

Understanding Proton Magnetic Resonance Spectroscopy Neurochemical Changes Using Alzheimer’s Disease Biofluid, PET, Postmortem Pathology Biomarkers, and APOE Genotype

Abstract
:1. Introduction
2. Commonly Studied 1H MRS Metabolites in AD
2.1. NAA
2.2. mIns

{kind=link}
{kind=link}
| References | Cohort | Magnet Field Strength and Acquisition Parameters | Voxel Locations and Size | Key Findings |
|---|---|---|---|---|
| [58] | CU (n = 30) | 7T, TR = 644, MRSI, FIDLOVS | Posterior cingulate gyrus and precuneus | ↑ GABA and ↑ Glu were associated with ↑ Aβ burden on PET (PiB) with a positive effect modification by APOE e4 allele. |
| [59] | AD (11), MCI (8), CU (n = 26) | 3T, TR/TE = 2000/30 ms, MRSI, PRESS | Posterior cingulate gyrus, dorsolateral prefrontal cortex | ↓Glu/tCr was associated with ↑ tau load on PET with florzolatau in the posterior cingulate gyrus of AD dementia patients. ↑ plasma NfL was associated with MRS metabolites (↓ tNAA/tCr and ↓ Glu/tCr) in the right dorsolateral prefrontal cortex of patients with AD dementia. |
| [60] | CU (Aβ–and Aβ+) (n = 338), MCI (Aβ+)(n = 90) | 3T, TR/TE = 2000/30 ms, single voxel, PRESS | Posterior cingulate cortex/precuneus region | ↑ mIns/tCr ratio in the posterior cingulate gyrus was associated with ↑ posterior cingulate gyrus and neocortical meta-ROI Aβ (flutemetamol) and tau (RO948) load on PET only in APOE e4 allele carriers. ↑ plasma GFAP was associated with ↑ mIns/tCr (posterior cingulate gyrus) only in APOE e4 allele carriers. |
| [61] | CU women: CSF-Aβ-negative (n = 71); CU-Aβ-positive women (n = 37); MCI (CSF-Aβ-positive) women (n = 12) | 3T, TR/TE = 2000/20 ms; TR/TE = 2000/68 ms/; single voxel; PRESS and MEGA-PRESS | Medial frontal cortex | ↑ Glx, ↓ GABA, and ↑ mIns/tCr ratio in MCI compared to CU CSF-Aβ42-negative and -positive participants. ↑ Age was associated with ↓ levels of GABA in CU and MCI groups. |
| [62] | CU (A−T−N−) (37); early AD (A+T+N−) (n = 16); late AD (A+T+N+) (n = 15) | 3T, TR/TE = 2000/32 ms; single voxel; PRESS | Posterior cingulate cortex/precuneus region | ↓ NAA/Cr in early AD (A+T+N−) and late AD (A+T+N+) compared to controls (A−T−N−; A+T−N−). ↑ mIns/Cr in late AD compared to controls. ↓ NAA/Cr correlated with ↑ global Aβ load (PIB) and tau load (flortaucipir) on PET in whole cohort. |
| [63] | CU (n = 40) | 3T, TR/TE= 3000/30 ms, single voxel; sLASER | Posterior cingulate gyrus (automated VOI prescription) | ↑ Tau PET (flortaucipir) in posterior cingulate gyrus correlated with ↓ NAA/tCr and ↓ Glu/tCr. |
| [64] | CSF Aβ42 positive (n = 111); CSF Aβ42 negative (n = 174); | 3T, TR/TE= 3000/30 ms, single voxel; PRESS | Posterior cingulate cortex/precuneus region | Visit 2 (~2.3 years after baseline): ↑ Cho/Cr, ↑ mIns/Cr, ↓ NAA/Cr, and ↓ NAA/mI in CSF-Aβ-positive compared to CSF-Aβ-negative cases. Visit 3 (~4 years after baseline): ↑mIns/Cr, ↓ NAA/Cr, and ↓ NAA/mI in CSF-Aβ-positive compared to CSF-Aβ-negative cases. CSF Aβ positivity at baseline was associated with ↑ mIns/Cr and ↓ NAA/mIns ↑ Rate of change in the MCI Aβ positive for mIns/Cr and NAA/mIns compared to MCI Aβ negative. |
| [65] | CU younger controls (<60 years) (n = 27); CU older controls (>60 years) (n = 27); AD (>60 years) (n = 25) | 3T, TR/TE = 1600/(31–229) ms ms, single voxel, 2D J-PRESS | Posterior cingulate cortex/precuneus region | ↑ mIns associated with ↑CSF tau and ↑ CSF p-Tau 181; ↑ GABA associated with ↑CSF p-Tau 181p in AD dementia group. |
| [66] | Two cohorts: younger age (n = 30) (20–40 years); CU (n = 151): older individuals (60–85 years). | 3T, TR/TE = 4000/8.5 ms, single voxel, SPECIAL | Posterior cingulate cortex/precuneus region | ↑ mIns, ↑ Cr, ↑ mIns/NAA, ↓ GSH, and ↓ Glu in older participants compared to younger participants. |
| [67] | CU (n = 289) | 1.5T, TR/TE = 2000/25 ms, single voxel, PRESS | Posterior cingulate gyrus | ↑ mIns/Cr ratio in participants with two copies of APOE e4 allele compared with participants with non-carriers. ↓ The NAA/mIns ratio in participants (APOE e4/e4) compared with those who were heterozygous for the APOE e4 allele and non-carriers. |
| [68]. | CU (n = 15) | 3T, TR/TE = 1500/68 ms, single voxel, J-edited spin echo difference method | Posterior cingulate cortex/precuneus region | ↓ GSH was associated with↑ the temporal and parietal Aβ load on PET with PiB. |
| [69] | aMCI (n = 14); CU (n = 32) | 3T, TR/TE= 3000/30 ms, single voxel, sLASER | Posterior cingulate gyrus | ↑ Global cortical Aβ load (PiB) on PET correlated with ↓ Glu/mIns ratio in the entire cohort. |
