
Ultrastructural Characterization of PBMCs and Extracellular Vesicles in Multiple Sclerosis: A Pilot Study



Abstract
:1. Introduction
2. Results
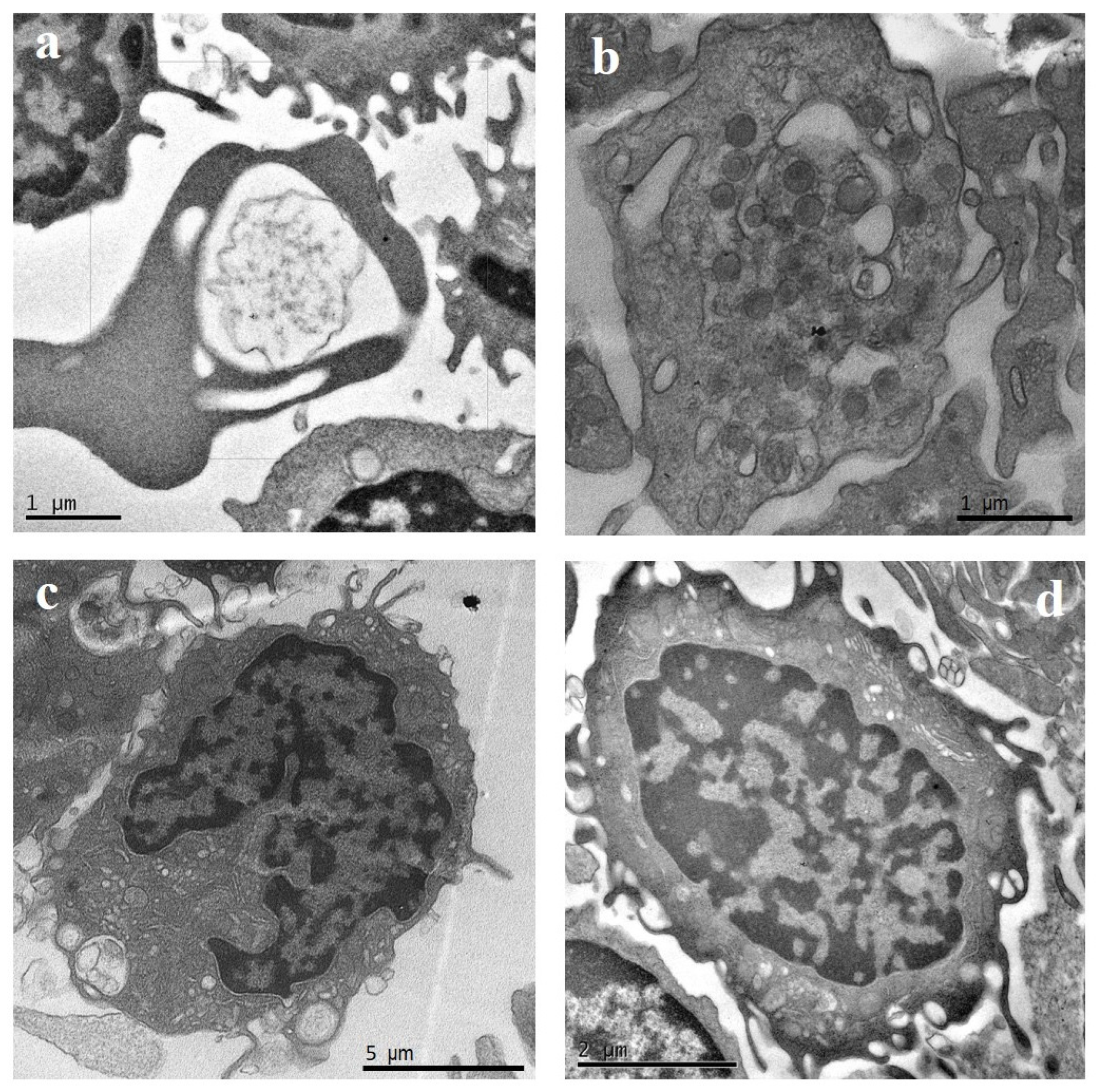
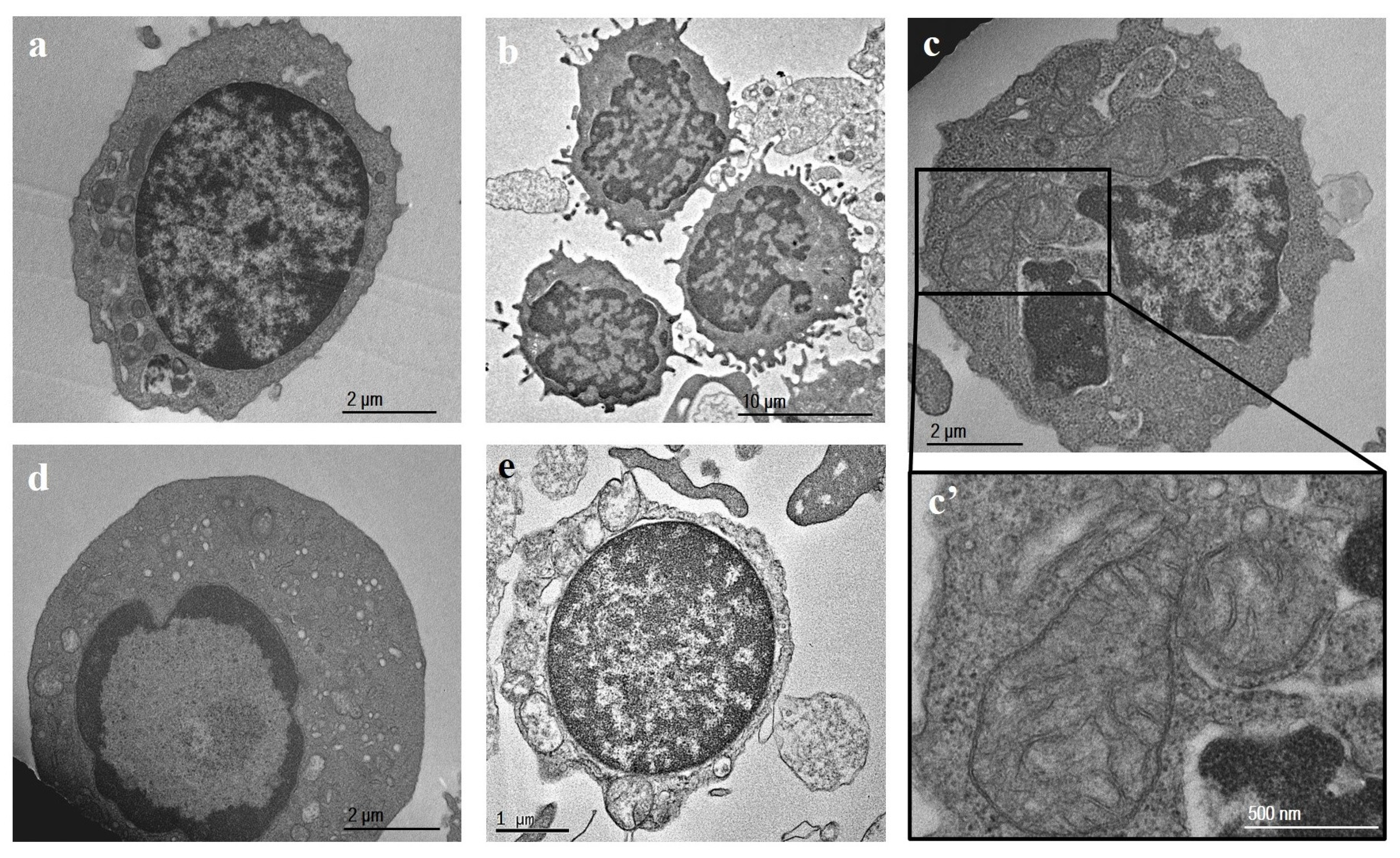
2.1. TEM
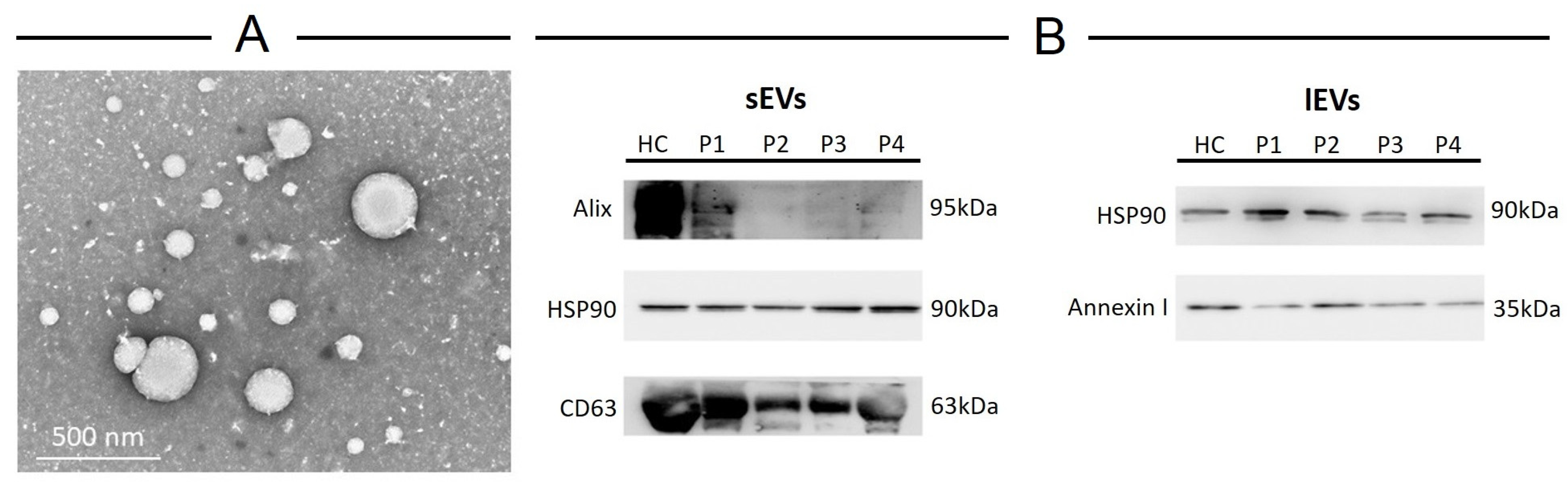
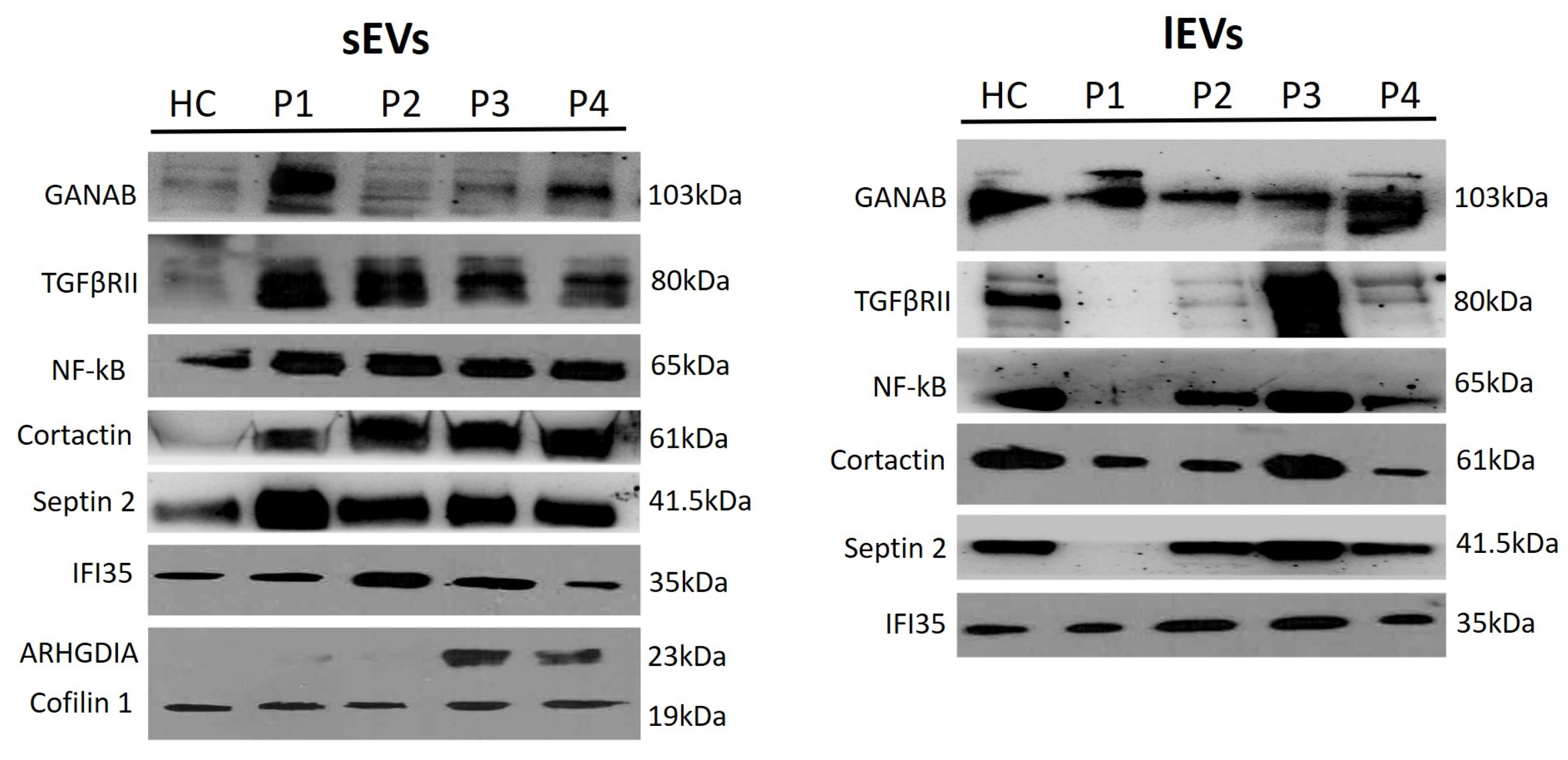
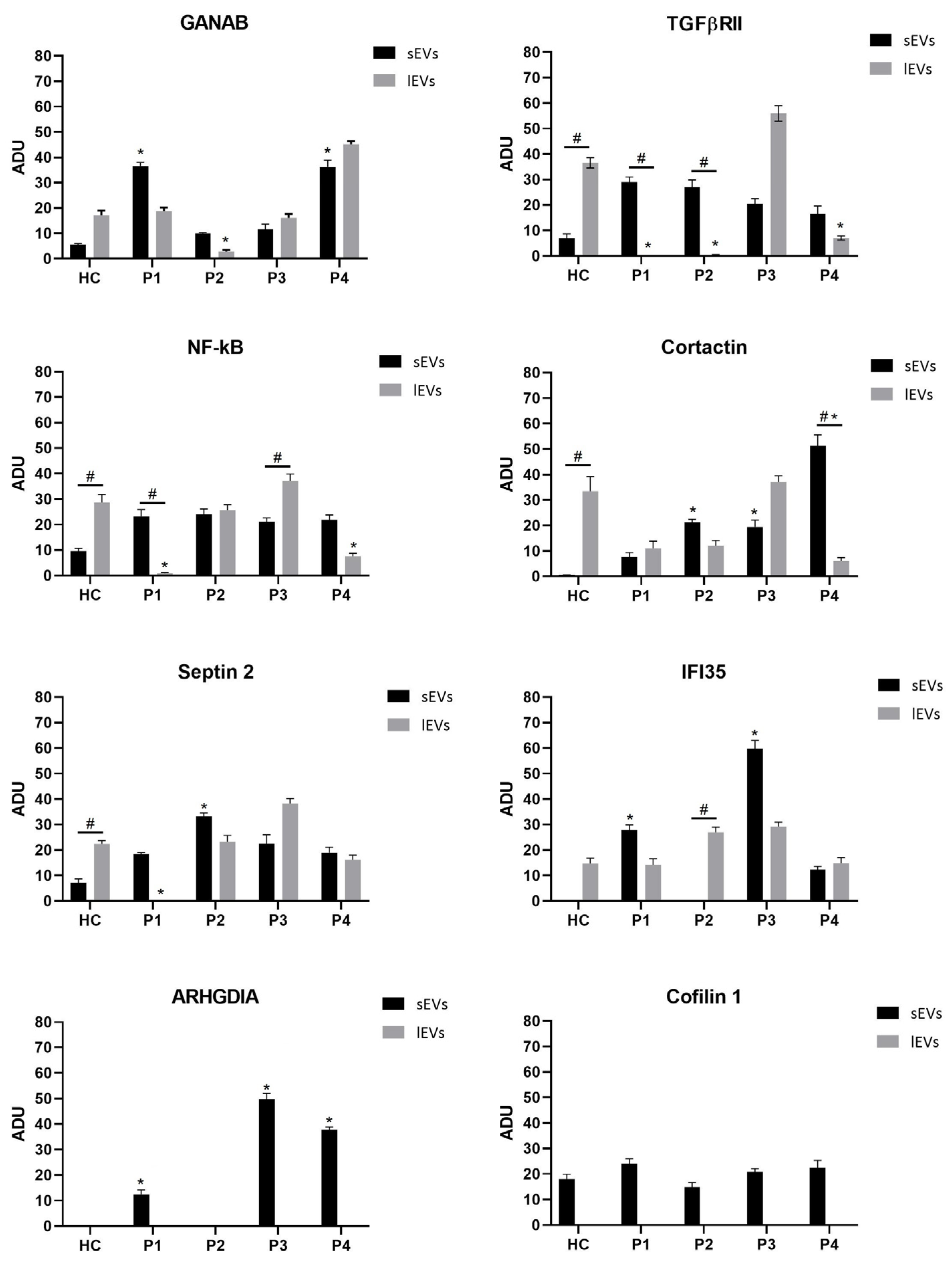
2.2. EV Characterization
3. Discussion
4. Materials and Methods
4.1. Study Design
4.2. Study Population
4.3. MRI Protocol
4.4. PBMC Isolation
4.5. EV Isolation
4.6. EV Characterization
4.7. TEM Protocol
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Palacio, P.L.; Pleet, M.L.; Reátegui, E.; Magaña, S.M. Emerging role of extracellular vesicles in multiple sclerosis: From cellular surrogates to pathogenic mediators and beyond. J. Neuroimmunol. 2023, 377, 578064. [Google Scholar] [CrossRef] [PubMed]
- Pistono, C.; Osera, C.; Cuccia, M.; Bergamaschi, R. Roles of Extracellular Vesicles in Multiple Sclerosis: From Pathogenesis to Potential Tools as Biomarkers and Therapeutics. Sclerosis 2023, 1, 91–112. [Google Scholar] [CrossRef]
- D’Anca, M.; Fenoglio, C.; Buccellato, F.R.; Visconte, C.; Galimberti, D.; Scarpini, E. Extracellular Vesicles in Multiple Sclerosis: Role in the Pathogenesis and Potential Usefulness as Biomarkers and Therapeutic Tools. Cells 2021, 10, 1733. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Tsunoda, I.; Fujinami, R.S. Inside-Out versus Outside-In models for virus induced demyelination: Axonal damage triggering demyelination. Springer Semin. Immunopathol. 2002, 24, 105–125. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Libbey, J.E.; McCoy, L.L.; Fujinami, R.S. Molecular mimicry in multiple sclerosis. Int. Rev. Neurobiol. 2007, 79, 127–147. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Mikita, J.; Dubourdieu-Cassagno, N.; Deloire, M.S.; Vekris, A.; Biran, M.; Raffard, G.; Brochet, B.; Canron, M.H.; Franconi, J.M.; Boiziau, C.; et al. Altered M1/M2 activation patterns of monocytes in severe relapsing experimental rat model of multiple sclerosis. Amelioration of clinical status by M2 activated monocyte administration. Mult. Scler. 2011, 17, 2–15. [Google Scholar] [CrossRef] [PubMed]
