholar] [CrossRef]
- Chiang, C.Y.; Liu, S.A.; Sheu, M.L.; Chen, F.C.; Chen, C.J.; Su, H.L.; Pan, H.C. Feasibility of Human Amniotic Fluid Derived Stem Cells in Alleviation of Neuropathic Pain in Chronic Constrictive Injury Nerve Model. PLoS ONE 2016, 11, e0159482. [Google Scholar] [CrossRef]
- Chiang, C.Y.; Sheu, M.L.; Cheng, F.C.; Chen, C.J.; Su, H.L.; Sheehan, J.; Pan, H.C. Comprehensive analysis of neurobehavior associated with histomorphological alterations in a chronic constrictive nerve injury model through use of the CatWalk XT system. J. Neurosurg. 2014, 120, 250–262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheu, M.L.; Chiang, C.Y.; Su, H.L.; Chen, C.J.; Sheehan, J.; Pan, H.C. Intrathecal Injection of Dual Zipper Kinase shRNA Alleviating the Neuropathic Pain in a Chronic Constrictive Nerve Injury Model. Int. J. Mol. Sci. 2018, 19, 2421. [Google Scholar] [CrossRef] [PubMed]
- Jin, U.H.; Lee, S.O.; Safe, S. Aryl hydrocarbon receptor (AHR)-active pharmaceuticals are selective AHR modulators in MDA-MB-468 and BT474 breast cancer cells. J. Pharmacol. Exp. Ther. 2012, 343, 333–341. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Sorrentino, C.; Denison, M.S.; Kolaja, K.; Fielden, M.R. Induction of cyp1a1 is a nonspecific biomarker of aryl hydrocarbon receptor activation: Results of large scale screening of pharmaceuticals and toxicants in vivo and in vitro. Mol. Pharmacol. 2007, 71, 1475–1486. [Google Scholar] [CrossRef]
- Shivanna, B.; Chu, C.; Welty, S.E.; Jiang, W.; Wang, L.; Couroucli, X.I.; Moorthy, B. Omeprazole attenuates hyperoxic injury in H441 cells via the aryl hydrocarbon receptor. Free Radic. Biol. Med. 2011, 51, 1910–1917. [Google Scholar] [CrossRef]
- Shivanna, B.; Jiang, W.; Wang, L.; Couroucli, X.I.; Moorthy, B. Omeprazole attenuates hyperoxic lung injury in mice via aryl hydrocarbon receptor activation and is associated with increased expression of cytochrome P4501A enzymes. J. Pharmacol. Exp. Ther. 2011, 339, 106–114. [Google Scholar] [CrossRef]
- Xing, H.; Chen, M.; Ling, J.; Tan, W.; Gu, J.G. TRPM8 mechanism of cold allodynia after chronic nerve injury. J. Neurosci. 2007, 27, 13680–13690. [Google Scholar] [CrossRef]
- Lai, D.W.; Lin, K.H.; Sheu, W.H.; Lee, M.R.; Chen, C.Y.; Lee, W.J.; Hung, Y.W.; Shen, C.C.; Chung, T.J.; Liu, S.H.; et al. TPL2 (Therapeutic Targeting Tumor Progression Locus-2)/ATF4 (Activating Transcription Factor-4)/SDF1α (Chemokine Stromal Cell-Derived Factor-α) Axis Suppresses Diabetic Retinopathy. Circ. Res. 2017, 121, e37–e52. [Google Scholar] [CrossRef]
- Lee, W.J.; Liu, S.H.; Chiang, C.K.; Lin, S.Y.; Liang, K.W.; Chen, C.H.; Tien, H.R.; Chen, P.H.; Wu, J.P.; Tsai, Y.C.; et al. Aryl Hydrocarbon Receptor Deficiency Attenuates Oxidative Stress-Related Mesangial Cell Activation and Macrophage Infiltration and Extracellular Matrix Accumulation in Diabetic Nephropathy. Antioxid. Redox Signal. 2016, 24, 217–231. [Google Scholar] [CrossRef]