| [70] | CU older adults (n = 594) c | 3T, TR/TE= 2000/30 ms, single voxel, PRESS | Posterior cingulate gyrus | ↓ NAA/mIns and ↑ mIns/Cr at baseline were associated with ↑ rate of Aβ deposition on serial PIB PET. |
| [71] | CU CSF Aβ42 negative (n = 156); CU CSF Aβ42 positive (n = 49), MCI CSF Aβ42 positive (n = 88) | 3T, TR/TE= 2000/30 ms, single voxel, PRESS | Posterior cingulate/precuneus | ↑ mIns/Cr, ↑ Cho/Cr, and ↓ NAA/Cr in MCI (CSF Aβ42 positive) compared to CU (CSF Aβ42 negative). ↑ mIns/Cr in CU (CSF Aβ42 positive) compared to CU (CSF Aβ42 negative). ↑ mIns/Cr in APOE e4 allele carrier CU (CSF Aβ42 negative) compared to non-e4-carrier CU (CSF Aβ42 negative). ↑ mIns/Cr and ↑ Cho/Cr were associated with ↑ Aβ deposition on PET (flutemetamol) in amyloid-positive (on PET) cognitively unimpaired participants. ↑ mIns/Cr was associated with ↑ Aβ deposition on PET (flutemetamol) and in CSF-Aβ42-positive cognitively unimpaired participants. |
| [72] | CU (n = 16), aMCI (n = 11) | 3T; TR/TE = 2000/32ms, single voxel, 2D-PRESS | Bilateral hippocampi | No difference in mIns/Cr between APOE e4 allele carriers and non-carriers |
| [73] | CU (n = 21); aMCI (n = 15) | 3T, TR/TE= 3000/68 ms, single voxel, MEGA-PRESS | Posterior cingulate gyrus | ↓ NAA was lower in Aβ-positive subjects compared to Aβ-negative (PiB PET) subjects. ↓ NAA was in APOE e4 allele carriers compared to non-carriers. |
| [74] | APOE e4 allele non-carriers (n = 89); APOE e4 allele carriers (n = 23) | 3T, TR/TE= 1600/30 ms, single voxel, PRESS | Posterior cingulate gyrus | ↑ Cho/Cr and ↑ mIns/Cr increase with age in APOE e4 allele carriers. ↑ Cho/Cr ratio APOE e4 carriers compared to non-carriers. |
| [29] | No to low likelihood of AD (n = 17); intermediate to high likelihood of AD (n = 24) | 3T, TR/TE= 2000/30 ms, single voxel, PRESS | Posterior cingulate gyrus | ↓ NAA/Cr and NAA/mIns were associated with ↓synaptic integrity and ↑higher p-tau pathology. ↑ Aβ burden was associated with ↑ mIns/Cr and ↓ NAA/mIns. ↑ GFAP-positive astrocytic burden showed a trend of association with decreased NAA/Cr and NAA/mIns. |
| [75] | CU (n = 17); AD (n = 19) | 3T, TR/TE= 2000/30 ms, single voxel, PRESS | Hippocampus, posterior cingulate gyrus, and right parietal gyrus | ↓ NAA/Cr (hippocampus) was correlated with ↓ CSF Aβ42. ↓ NAA/Cr (parietal gyrus) was correlated with ↑ CSF p-tau. ↑ mIns/Cr (posterior cingulate gyrus) was correlated with ↑ t-tau. |
| [76] | All subjects (n = 109); AD dementia (n = 40); non-AD dementia, (n = 14); MCI of AD type (n = 29) MCI of non-AD type (n = 26) | 1.5T, TR/TE= 2000/272, single voxel, PRESS | Medial temporal lobe | ↓ NAA was correlated with ↓ CSF Aβ42 in patient with AD dementia. |
| [77] | CU (n = 311) | 1.5 T, 2000/30 ms, single voxel, PRESS | Posterior cingulate gyrus | ↑ mIns/Cr and ↑ Cho/Cr were associated with ↑ Aβ load on PET (PIB). |
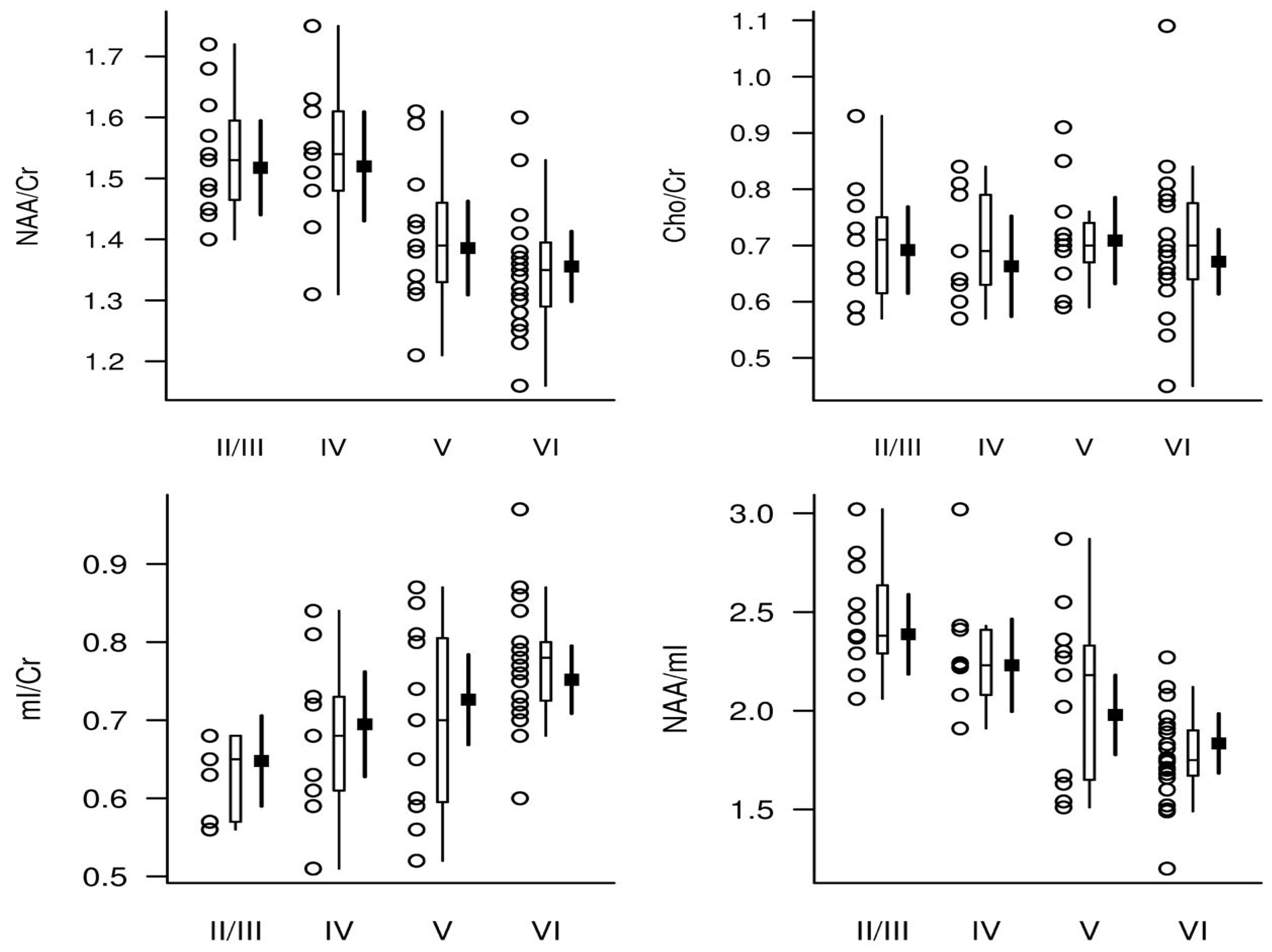
| [78] | Low AD likelihood (n = 11); intermediate AD likelihood (n = 9); high AD likelihood (n = 34) | 1.5 T/2000/30 ms, single voxel, PRESS | Posterior cingulate gyrus | ↓ NAA/Cr, ↑ mIns/Cr, and ↓ NAA/mIns in postmortem frequent neuritic plaque group compared to neuritic sparse plaque group. ↓ NAA/Cr in frequent neuritic plaque group compared to neuritic moderate plaque group. ↑ mIns/Cr and ↓ NAA/mIns in neuritic moderate plaque group compared to neuritic sparse plaque group. ↓ NAA/Cr, ↑ mIns/Cr, and ↓ NAA/mIns in high-likelihood AD group compared to low-likelihood AD group. ↑ mIns/Cr in high-likelihood AD group compared to intermediate-likelihood AD group. ↓ NAA/Cr, ↑ mI/Cr, and ↓ NAA/mI ratios were associated with higher Braak NFT stage, higher neuritic plaque score, and greater likelihood of AD. |
| [79] | CU (n = 61); patient group (MCI + AD dementia (n = 46) | 1.5 T/2000/30 ms, single voxel, PRESS | Posterior cingulate/precuneus | No differences were noted on 1H-MRS metabolite ratios (NAA/Cr, mIns/Cr, NAA/mIns) across APOE e4 carriers and non-carriers. |
| [50] | CU (63); MCI (21); AD dementia (21) | 1.5 T/2000/30 or 135 ms, single voxel, PRESS | Posterior cingulate gyrus; medial occipital; left superior temporal lobe | ↑ NAA/Cr ratios (medial occipital) in patients with AD dementia correlated with APOE e4 carrier status. |
| [80] | Postmortem brain with AD pathology (49); non-demented control (5) | In vitro, 11.7 T, perchloric acid extracts | Autopsy brain samples from various brain regions | ↑ mIns, ↑ GPC, and ↓ Glu in APOE e3/e3 samples from AD dementia patients compared to samples from normal control brain samples. ↓ NAA in APOE e3/e3 and APOE e4/e4 AD samples from AD dementia patients compared to samples from normal control brains (APOE e3/e3). |
2.3. Cho
2.4. Glu, Gln, Glx
2.5. GABA
2.6. GSH