- Larochelle, C.; Alvarez, J.I.; Prat, A. How do immune cells overcome the blood-brain barrier in multiple sclerosis? FEBS Lett. 2011, 585, 3770–3780. [Google Scholar] [CrossRef] [PubMed]
- Trapp, B.D.; Bö, L.; Mörk, S.; Chang, A. Pathogenesis of tissue injury in MS lesions. J. Neuroimmunol. 1999, 98, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Bross, M.; Hackett, M.; Bernitsas, E. Approved and Emerging Disease Modifying Therapies on Neurodegeneration in Multiple Sclerosis. Int. J. Mol. Sci. 2020, 21, 4312. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Carata, E.; Muci, M.; Di Giulio, S.; Mariano, S.; Panzarini, E. Looking to the Future of the Role of Macrophages and Extracellular Vesicles in Neuroinflammation in ALS. Int. J. Mol. Sci. 2023, 24, 11251. [Google Scholar] [CrossRef]
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): A position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef] [PubMed]
- Di Bella, M.A. Overview and Update on Extracellular Vesicles: Considerations on Exosomes and Their Application in Modern Medicine. Biology 2022, 11, 804. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Picca, A.; Guerra, F.; Calvani, R.; Coelho-Junior, H.J.; Bucci, C.; Marzetti, E. Circulating extracellular vesicles: Friends and foes in neurodegeneration. Neural Regen. Res. 2022, 17, 534–542. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- De Masi, R.; Orlando, S. GANAB as a Novel Biomarker in Multiple Sclerosis: Correlation with Neuroinflammation and IFI35. Pharmaceuticals 2021, 14, 1195. [Google Scholar] [CrossRef] [PubMed]
- De Masi, R.; Orlando, S. GANAB and N-Glycans Substrates Are Relevant in Human Physiology, Polycystic Pathology and Multiple Sclerosis: A Review. Int. J. Mol. Sci. 2022, 23, 7373. [Google Scholar] [CrossRef] [PubMed]
- Field, E.J.; Raine, C.S. Experimental allergic encephalomyelitis. An electron microscopic study. Am. J. Pathol. 1966, 49, 537–553. [Google Scholar] [PubMed] [PubMed Central]