- Chen, G.; Zhang, Y.Q.; Qadri, Y.J.; Serhan, C.N.; Ji, R.R. Microglia in Pain: Detrimental and Protective Roles in Pathogenesis and Resolution of Pain. Neuron 2018, 100, 1292–1311. [Google Scholar] [CrossRef]
- Wekerle, H. Brain inflammatory cascade controlled by gut-derived molecules. Nature 2018, 557, 642–643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Hua, D.; Wang, Q.; Yang, L.; Wang, X.; Luo, A.; Yang, C. The Role of Bacteria and Its Derived Metabolites in Chronic Pain and Depression: Recent Findings and Research Progress. Int. J. Neuropsychopharmacol. 2020, 23, 26–41. [Google Scholar] [CrossRef]
- Chanchal, S.K.; Mahajan, U.B.; Siddharth, S.; Reddy, N.; Goyal, S.N.; Patil, P.H.; Bommanahalli, B.P.; Kundu, C.N.; Patil, C.R.; Ojha, S. In vivo and in vitro protective effects of omeprazole against neuropathic pain. Sci. Rep. 2016, 6, 30007. [Google Scholar] [CrossRef] [PubMed]
- Pan, H.C.; Yang, D.Y.; Ou, Y.C.; Ho, S.P.; Cheng, F.C.; Chen, C.J. Neuroprotective effect of atorvastatin in an experimental model of nerve crush injury. Neurosurgery 2010, 67, 376–388, discussion 388–389. [Google Scholar] [CrossRef]
- Bennett, D.L. Neurotrophic factors: Important regulators of nociceptive function. Neuroscientist 2001, 7, 13–17. [Google Scholar] [CrossRef]
- Koltzenburg, M.; Bennett, D.L.; Shelton, D.L.; McMahon, S.B. Neutralization of endogenous NGF prevents the sensitization of nociceptors supplying inflamed skin. Eur. J. Neurosci. 1999, 11, 1698–1704. [Google Scholar] [CrossRef]
- Herzberg, U.; Eliav, E.; Dorsey, J.M.; Gracely, R.H.; Kopin, I.J. NGF involvement in pain induced by chronic constriction injury of the rat sciatic nerve. Neuroreport 1997, 8, 1613–1618. [Google Scholar] [CrossRef]
- Su, H.L.; Chiang, C.Y.; Lu, Z.H.; Cheng, F.C.; Chen, C.J.; Sheu, M.L.; Sheehan, J.; Pan, H.C. Late administration of high-frequency electrical stimulation increases nerve regeneration without aggravating neuropathic pain in a nerve crush injury. BMC Neurosci. 2018, 19, 37. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.Y.; Liu, M.G.; Yuan, D.L.; Wang, Y.; He, Y.; Wang, D.D.; Chen, X.F.; Zhang, F.K.; Li, H.; He, X.S.; et al. Nociception-induced spatial and temporal plasticity of synaptic connection and function in the hippocampal formation of rats: A multi-electrode array recording. Mol. Pain 2009, 5, 55. [Google Scholar]
- Liu, M.G.; Chen, J. Roles of the hippocampal formation in pain information processing. Neurosci. Bull. 2009, 25, 237–266. [Google Scholar] [CrossRef]
- Zelenka, M.; Schäfers, M.; Sommer, C. Intraneural injection of interleukin-1beta and tumor necrosis factor-alpha into rat sciatic nerve at physiological doses induces signs of neuropathic pain. Pain 2005, 116, 257–263. [Google Scholar] [CrossRef] [PubMed]