2.7. Lactate
2.8. Cr
3. Association of 1H MRS Metabolites with Postmortem Neuropathology
4. Association of 1H MRS Metabolites with Tau and Amyloid PET
4.1. NAA
4.2. mIns
4.3. Cho
4.4. Glx and Glu
4.5. GABA
4.6. GSH
5. Association of 1H MRS Metabolites with Biofluid Biomarkers
5.1. NAA
5.2. mIns
5.3. Cho
5.4. Glu
5.5. GABA
6. Influence of APOE ε4 Allele on 1H MRS Metabolites
7. Future Directions
Author Contributions
Funding
Conflicts of Interest
References
- 2024 Alzheimer’s disease facts and figures. Alzheimer’s Dement. 2024, 20, 3708–3821. [CrossRef]
- Nichols, E.; Szoeke, C.E.I.; Vollset, S.E.; Abbasi, N.; Abd-Allah, F.; Abdela, J.; Aichour, M.T.E.; Akinyemi, R.O.; Alahdab, F.; Asgedom, S.W.; et al. Global, regional, and national burden of Alzheimer’s disease and other dementias, 1990–2016: A systematic analysis for the Global Burden of Disease Study 2016. Lancet Neurol. 2019, 18, 88–106. [Google Scholar] [CrossRef]
- Hay, S.I.; Abajobir, A.A.; Abate, K.H.; Abbafati, C.; Abbas, K.M.; Abd-Allah, F.; Abdulkader, R.S.; Abdulle, A.M.; Abebo, T.A.; Abera, S.F.; et al. Global, regional, and national disability-adjusted life-years (DALYs) for 333 diseases and injuries and healthy life expectancy (HALE) for 195 countries and territories, 1990–2016: A systematic analysis for the Global Burden of Disease Study 2016. Lancet 2017, 390, 1260–1344. [Google Scholar] [CrossRef]
- Li, R.; Qi, J.; Yang, Y.; Wu, Y.; Yin, P.; Zhou, M.; Qian, Z.; LeBaige, M.H.; McMillin, S.E.; Guo, H.; et al. Disease Burden and Attributable Risk Factors of Alzheimer’s Disease and Dementia in China from 1990 to 2019. J. Prev. Alzheimers Dis. 2022, 9, 306–314. [Google Scholar] [CrossRef]
- van Dyck, C.H.; Swanson, C.J.; Aisen, P.; Bateman, R.J.; Chen, C.; Gee, M.; Kanekiyo, M.; Li, D.; Reyderman, L.; Cohen, S.; et al. Lecanemab in Early Alzheimer’s Disease. N. Engl. J. Med. 2023, 388, 9–21. [Google Scholar] [CrossRef]
- Jack, C.R.; Knopman, D.S.; Jagust, W.J.; Petersen, R.C.; Weiner, M.W.; Aisen, P.S.; Shaw, L.M.; Vemuri, P.; Wiste, H.J.; Weigand, S.D.; et al. Tracking pathophysiological processes in Alzheimer’s disease: An updated hypothetical model of dynamic biomarkers. Lancet Neurol. 2013, 12, 207–216. [Google Scholar] [CrossRef]
- Bateman, R.J.; Xiong, C.; Benzinger, T.L.; Fagan, A.M.; Goate, A.; Fox, N.C.; Marcus, D.S.; Cairns, N.J.; Xie, X.; Blazey, T.M.; et al. Clinical and biomarker changes in dominantly inherited Alzheimer’s disease. N. Engl. J. Med. 2012, 367, 795–804. [Google Scholar] [CrossRef]
- Villemagne, V.L.; Burnham, S.; Bourgeat, P.; Brown, B.; Ellis, K.A.; Salvado, O.; Szoeke, C.; Macaulay, S.L.; Martins, R.; Maruff, P.; et al. Amyloid β deposition, neurodegeneration, and cognitive decline in sporadic Alzheimer’s disease: A prospective cohort study. Lancet Neurol. 2013, 12, 357–367. [Google Scholar] [CrossRef]
- Jack, C.R., Jr.; Bennett, D.A.; Blennow, K.; Carrillo, M.C.; Dunn, B.; Haeberlein, S.B.; Holtzman, D.M.; Jagust, W.; Jessen, F.; Karlawish, J.; et al. NIA-AA Research Framework: Toward a biological definition of Alzheimer’s disease. Alzheimer’s Dement. 2018, 14, 535–562. [Google Scholar] [CrossRef]
- Jack, C.R., Jr.; Andrews, J.S.; Beach, T.G.; Buracchio, T.; Dunn, B.; Graf, A.; Hansson, O.; Ho, C.; Jagust, W.; McDade, E.; et al. Revised criteria for diagnosis and staging of Alzheimer’s disease: Alzheimer’s Association Workgroup. Alzheimer’s Dement. 2024, 20, 5143–5169. [Google Scholar] [CrossRef]
- Lippa, C.F.; Fujiwara, H.; Mann, D.M.; Giasson, B.; Baba, M.; Schmidt, M.L.; Nee, L.E.; O’Connell, B.; Pollen, D.A.; St George-Hyslop, P.; et al. Lewy bodies contain altered alpha-synuclein in brains of many familial Alzheimer’s disease patients with mutations in presenilin and amyloid precursor protein genes. Am. J. Pathol. 1998, 153, 1365–1370. [Google Scholar] [CrossRef]
- Hamilton, R.L. Lewy bodies in Alzheimer’s disease: A neuropathological review of 145 cases using alpha-synuclein immunohistochemistry. Brain Pathol. 2000, 10, 378–384. [Google Scholar] [CrossRef]
- Twohig, D.; Nielsen, H.M. α-synuclein in the pathophysiology of Alzheimer’s disease. Mol. Neurodegener. 2019, 14, 23. [Google Scholar] [CrossRef]
- Shah, D.; Gsell, W.; Wahis, J.; Luckett, E.S.; Jamoulle, T.; Vermaercke, B.; Preman, P.; Moechars, D.; Hendrickx, V.; Jaspers, T.; et al. Astrocyte calcium dysfunction causes early network hyperactivity in Alzheimer’s disease. Cell Rep. 2022, 40, 111280. [Google Scholar] [CrossRef]
- Huang, S.; Xu, B.; Liu, Y. Calcium promotes α-synuclein liquid-liquid phase separation to accelerate amyloid aggregation. Biochem. Biophys. Res. Commun. 2022, 603, 13–20. [Google Scholar] [CrossRef]