- Brenton, J.N.; Kammeyer, R.; Gluck, L.; Schreiner, T.; Makhani, N. Multiple Sclerosis in Children: Current and Emerging Concepts. Semin. Neurol. 2020, 40, 192–200. [Google Scholar] [CrossRef] [PubMed]
- Filippi, M.; Bar-Or, A.; Piehl, F.; Preziosa, P.; Solari, A.; Vukusic, S.; Rocca, M.A. Multiple sclerosis. Nat. Rev. Dis. Primers 2018, 4, 43. [Google Scholar] [CrossRef] [PubMed]
- García-Domínguez, J.M.; Maurino, J.; Martínez-Ginés, M.L.; Carmona, O.; Caminero, A.B.; Medrano, N.; Ruíz-Beato, E.; W-IMPACT Clinical Investigators. Economic burden of multiple sclerosis in a population with low physical disability. BMC Public Health 2019, 19, 609. [Google Scholar] [CrossRef]
- Barzegar, M.; Najdaghi, S.; Afshari-Safavi, A.; Nehzat, N.; Mirmosayyeb, O.; Shaygannejad, V. Early predictors of conversion to secondary progressive multiple sclerosis. Mult. Scler. Relat. Disord. 2021, 54, 103115. [Google Scholar] [CrossRef]
- Acquaviva, M.; Menon, R.; Di Dario, M.; Dalla Costa, G.; Romeo, M.; Sangalli, F.; Colombo, B.; Moiola, L.; Martinelli, V.; Comi, G.; et al. Inferring Multiple Sclerosis Stages from the Blood Transcriptome via Machine Learning. Cell Rep. Med. 2020, 1, 100053. [Google Scholar] [CrossRef] [PubMed]
- Dörr, J.; Paul, F. The transition from first-line to second-line therapy in multiple sclerosis. Curr. Treat. Options Neurol. 2015, 17, 354. [Google Scholar] [CrossRef] [PubMed]
- Hemmer, B.; Kerschensteiner, M.; Korn, T. Role of the innate and adaptive immune responses in the course of multiple sclerosis. Lancet Neurol. 2015, 14, 406–419. [Google Scholar] [CrossRef] [PubMed]
- Carstensen, M.; Christensen, T.; Stilund, M.; Møller, H.J.; Petersen, E.L.; Petersen, T. Activated monocytes and markers of inflammation in newly diagnosed multiple sclerosis. Immunol. Cell Biol. 2020, 98, 549–562. [Google Scholar] [CrossRef] [PubMed]
- Altan-Bonnet, G.; Mukherjee, R. Cytokine-mediated communication: A quantitative appraisal of immune complexity. Nat. Rev. Immunol. 2019, 19, 205–217. [Google Scholar] [CrossRef] [PubMed]
- Buzas, E.I. The roles of extracellular vesicles in the immune system. Nat. Rev. Immunol. 2023, 23, 236–250. [Google Scholar] [CrossRef]