- Siniscalco, D.; Giordano, C.; Rossi, F.; Maione, S.; de Novellis, V. Role of neurotrophins in neuropathic pain. Curr. Neuropharmacol. 2011, 9, 523–529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vallejo, R.; Tilley, D.M.; Vogel, L.; Benyamin, R. The role of glia and the immune system in the development and maintenance of neuropathic pain. Pain Pract. 2010, 10, 167–184. [Google Scholar] [CrossRef]
- Echeverry, S.; Shi, X.Q.; Zhang, J. Characterization of cell proliferation in rat spinal cord following peripheral nerve injury and the relationship with neuropathic pain. Pain 2008, 135, 37–47. [Google Scholar] [CrossRef]
- Gu, N.; Peng, J.; Murugan, M.; Wang, X.; Eyo, U.B.; Sun, D.; Ren, Y.; DiCicco-Bloom, E.; Young, W.; Dong, H.; et al. Spinal Microgliosis Due to Resident Microglial Proliferation Is Required for Pain Hypersensitivity after Peripheral Nerve Injury. Cell Rep. 2016, 16, 605–614. [Google Scholar] [CrossRef] [PubMed]
- Guan, Z.; Kuhn, J.A.; Wang, X.; Colquitt, B.; Solorzano, C.; Vaman, S.; Guan, A.K.; Evans-Reinsch, Z.; Braz, J.; Devor, M.; et al. Injured sensory neuron-derived CSF1 induces microglial proliferation and DAP12-dependent pain. Nat. Neurosci. 2016, 19, 94–101. [Google Scholar] [CrossRef]
- Sorge, R.E.; Mapplebeck, J.C.; Rosen, S.; Beggs, S.; Taves, S.; Alexander, J.K.; Martin, L.J.; Austin, J.S.; Sotocinal, S.G.; Chen, D.; et al. Different immune cells mediate mechanical pain hypersensitivity in male and female mice. Nat. Neurosci. 2015, 18, 1081–1083. [Google Scholar] [CrossRef]
- Seeliger, D.; de Groot, B.L. Ligand docking and binding site analysis with PyMOL and Autodock/Vina. J. Comput. Aided Mol. Des. 2010, 24, 417–422. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Day | p Value | ||||||
|---|---|---|---|---|---|---|---|
| 0 | 7 | 14 | 21 | 28 | |||
| Stands (msec) | Wild type-sham | 245.2 ± 5.1 | 248.8 ± 6.1 | 253.5 ± 8.7 | 257.7 ± 3.9 | 255.2 ± 6.6 | <0.001 |
| Wild type-CCI | 244.2 ± 8.4 | 141.3 ± 11.1 | 162.2 ± 9.1 | 178.5 ± 11.5 | 186.8 ± 6.9 | ||
| AhR(−/−)-CCI | 244.7 ± 12.1 | 119.2 ± 5.4 | 125.7 ± 6.2 | 135.8 ± 6.4 | 142.2 ± 7.2 | ||
| Wild type-CCI- omeprazole | 246.2 ± 6.1 | 164.5 ± 8.4 | 183.1 ± 8.7 | 195.2 ± 3.9 | 216.8 ± 3.6 | ||
| Swings (msec) | Wild type-sham | 115.8 ± 4.8 | 118.1 ± 3.1 | 117.8 ± 4.5 | 117.8 ± 3.3 | 116.5 ± 3.2 | <0.001 |
| Wild type-CCI | 114.2 ± 5.4 | 210.5 ± 2.1 | 192.7 ± 5.1 | 180.3 ± 9.3, | 157.8 ± 2.9 | ||
| AhR(−/−)-CCI | 112.2 ± 4.8 | 319.8 ± 8.9 | 282 ± 8.1 | 238.7 ± 12.1 | 210.1 ± 9.7 | ||
| Wild type-CCI omeprazole | 115.1 ± 6.1 | 184 ± 6.6 | 162.2 ± 8.7 | 150.8 ± 9.9 | 135.3 ± 5.1 | ||
| Max Contact Max Intensity (Green Intensity) | Wild type-sham | 235.2 ± 5.4 | 238.8 ± 8.4 | 237.8 ± 2.8 | 238.5 ± 1.9 | 236.8 ± 3.5 | <0.001 |
| Wild type-CCI | 235 ± 3.1 | 188.5 ± 3.2 | 197.8 ± 3.3 | 240.5 ± 4.2 | 214.7 ± 2.1 | ||
| AhR(−/−)-CCI | 236.2 ± 5.4 | 175.8 ± 6.2 | 178.8 ± 4.2 | 187 ± 2.4 | 195.3 ± 2.9 | ||
| Wild type-CCI- omeprazole | 237.3 ± 3.8 | 198.2 ± 3.4 | 210.5 ± 1.2 | 218.8 ± 3.3 | 228 ± 3.2 | ||