- Carapeto, A.P.; Marcuello, C.; Faísca, P.F.N.; Rodrigues, M.S. Morphological and Biophysical Study of S100A9 Protein Fibrils by Atomic Force Microscopy Imaging and Nanomechanical Analysis. Biomolecules 2024, 14, 1091. [Google Scholar] [CrossRef]
- Wang, C.; Klechikov, A.G.; Gharibyan, A.L.; Wärmländer, S.K.; Jarvet, J.; Zhao, L.; Jia, X.; Narayana, V.K.; Shankar, S.K.; Olofsson, A.; et al. The role of pro-inflammatory S100A9 in Alzheimer’s disease amyloid-neuroinflammatory cascade. Acta Neuropathol. 2014, 127, 507–522. [Google Scholar] [CrossRef]
- Dubois, B.; Hampel, H.; Feldman, H.H.; Scheltens, P.; Aisen, P.; Andrieu, S.; Bakardjian, H.; Benali, H.; Bertram, L.; Blennow, K.; et al. Preclinical Alzheimer’s disease: Definition, natural history, and diagnostic criteria. Alzheimer’s Dement. 2016, 12, 292–323. [Google Scholar] [CrossRef]
- Dubois, B.; Villain, N.; Frisoni, G.B.; Rabinovici, G.D.; Sabbagh, M.; Cappa, S.; Bejanin, A.; Bombois, S.; Epelbaum, S.; Teichmann, M.; et al. Clinical diagnosis of Alzheimer’s disease: Recommendations of the International Working Group. Lancet Neurol. 2021, 20, 484–496. [Google Scholar] [CrossRef]
- Valenzuela, M.J.; Sachdev, P. Magnetic resonance spectroscopy in AD. Neurology 2001, 56, 592–598. [Google Scholar] [CrossRef]
- Firbank, M.J.; Harrison, R.M.; O’Brien, J.T. A Comprehensive Review of Proton Magnetic Resonance Spectroscopy Studies in Dementia and Parkinson’s Disease. Dement. Geriatr. Cogn. Disord. 2002, 14, 64–76. [Google Scholar] [CrossRef] [PubMed]
- McKiernan, E.; Su, L.; O’Brien, J. MRS in neurodegenerative dementias, prodromal syndromes and at-risk states: A systematic review of the literature. NMR Biomed. 2023, 36, e4896. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.; Barker, P.B. Various MRS Application Tools for Alzheimer Disease and Mild Cognitive Impairment. Am. J. Neuroradiol. 2014, 35, S4–S11. [Google Scholar] [CrossRef] [PubMed]
- Piersson, A.D.; Mohamad, M.; Rajab, F.; Suppiah, S. Cerebrospinal Fluid Amyloid Beta, Tau Levels, Apolipoprotein, and 1H-MRS Brain Metabolites in Alzheimer’s Disease: A Systematic Review. Acad. Radiol. 2021, 28, 1447–1463. [Google Scholar] [CrossRef]
- Wang, H.; Tan, L.; Wang, H.F.; Liu, Y.; Yin, R.H.; Wang, W.Y.; Chang, X.L.; Jiang, T.; Yu, J.T. Magnetic Resonance Spectroscopy in Alzheimer’s Disease: Systematic Review and Meta-Analysis. J. Alzheimer’s Dis. 2015, 46, 1049–1070. [Google Scholar] [CrossRef] [PubMed]
- Sheikh-Bahaei, N.; Chen, M.; Pappas, I. Magnetic Resonance Spectroscopy (MRS) in Alzheimer’s Disease. Methods Mol. Biol. 2024, 2785, 115–142. [Google Scholar] [CrossRef]
- Zhang, N.; Song, X.; Bartha, R.; Beyea, S.; D’Arcy, R.; Zhang, Y.; Rockwood, K. Advances in high-field magnetic resonance spectroscopy in Alzheimer’s disease. Curr. Alzheimer Res. 2014, 11, 367–388. [Google Scholar] [CrossRef]
- Graff-Radford, J.; Kantarci, K. Magnetic resonance spectroscopy in Alzheimer’s disease. Neuropsychiatr. Dis. Treat. 2013, 9, 687–696. [Google Scholar] [CrossRef]
- Murray, M.E.; Przybelski, S.A.; Lesnick, T.G.; Liesinger, A.M.; Spychalla, A.; Zhang, B.; Gunter, J.L.; Parisi, J.E.; Boeve, B.F.; Knopman, D.S.; et al. Early Alzheimer’s disease neuropathology detected by proton MR spectroscopy. J. Neurosci. 2014, 34, 16247–16255. [Google Scholar] [CrossRef]
- Öz, G.; Alger, J.R.; Barker, P.B.; Bartha, R.; Bizzi, A.; Boesch, C.; Bolan, P.J.; Brindle, K.M.; Cudalbu, C.; Dinçer, A.; et al. Clinical Proton MR Spectroscopy in Central Nervous System Disorders. Radiology 2014, 270, 658–679. [Google Scholar] [CrossRef]
- Moffett, J.R.; Ross, B.; Arun, P.; Madhavarao, C.N.; Namboodiri, A.M. N-Acetylaspartate in the CNS: From neurodiagnostics to neurobiology. Progress. Neurobiol. 2007, 81, 89–131. [Google Scholar] [CrossRef] [PubMed]
- Luyten, P.R.; den Hollander, J.A. Observation of metabolites in the human brain by MR spectroscopy. Radiology 1986, 161, 795–798. [Google Scholar] [CrossRef] [PubMed]
- Urenjak, J.; Williams, S.R.; Gadian, D.G.; Noble, M. Specific expression of N-acetylaspartate in neurons, oligodendrocyte-type-2 astrocyte progenitors, and immature oligodendrocytes in vitro. J. Neurochem. 1992, 59, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Tallan, H.H. Studies on the distribution of N-acetyl-L-aspartic acid in brain. J. Biol. Chem. 1957, 224, 41–45. [Google Scholar] [CrossRef] [PubMed]
- de Graaf, R.A. Vivo NMR Spectroscopy: Principles and Techniques, 2nd ed.; Wiley: Hoboken, NJ, USA, 2007; p. 592. [Google Scholar]
- Miller, B.L.; Moats, R.A.; Shonk, T.; Ernst, T.; Woolley, S.; Ross, B.D. Alzheimer disease: Depiction of increased cerebral myo-inositol with proton MR spectroscopy. Radiology 1993, 187, 433–437. [Google Scholar] [CrossRef]