- Jiang, Y.; Liu, X.; Ye, J.; Ma, Y.; Mao, J.; Feng, D.; Wang, X. Migrasomes, a new mode of intercellular communication. Cell Commun. Signal 2023, 21, 105. [Google Scholar] [CrossRef]
- Yarwood, R.; Hellicar, J.; Woodman, P.G.; Lowe, M. Membrane trafficking in health and disease. Dis. Model. Mech. 2020, 13, dmm043448. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Möckl, L. The Emerging Role of the Mammalian Glycocalyx in Functional Membrane Organization and Immune System Regulation. Front. Cell Dev. Biol. 2020, 8, 253. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Kuo, J.C.; Paszek, M.J. Glycocalyx Curving the Membrane: Forces Emerging from the Cell Exterior. Annu. Rev. Cell Dev. Biol. 2021, 37, 257–283. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- De Masi, R.; Orlando, S. IFI35 as a biomolecular marker of neuroinflammation and treatment response in multiple sclerosis. Life Sci. 2020, 259, 118233. [Google Scholar] [CrossRef] [PubMed]
- De Masi, R.; Orlando, S.; Bagordo, F.; Grassi, T. IFP35 Is a Relevant Factor in Innate Immunity, Multiple Sclerosis, and Other Chronic Inflammatory Diseases: A Review. Biology 2021, 10, 1325. [Google Scholar] [CrossRef] [PubMed]
- Du, S.; Guan, Y.; Xie, A.; Yan, Z.; Gao, S.; Li, W.; Rao, L.; Chen, X.; Chen, T. Extracellular vesicles: A rising star for therapeutics and drug delivery. Extracellular vesicles: A rising star for therapeutics and drug delivery. J. Nanobiotechnol. 2023, 21, 231. [Google Scholar] [CrossRef] [PubMed]
- De Masi, R.; Vergara, D.; Pasca, S.; Acierno, R.; Greco, M.; Spagnolo, L.; Blasi, E.; Sanapo, F.; Trianni, G.; Maffia, M. PBMCs protein expression profile in relapsing IFN-treated multiple sclerosis: A pilot study on relation to clinical findings and brain atrophy. J. Neuroimmunol. 2009, 210, 80–86. [Google Scholar] [CrossRef] [PubMed]
- Holman, D.W.; Klein, R.S.; Ransohoff, R.M. The blood-brain barrier, chemokines and multiple sclerosis. Biochim. Biophys. Acta 2011, 1812, 220–230. [Google Scholar] [CrossRef] [PubMed]