| Printed area (cm2) | Wild type-sham | 0.315 ± 0.09 | 0.317 ± 0.08 | 0.318 ± 0.03 | 0.32 ± 0.03 | 0.319 ± 0.02 | <0.001 |
| Wild type-CCI | 0.319 ± 0.07 | 0.078 ± 0.01 | 0.138 ± 0.02 | 0.186 ± 0.02 | 0.247 ± 0.06 | ||
| AhR(−/−)-CCI | 0.318 ± 0.08 | 0.051 ± 0.002 | 0.094 ± 0.03 | 0.152 ± 0.03 | 0.194 ± 0.03 | ||
| Wild type-CCI- omeprazole | 0.320 ± 0.09 | 0.115 ± 0.01 | 0.180 ± 0.01 | 0.214 ± 0.04 | 0.282 ± 0.03 | ||
| Single Stance (msec) | Wild type-sham | 125.2 ± 4.1 | 124.3 ± 3.6 | 123.8 ± 4.2 | 123.3 ± 2.9 | 126 ± 2.2 | <0.001 |
| Wild type-CCI | 125.8 ± 4.2 | 71 ± 2.9 | 78.3 ± 1.2 | 85.9 ± 2.8 | 91.8 ± 3.9 | ||
| AhR(−/−)-CCI | 126.7 ± 3.6 | 64 ± 1.8 | 67.8 ± 1.9 | 73.2 ± 2.2 | 79.5 ± 2.1 | ||
| Wild type-CCI omeprazole | 126.2 ± 6.3 | 83.5 ± 3.9 | 89.3 ± 3.3 | 93.3 ± 3.1 | 104.8 ± 5.5 | ||
| Wild Type-Sham | Wild Type-CCI | AhR(−/−)-CCI | Wild Type-CCI-Omeprazole | p Value | |
|---|---|---|---|---|---|
| CD 68 | 3 ± 0.5 | 95.3 ± 3.3 | 143.7 ± 3.3 | 50.7 ± 2.9 | p < 0.001 |
| NF | 981 ± 16.5 | 426 ± 13.1 | 287 ± 19.7 | 604 ± 57.4 | p < 0.001 |
| NGF | 12.7 ± 1.2 | 399.3 ± 10.3 | 583 ± 16.6 | 180.3 ± 15.5 | p < 0.001 |
| TNF-α | 7 ± 1.2 | 778 ± 22.2 | 830.3 ± 20.6 | 183 ± 16.5 | p < 0.01 |
| Wild Type-Sham | Wild Type-CCI | AhR(−/−)-CCI | Wild Type-CCI- Omeprazole | p Value | |
|---|---|---|---|---|---|
| NGF | 19 ± 1.2 | 244.3 ± 14.5 | 429.3 ± 12.4 | 147.7 ± 14.8 | p < 0.001 |
| TNF-α | 9 ± 1.1 | 188.7 ± 5.5 | 477 ± 14.2 | 103.1 ± 4.1 | p < 0.001 |
| Synaptophysin | 12.1 ± 0.6 | 155.2 ± 6.9 | 241.1 ± 4.9 | 89.2 ± 6.1 | p < 0.001 |
| AhR | 102.7 ± 3.8 | 298.7 ± 6.4 | 14.1 ± 4.6 | 347.3 ± 14.5 | p < 0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sheu, M.-L.; Pan, L.-Y.; Sheehan, J.; Yang, M.-Y.; Pan, H.-C. Modulation of Aryl Hydrocarbon Receptor Expression Alleviated Neuropathic Pain in a Chronic Constriction Nerve Injury Animal Model. Int. J. Mol. Sci. 2022, 23, 11255. https://doi.org/10.3390/ijms231911255
Sheu M-L, Pan L-Y, Sheehan J, Yang M-Y, Pan H-C. Modulation of Aryl Hydrocarbon Receptor Expression Alleviated Neuropathic Pain in a Chronic Constriction Nerve Injury Animal Model. International Journal of Molecular Sciences. 2022; 23(19):11255. https://doi.org/10.3390/ijms231911255
Chicago/Turabian StyleSheu, Meei-Ling, Liang-Yi Pan, Jason Sheehan, Meng-Yin Yang, and Hung-Chuan Pan. 2022. "Modulation of Aryl Hydrocarbon Receptor Expression Alleviated Neuropathic Pain in a Chronic Constriction Nerve Injury Animal Model" International Journal of Molecular Sciences 23, no. 19: 11255. https://doi.org/10.3390/ijms231911255
APA StyleSheu, M. -L., Pan, L. -Y., Sheehan, J., Yang, M. -Y., & Pan, H. -C. (2022). Modulation of Aryl Hydrocarbon Receptor Expression Alleviated Neuropathic Pain in a Chronic Constriction Nerve Injury Animal Model. International Journal of Molecular Sciences, 23(19), 11255. https://doi.org/10.3390/ijms231911255