- Huang, W.; Alexander, G.E.; Chang, L.; Shetty, H.U.; Krasuski, J.S.; Rapoport, S.I.; Schapiro, M.B. Brain metabolite concentration and dementia severity in Alzheimer’s disease: A (1)H MRS study. Neurology 2001, 57, 626–632. [Google Scholar] [CrossRef]
- Moats, R.A.; Ernst, T.; Shonk, T.K.; Ross, B.D. Abnormal cerebral metabolite concentrations in patients with probable Alzheimer disease. Magn. Reson. Med. 1994, 32, 110–115. [Google Scholar] [CrossRef]
- Shonk, T.K.; Moats, R.A.; Gifford, P.; Michaelis, T.; Mandigo, J.C.; Izumi, J.; Ross, B.D. Probable Alzheimer disease: Diagnosis with proton MR spectroscopy. Radiology 1995, 195, 65–72. [Google Scholar] [CrossRef]
- Frederick, B.B.; Satlin, A.; Yurgelun-Todd, D.A.; Renshaw, P.F. In vivo proton magnetic resonance spectroscopy of Alzheimer’s disease in the parietal and temporal lobes. Biol. Psychiatry 1997, 42, 147–150. [Google Scholar] [CrossRef]
- Parnetti, L.; Tarducci, R.; Presciutti, O.; Lowenthal, D.T.; Pippi, M.; Palumbo, B.; Gobbi, G.; Pelliccioli, G.P.; Senin, U. Proton magnetic resonance spectroscopy can differentiate Alzheimer’s disease from normal aging. Mech. Ageing Dev. 1997, 97, 9–14. [Google Scholar] [CrossRef]
- Jessen, F.; Block, W.; Träber, F.; Keller, E.; Flacke, S.; Papassotiropoulos, A.; Lamerichs, R.; Heun, R.; Schild, H.H. Proton MR spectroscopy detects a relative decrease of N-acetylaspartate in the medial temporal lobe of patients with AD. Neurology 2000, 55, 684–688. [Google Scholar] [CrossRef] [PubMed]
- Christiansen, P.; Schlosser, A.; Henriksen, O. Reduced N-acetylaspartate content in the frontal part of the brain in patients with probable Alzheimer’s disease. Magn. Reson. Imaging 1995, 13, 457–462. [Google Scholar] [CrossRef] [PubMed]
- Antuono, P.G.; Jones, J.L.; Wang, Y.; Li, S.J. Decreased glutamate + glutamine in Alzheimer’s disease detected in vivo with (1)H-MRS at 0.5 T. Neurology 2001, 56, 737–742. [Google Scholar] [CrossRef] [PubMed]
- Dixon, R.M.; Bradley, K.M.; Budge, M.M.; Styles, P.; Smith, A.D. Longitudinal quantitative proton magnetic resonance spectroscopy of the hippocampus in Alzheimer’s disease. Brain 2002, 125, 2332–2341. [Google Scholar] [CrossRef] [PubMed]
- Catani, M.; Cherubini, A.; Howard, R.; Tarducci, R.; Pelliccioli, G.P.; Piccirilli, M.; Gobbi, G.; Senin, U.; Mecocci, P. (1)H-MR spectroscopy differentiates mild cognitive impairment from normal brain aging. Neuroreport 2001, 12, 2315–2317. [Google Scholar] [CrossRef] [PubMed]
- Kantarci, K.; Petersen, R.C.; Boeve, B.F.; Knopman, D.S.; Tang-Wai, D.F.; O’Brien, P.C.; Weigand, S.D.; Edland, S.D.; Smith, G.E.; Ivnik, R.J.; et al. 1H MR spectroscopy in common dementias. Neurology 2004, 63, 1393–1398. [Google Scholar] [CrossRef]
- Shiino, A.; Watanabe, T.; Shirakashi, Y.; Kotani, E.; Yoshimura, M.; Morikawa, S.; Inubushi, T.; Akiguchi, I. The profile of hippocampal metabolites differs between Alzheimer’s disease and subcortical ischemic vascular dementia, as measured by proton magnetic resonance spectroscopy. J. Cereb. Blood Flow. Metab. 2012, 32, 805–815. [Google Scholar] [CrossRef]
- Hattori, N.; Abe, K.; Sakoda, S.; Sawada, T. Proton MR spectroscopic study at 3 Tesla on glutamate/glutamine in Alzheimer’s disease. Neuroreport 2002, 13, 183–186. [Google Scholar] [CrossRef]
- Kantarci, K.; Jack, C.R.; Xu, Y.C.; Campeau, N.G.; O’Brien, P.C.; Smith, G.E.; Ivnik, R.J.; Boeve, B.F.; Kokmen, E.; Tangalos, E.G.; et al. Regional metabolic patterns in mild cognitive impairment and Alzheimer’s disease. A 1h MRS Study 2000, 55, 210–217. [Google Scholar] [CrossRef]
- Michaelis, T.; Merboldt, K.D.; Hänicke, W.; Gyngell, M.L.; Bruhn, H.; Frahm, J. On the identification of cerebral metabolites in localized 1H NMR spectra of human brain in vivo. NMR Biomed. 1991, 4, 90–98. [Google Scholar] [CrossRef]
- Govindaraju, V.; Young, K.; Maudsley, A.A. Proton NMR chemical shifts and coupling constants for brain metabolites. NMR Biomed. 2000, 13, 129–153. [Google Scholar] [CrossRef] [PubMed]
- Rae, C.D. A guide to the metabolic pathways and function of metabolites observed in human brain 1H magnetic resonance spectra. Neurochem. Res. 2014, 39, 1–36. [Google Scholar] [CrossRef] [PubMed]
- Ratai, E.-M.; Alshikho, M.J.; Zürcher, N.R.; Loggia, M.L.; Cebulla, C.L.; Cernasov, P.; Reynolds, B.; Fish, J.; Seth, R.; Babu, S.; et al. Integrated imaging of [11C]-PBR28 PET, MR diffusion and magnetic resonance spectroscopy 1H-MRS in amyotrophic lateral sclerosis. NeuroImage Clin. 2018, 20, 357–364. [Google Scholar] [CrossRef] [PubMed]