- Marchetti, L.; Engelhardt, B. Immune cell trafficking across the blood-brain barrier in the absence and presence of neuroinflammation. Vasc. Biol. 2020, 2, H1–H18. [Google Scholar] [CrossRef] [PubMed]
- Xiahou, Z.; Wang, X.; Shen, J.; Zhu, X.; Xu, F.; Hu, R.; Guo, D.; Li, H.; Tian, Y.; Liu, Y.; et al. NMI and IFP35 serve as proinflammatory DAMPs during cellular infection and injury. Nat. Commun. 2017, 8, 950. [Google Scholar] [CrossRef] [PubMed]
- De Masi, R.; Pasca, S.; Scarpello, R.; Idolo, A.; De Donno, A. The clinical potential of blood-proteomics in multiple sclerosis. BMC Neurol. 2013, 13, 45. [Google Scholar] [CrossRef] [PubMed]
- Cosen-Binker, L.I.; Kapus, A. Cortactin: The gray eminence of the cytoskeleton. Physiology 2006, 21, 352–361. [Google Scholar] [CrossRef]
- Schnoor, M.; Stradal, T.E.; Rottner, K. Cortactin: Cell functions of A multifaceted actin-binding protein. Trends Cell Biol. 2018, 28, 79–98. [Google Scholar] [CrossRef]
- Dudek, S.M.; Jacobson, J.R.; Chiang, E.T.; Birukov, K.G.; Wang, P.; Zhan, X.; Garcia, J.G. Pulmonary endothelial cell barrier enhancement by sphingosine 1-phosphate: Roles for cortactin and myosin light chain kinase. J. Biol. Chem. 2004, 279, 24692–24700. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, J.R.; Dudek, S.M.; Singleton, P.A.; Kolosova, I.A.; Verin, A.D.; Garcia, J.G. Endothelial cell barrier enhancement by ATP is mediated by the small GTPase Rac and cortactin. Am. J. Physiol. Lung Cell Mol. Physiol. 2006, 291, L289–L295. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Kowalski, J.R.; Yacono, P.; Bajmoczi, M.; Shaw, S.K.; Froio, R.M.; Golan, D.E.; Thomas, S.M.; Luscinskas, F.W. Endothelial cell cortactin coordinates intercellular adhesion molecule-1 clustering and actin cytoskeleton remodeling during polymorphonuclear leukocyte adhesion and transmigration. J. Immunol. 2006, 177, 6440–6449. [Google Scholar] [CrossRef] [PubMed]
- García Ponce, A.; Citalán Madrid, A.F.; Vargas Robles, H.; Chánez Paredes, S.; Nava, P.; Betanzos, A.; Zarbock, A.; Rottner, K.; Vestweber, D.; Schnoor, M. Loss of cortactin causes endothelial barrier dysfunction via disturbed adrenomedullin secretion and actomyosin contractility. Sci. Rep. 2016, 6, 29003. [Google Scholar] [CrossRef] [PubMed]
- Samus, M.; Li, Y.T.; Sorokin, L.; Rottner, K.; Vestweber, D. Actin-Binding Protein Cortactin Promotes Pathogenesis of Experimental Autoimmune Encephalomyelitis by Supporting Leukocyte Infiltration into the Central Nervous System. J. Neurosci. 2020, 40, 1389–1404. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Cooper, J.A. Junctional Localization of Septin 2 Is Required for Organization of Junctional Proteins in Static Endothelial Monolayers. Arterioscler. Thromb. Vasc. Biol. 2021, 41, 346–359. [Google Scholar] [CrossRef]
- Kim, J.; Cooper, J.A. Septins regulate junctional integrity of endothelial monolayers. Mol. Biol. Cell 2018, 29, 1693–1703. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Benoit, B.; Poüs, C.; Baillet, A. Septins as membrane influencers: Direct play or in association with other cytoskeleton partners. Front. Cell Dev. Biol. 2023, 11, 1112319. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Kanellos, G.; Frame, M.C. Cellular functions of the ADF/cofilin family at a glance. J. Cell Sci. 2016, 129, 3211–3218. [Google Scholar] [CrossRef]
- Alsegiani, A.; Shah, Z. The role of cofilin in age-related neuroinflammation. Neural Regen. Res. 2020, 15, 1451–1459. [Google Scholar]
- Sun, Y.; Liang, L.; Dong, M.; Li, C.; Liu, Z.; Gao, H. Cofilin 2 in serum as a novel biomarker for Alzheimer’s disease in Han Chinese. Front. Aging Neurosci. 2019, 11, 214. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Cichon, J.; Sun, C.; Chen, B.; Jiang, M.; Chen, X.A.; Sun, Y.; Wang, Y.; Chen, G. Cofilin aggregation blocks intracellular trafficking and induces synaptic loss in hippocampal neurons. J. Biol. Chem. 2012, 287, 3919–3929. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Alugba, G.; Urhi, A.; Olateju, I.V.; Onyemarin, H.; Uzzi, C.; Oshiba-Fowode, T.; Obomanu, E.; Popoola, H.A.; Okoronkwo, E.J.; Ukenenye, E.; et al. Renal diseases associated with multiple sclerosis: A narrative review. Medicine 2022, 101, e31959. [Google Scholar] [CrossRef] [PubMed]