- Nutma, E.; Fancy, N.; Weinert, M.; Tsartsalis, S.; Marzin, M.C.; Muirhead, R.C.J.; Falk, I.; Breur, M.; de Bruin, J.; Hollaus, D.; et al. Translocator protein is a marker of activated microglia in rodent models but not human neurodegenerative diseases. Nat. Commun. 2023, 14, 5247. [Google Scholar] [CrossRef] [PubMed]
- Best, J.G.; Stagg, C.J.; Dennis, A. Chapter 2.5—Other Significant Metabolites: Myo-Inositol, GABA, Glutamine, and Lactate. In Magnetic Resonance Spectroscopy; Stagg, C., Rothman, D., Eds.; Academic Press: San Diego, CA, USA, 2014; pp. 122–138. [Google Scholar]
- Bitsch, A.; Bruhn, H.; Vougioukas, V.; Stringaris, A.; Lassmann, H.; Frahm, J.; Brück, W. Inflammatory CNS demyelination: Histopathologic correlation with in vivo quantitative proton MR spectroscopy. AJNR Am. J. Neuroradiol. 1999, 20, 1619–1627. [Google Scholar]
- Schreiner, S.J.; Van Bergen, J.M.G.; Gietl, A.F.; Buck, A.; Hock, C.; Pruessmann, K.P.; Henning, A.; Unschuld, P.G. Gray matter gamma-hydroxy-butyric acid and glutamate reflect beta-amyloid burden at old age. Alzheimer’s Dement. 2024, 16, e12587. [Google Scholar] [CrossRef]
- Matsuoka, K.; Hirata, K.; Kokubo, N.; Maeda, T.; Tagai, K.; Endo, H.; Takahata, K.; Shinotoh, H.; Ono, M.; Seki, C.; et al. Investigating neural dysfunction with abnormal protein deposition in Alzheimer’s disease through magnetic resonance spectroscopic imaging, plasma biomarkers, and positron emission tomography. Neuroimage Clin. 2024, 41, 103560. [Google Scholar] [CrossRef]
- Spotorno, N.; Najac, C.; Stomrud, E.; Mattsson-Carlgren, N.; Palmqvist, S.; van Westen, D.; Ronen, I.; Hansson, O. Astrocytic function is associated with both amyloid-beta and tau pathology in non-demented APOE ϵ4 carriers. Brain Commun. 2022, 4, fcac135. [Google Scholar] [CrossRef]
- Hone-Blanchet, A.; Bohsali, A.; Krishnamurthy, L.C.; Shahid, S.S.; Lin, Q.; Zhao, L.; Bisht, A.S.; John, S.E.; Loring, D.; Goldstein, F.; et al. Frontal Metabolites and Alzheimer’s Disease Biomarkers in Healthy Older Women and Women Diagnosed with Mild Cognitive Impairment. J. Alzheimer’s Dis. 2022, 87, 1131–1141. [Google Scholar] [CrossRef]
- Chen, Q.; Abrigo, J.; Liu, W.; Han, E.Y.; Yeung, D.K.W.; Shi, L.; Au, L.W.C.; Deng, M.; Chen, S.; Leung, E.Y.L.; et al. Lower Posterior Cingulate N-Acetylaspartate to Creatine Level in Early Detection of Biologically Defined Alzheimer’s Disease. Brain Sci. 2022, 12, 722. [Google Scholar] [CrossRef]
- Kara, F.; Joers, J.M.; Deelchand, D.K.; Park, Y.W.; Przybelski, S.A.; Lesnick, T.G.; Senjem, M.L.; Zeydan, B.; Knopman, D.S.; Lowe, V.J.; et al. (1)H MR spectroscopy biomarkers of neuronal and synaptic function are associated with tau deposition in cognitively unimpaired older adults. Neurobiol. Aging 2022, 112, 16–26. [Google Scholar] [CrossRef] [PubMed]
- Voevodskaya, O.; Poulakis, K.; Sundgren, P.; van Westen, D.; Palmqvist, S.; Wahlund, L.-O.; Stomrud, E.; Hansson, O.; Westman, E.; Swedish Bio, F.S.G. Brain myoinositol as a potential marker of amyloid-related pathology: A longitudinal study. Neurology 2019, 92, e395–e405. [Google Scholar] [CrossRef] [PubMed]
- Mullins, R.; Reiter, D.; Kapogiannis, D. Magnetic resonance spectroscopy reveals abnormalities of glucose metabolism in the Alzheimer’s brain. Ann. Clin. Transl. Neurol. 2018, 5, 262–272. [Google Scholar] [CrossRef] [PubMed]
- Suri, S.; Emir, U.; Stagg, C.J.; Near, J.; Mekle, R.; Schubert, F.; Zsoldos, E.; Mahmood, A.; Singh-Manoux, A.; Kivimäki, M.; et al. Effect of age and the APOE gene on metabolite concentrations in the posterior cingulate cortex. Neuroimage 2017, 152, 509–516. [Google Scholar] [CrossRef] [PubMed]
- Waragai, M.; Moriya, M.; Nojo, T. Decreased N-Acetyl Aspartate/Myo-Inositol Ratio in the Posterior Cingulate Cortex Shown by Magnetic Resonance Spectroscopy May Be One of the Risk Markers of Preclinical Alzheimer’s Disease: A 7-Year Follow-Up Study. J. Alzheimer’s Dis. 2017, 60, 1411–1427. [Google Scholar] [CrossRef]
- Chiang, G.C.; Mao, X.; Kang, G.; Chang, E.; Pandya, S.; Vallabhajosula, S.; Isaacson, R.; Ravdin, L.D.; Shungu, D.C. Relationships among Cortical Glutathione Levels, Brain Amyloidosis, and Memory in Healthy Older Adults Investigated In Vivo with (1)H-MRS and Pittsburgh Compound-B PET. AJNR Am. J. Neuroradiol. 2017, 38, 1130–1137. [Google Scholar] [CrossRef]
- Zeydan, B.; Deelchand, D.K.; Tosakulwong, N.; Lesnick, T.G.; Kantarci, O.H.; Machulda, M.M.; Knopman, D.S.; Lowe, V.J.; Jack, C.R., Jr.; Petersen, R.C.; et al. Decreased Glutamate Levels in Patients with Amnestic Mild Cognitive Impairment: An sLASER Proton MR Spectroscopy and PiB-PET Study. J. Neuroimaging 2017, 27, 630–636. [Google Scholar] [CrossRef]