- Zhao, B.; Chen, Y.G. Regulation of TGF-β Signal Transduction. Scientifica 2014, 2014, 874065. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Gutiérrez-Fernández, M.; de la Cuesta, F.; Tallón, A.; Cuesta, I.; Fernández-Fournier, M.; Laso-García, F.; Gómez-de Frutos, M.C.; Díez-Tejedor, E.; Otero-Ortega, L. Potential Roles of Extracellular Vesicles as Biomarkers and a Novel Treatment Approach in Multiple Sclerosis. Int. J. Mol. Sci. 2021, 22, 9011. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Manu, M.S.; Hohjoh, H.; Yamamura, T. Extracellular Vesicles as Pro- and Anti-inflammatory Mediators, Biomarkers and Potential Therapeutic Agents in Multiple Sclerosis. Aging Dis. 2021, 12, 1451–1461. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Thompson, A.J.; Banwell, B.L.; Barkhof, F.; Carroll, W.M.; Coetzee, T.; Comi, G.; Correale, J.; Fazekas, F.; Filippi, M.; Freedman, M.S.; et al. Diagnosis of multiple sclerosis: 2017 revisions of the McDonald criteria. Lancet Neurol. 2018, 17, 162–173. [Google Scholar] [CrossRef] [PubMed]
- Miller, D.H.; Barkhof, F.; Berry, I.; Kappos, L.; Scotti, G. Magnetic resonance imaging in monitoring the treatment of multiple sclerosis: Concerted action guidelines. J. Neurol. Neurosurg. Psychiatry 1991, 54, 683–688. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.; Xu, H.; Li, Y.; Wei, C.; Guo, R.; Wang, F.; Wu, Y.; Liu, J.; Jia, J.; Yan, J.; et al. A Modified Ficoll-Paque Gradient Method for Isolating Mononuclear Cells from the Peripheral and Umbilical Cord Blood of Humans for Biobanks and Clinical Laboratories. Biopreserv. Biobank. 2018, 16, 82–91. [Google Scholar] [CrossRef] [PubMed]
- Panzarini, E.; Tacconi, S.; Carata, E.; Mariano, S.; Tata, A.M.; Dini, L. Molecular Characterization of Temozolomide-Treated and Non Temozolomide-Treated Glioblastoma Cells Released Extracellular Vesicles and Their Role in the Macrophage Response. Int. J. Mol. Sci. 2020, 21, 8353. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| sEVs | lEVs | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| HC | P1 | P2 | P3 | P4 | HC | P1 | P2 | P3 | P4 | |
| GANAB | 5.51 (±0.28) | 36.58 (±1.6) | 10.04 (±0.47) | 11.62 (±0.5) | 36.26 (±1.77) | 17.14 (±0.86) | 18.74 (±0.98) | 2.81 (±0.18) | 16.07 (±0.73) | 45.24 (±1.99) |
| TGFβRII | 6.91 (±0.27) | 29.10 (±1.44) | 26.97 (±1.36) | 20.49 (±0.98) | 16.54 (±0.79) | 36.62 (±1.93) | N.D. | 0.35 (±0.01) | 55.96 (±2.8) | 7.06 (±0.29) |
| NF-κB | 9.57 (±0.47) | 23.29 (±1.1) | 24.10 (±1.2) | 21.13 (±0.99) | 21.90 (±1.05) | 28.60 (±1.39) | 0.88 (±0.04) | 25.66 (±1.3) | 37.18 (±1.91) | 7.67 (±0.38) |
| Cortactin | 0.54 (±0.03) | 8.13 (±0.39) | 21.17 (±1.01) | 19.41 (±1.01) | 51.24 (±2.6) | 33.50 (±1.77) | 11.06 (±0.53) | 12.15 (±0.56) | 37.19 (±1.85) | 6.1 (±0.28) |
| Septin 2 | 7.13 (±0.36) | 18.4 (±0.94) | 33.17 (±1.53) | 22.36 (±1.23) | 18.94 (±0.97) | 22.38 (±0.99) | N.D. | 23.22 (±1.13) | 38.23 (±1.91) | 16.18 (±0.76) |
| IFI35 | 0.04 (±0.01) | 27.76 (±1.38) | 0.05 (±0.01) | 59.81 (±2.88) | 12.34 (±0.77) | 14.79 (±0.7) | 14.17 (±0.72) | 26.98 (±1.32) | 29.2 (±1.52) | 14.86 (±0.7) |
| ARHGDIA | N.D. | 12.41 (±0.65) | N.D. | 57.92 (±2.76) | 40.84 (±2.1) | N.D. | N.D. | N.D. | N.D. | N.D. |
| Cofilin 1 | 17.95 (±0.87) | 24.01 (±1.1) | 14.74 (±0.77) | 20.81 (±0.96) | 22.49 (±1.15) | N.D. | N.D. | N.D. | N.D. | N.D. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Masi, R.; Orlando, S.; Carata, E.; Panzarini, E. Ultrastructural Characterization of PBMCs and Extracellular Vesicles in Multiple Sclerosis: A Pilot Study. Int. J. Mol. Sci. 2024, 25, 6867. https://doi.org/10.3390/ijms25136867
De Masi R, Orlando S, Carata E, Panzarini E. Ultrastructural Characterization of PBMCs and Extracellular Vesicles in Multiple Sclerosis: A Pilot Study. International Journal of Molecular Sciences. 2024; 25(13):6867. https://doi.org/10.3390/ijms25136867
Chicago/Turabian StyleDe Masi, Roberto, Stefania Orlando, Elisabetta Carata, and Elisa Panzarini. 2024. "Ultrastructural Characterization of PBMCs and Extracellular Vesicles in Multiple Sclerosis: A Pilot Study" International Journal of Molecular Sciences 25, no. 13: 6867. https://doi.org/10.3390/ijms25136867