- Nedelska, Z.; Przybelski, S.A.; Lesnick, T.G.; Schwarz, C.G.; Lowe, V.J.; Machulda, M.M.; Kremers, W.K.; Mielke, M.M.; Roberts, R.O.; Boeve, B.F.; et al. H-1-MRS metabolites and rate of beta-amyloid accumulation on serial PET in clinically normal adults. Neurology 2017, 89, 1391–1399. [Google Scholar] [CrossRef]
- Voevodskaya, O.; Sundgren, P.C.; Strandberg, O.; Zetterberg, H.; Minthon, L.; Blennow, K.; Wahlund, L.O.; Westman, E.; Hansson, O.; Swedish Bio, F.s.g. Myo-inositol changes precede amyloid pathology and relate to APOE genotype in Alzheimer disease. Neurology 2016, 86, 1754–1761. [Google Scholar] [CrossRef]
- Yin, Z.; Wu, W.; Liu, R.; Liang, X.; Yu, T.; Chen, X.; Feng, J.; Guo, A.; Xie, Y.; Yang, H.; et al. APOE genotype and age modifies the correlation between cognitive status and metabolites from hippocampus by a 2D (1)H-MRS in non-demented elders. PeerJ 2015, 3, e1202. [Google Scholar] [CrossRef]
- Riese, F.; Gietl, A.; Zölch, N.; Henning, A.; O’Gorman, R.; Kälin, A.M.; Leh, S.E.; Buck, A.; Warnock, G.; Edden, R.A.E.; et al. Posterior cingulate γ-aminobutyric acid and glutamate/glutamine are reduced in amnestic mild cognitive impairment and are unrelated to amyloid deposition and apolipoprotein E genotype. Neurobiol. Aging 2015, 36, 53–59. [Google Scholar] [CrossRef]
- Gomar, J.J.; Gordon, M.L.; Dickinson, D.; Kingsley, P.B.; Uluğ, A.M.; Keehlisen, L.; Huet, S.; Buthorn, J.J.; Koppel, J.; Christen, E.; et al. APOE genotype modulates proton magnetic resonance spectroscopy metabolites in the aging brain. Biol. Psychiatry 2014, 75, 686–692. [Google Scholar] [CrossRef]
- Bittner, D.M.; Heinze, H.J.; Kaufmann, J. Association of 1H-MR spectroscopy and cerebrospinal fluid biomarkers in Alzheimer’s disease: Diverging behavior at three different brain regions. J. Alzheimer’s Dis. 2013, 36, 155–163. [Google Scholar] [CrossRef]
- Jessen, F.; Lewczuk, P.; Gür, O.; Block, W.; Ende, G.; Frölich, L.; Hammen, T.; Arlt, S.; Kornhuber, J.; Kucinski, T.; et al. Association of N-acetylaspartate and cerebrospinal fluid Aβ42 in dementia. J. Alzheimer’s Dis. 2011, 27, 393–399. [Google Scholar] [CrossRef]
- Kantarci, K.; Lowe, V.; Przybelski, S.A.; Senjem, M.L.; Weigand, S.D.; Ivnik, R.J.; Roberts, R.; Geda, Y.E.; Boeve, B.F.; Knopman, D.S.; et al. Magnetic resonance spectroscopy, β-amyloid load, and cognition in a population-based sample of cognitively normal older adults. Neurology 2011, 77, 951–958. [Google Scholar] [CrossRef]
- Kantarci, K.; Knopman, D.S.; Dickson, D.W.; Parisi, J.E.; Whitwell, J.L.; Weigand, S.D.; Josephs, K.A.; Boeve, B.F.; Petersen, R.C.; Jack, C.R., Jr. Alzheimer disease: Postmortem neuropathologic correlates of antemortem 1H MR spectroscopy metabolite measurements. Radiology 2008, 248, 210–220. [Google Scholar] [CrossRef]
- Kantarci, K.; Smith, G.E.; Ivnik, R.J.; Petersen, R.C.; Boeve, B.F.; Knopman, D.S.; Tangalos, E.G.; Jack, C.R., Jr. 1H magnetic resonance spectroscopy, cognitive function, and apolipoprotein E genotype in normal aging, mild cognitive impairment and Alzheimer’s disease. J. Int. Neuropsychol. Soc. 2002, 8, 934–942. [Google Scholar] [CrossRef]
- Klunk, W.E.; Panchalingam, K.; McClure, R.J.; Stanley, J.A.; Pettegrew, J.W. Metabolic alterations in postmortem Alzheimer’s disease brain are exaggerated by Apo-E4. Neurobiol. Aging 1998, 19, 511–515. [Google Scholar] [CrossRef]
- Lin, J.C.; Gant, N. Chapter 2.3—The Biochemistry of Choline. In Magnetic Resonance Spectroscopy; Stagg, C., Rothman, D., Eds.; Academic Press: San Diego, CA, USA, 2014; pp. 104–110. [Google Scholar]
- Miller, B.L.; Chang, L.; Booth, R.; Ernst, T.; Cornford, M.; Nikas, D.; McBride, D.; Jenden, D.J. In vivo 1H MRS choline: Correlation with in vitro chemistry/histology. Life Sci. 1996, 58, 1929–1935. [Google Scholar] [CrossRef]
- Barker, P.B.; Breiter, S.N.; Soher, B.J.; Chatham, J.C.; Forder, J.R.; Samphilipo, M.A.; Magee, C.A.; Anderson, J.H. Quantitative proton spectroscopy of canine brain: In vivo and in vitro correlations. Magn. Reson. Med. 1994, 32, 157–163. [Google Scholar] [CrossRef]
- Meyerhoff, D.J.; MacKay, S.; Constans, J.M.; Norman, D.; Van Dyke, C.; Fein, G.; Weiner, M.W. Axonal injury and membrane alterations in Alzheimer’s disease suggested by in vivo proton magnetic resonance spectroscopic imaging. Ann. Neurol. 1994, 36, 40–47. [Google Scholar] [CrossRef]
- MacKay, S.; Meyerhoff, D.J.; Constans, J.M.; Norman, D.; Fein, G.; Weiner, M.W. Regional gray and white matter metabolite differences in subjects with AD, with subcortical ischemic vascular dementia, and elderly controls with 1H magnetic resonance spectroscopic imaging. Arch. Neurol. 1996, 53, 167–174. [Google Scholar] [CrossRef]
- MacKay, S.; Ezekiel, F.; Di Sclafani, V.; Meyerhoff, D.J.; Gerson, J.; Norman, D.; Fein, G.; Weiner, M.W. Alzheimer disease and subcortical ischemic vascular dementia: Evaluation by combining MR imaging segmentation and H-1 MR spectroscopic imaging. Radiology 1996, 198, 537–545. [Google Scholar] [CrossRef]
- Fayed, N.; Andrés, E.; Viguera, L.; Modrego, P.J.; Garcia-Campayo, J. Higher glutamate+glutamine and reduction of N-acetylaspartate in posterior cingulate according to age range in patients with cognitive impairment and/or pain. Acad. Radiol. 2014, 21, 1211–1217. [Google Scholar] [CrossRef]
- Su, L.; Blamire, A.M.; Watson, R.; He, J.; Hayes, L.; O’Brien, J.T. Whole-brain patterns of 1H-magnetic resonance spectroscopy imaging in Alzheimer’s disease and dementia with Lewy bodies. Transl. Psychiatry 2016, 6, e877. [Google Scholar] [CrossRef]
- Chantal, S.; Labelle, M.; Bouchard, R.W.; Braun, C.M.; Boulanger, Y. Correlation of regional proton magnetic resonance spectroscopic metabolic changes with cognitive deficits in mild Alzheimer disease. Arch. Neurol. 2002, 59, 955–962. [Google Scholar] [CrossRef]
- Wurtman, R.J.; Blusztajn, J.K.; Maire, J.C. “Autocannibalism” of choline-containing membrane phospholipids in the pathogenesis of Alzheimer’s disease-A hypothesis. Neurochem. Int. 1985, 7, 369–372. [Google Scholar] [CrossRef]
- Satlin, A.; Bodick, N.; Offen, W.W.; Renshaw, P.F. Brain proton magnetic resonance spectroscopy (1H-MRS) in Alzheimer’s disease: Changes after treatment with xanomeline, an M1 selective cholinergic agonist. Am. J. Psychiatry 1997, 154, 1459–1461. [Google Scholar] [CrossRef]
- Hertz, L. Functional interactions between neurons and astrocytes I. Turnover and metabolism of putative amino acid transmitters. Progress. Neurobiol. 1979, 13, 277–323. [Google Scholar] [CrossRef]
- Bak, L.K.; Schousboe, A.; Waagepetersen, H.S. The glutamate/GABA-glutamine cycle: Aspects of transport, neurotransmitter homeostasis and ammonia transfer. J. Neurochem. 2006, 98, 641–653. [Google Scholar] [CrossRef]
- Hyman, B.T.; Van Hoesen, G.W.; Damasio, A.R. Alzheimer’s disease: Glutamate depletion in the hippocampal perforant pathway zone. Ann. Neurol. 1987, 22, 37–40. [Google Scholar] [CrossRef]
- Rupsingh, R.; Borrie, M.; Smith, M.; Wells, J.L.; Bartha, R. Reduced hippocampal glutamate in Alzheimer disease. Neurobiol. Aging 2011, 32, 802–810. [Google Scholar] [CrossRef]
- Puts, N.A.; Edden, R.A. In vivo magnetic resonance spectroscopy of GABA: A methodological review. Prog. Nucl. Magn. Reson. Spectrosc. 2012, 60, 29–41. [Google Scholar] [CrossRef]
- Huang, D.; Liu, D.; Yin, J.; Qian, T.; Shrestha, S.; Ni, H. Glutamate-glutamine and GABA in brain of normal aged and patients with cognitive impairment. Eur. Radiol. 2017, 27, 2698–2705. [Google Scholar] [CrossRef]
- Rae, C.D.; Williams, S.R. Glutathione in the human brain: Review of its roles and measurement by magnetic resonance spectroscopy. Anal. Biochem. 2017, 529, 127–143. [Google Scholar] [CrossRef]
- Mandal, P.K.; Tripathi, M.; Sugunan, S. Brain oxidative stress: Detection and mapping of anti-oxidant marker ‘Glutathione’ in different brain regions of healthy male/female, MCI and Alzheimer patients using non-invasive magnetic resonance spectroscopy. Biochem. Biophys. Res. Commun. 2012, 417, 43–48. [Google Scholar] [CrossRef]
- Shukla, D.; Mandal, P.K.; Tripathi, M.; Vishwakarma, G.; Mishra, R.; Sandal, K. Quantitation of in vivo brain glutathione conformers in cingulate cortex among age-matched control, MCI, and AD patients using MEGA-PRESS. Hum. Brain Mapp. 2020, 41, 194–217. [Google Scholar] [CrossRef]
- Mandal, P.K.; Saharan, S.; Tripathi, M.; Murari, G. Brain Glutathione Levels—A Novel Biomarker for Mild Cognitive Impairment and Alzheimer’s Disease. Biol. Psychiatry 2015, 78, 702–710. [Google Scholar] [CrossRef]
- Zabel, M.; Nackenoff, A.; Kirsch, W.M.; Harrison, F.E.; Perry, G.; Schrag, M. Markers of oxidative damage to lipids, nucleic acids and proteins and antioxidant enzymes activities in Alzheimer’s disease brain: A meta-analysis in human pathological specimens. Free Radic. Biol. Med. 2018, 115, 351–360. [Google Scholar] [CrossRef]
- Ernst, T.; Chang, L.; Melchor, R.; Mehringer, C.M. Frontotemporal dementia and early Alzheimer disease: Differentiation with frontal lobe H-1 MR spectroscopy. Radiology 1997, 203, 829–836. [Google Scholar] [CrossRef]
- Stoppe, G.; Bruhn, H.; Pouwels, P.J.; Hänicke, W.; Frahm, J. Alzheimer disease: Absolute quantification of cerebral metabolites in vivo using localized proton magnetic resonance spectroscopy. Alzheimer Dis. Assoc. Disord. 2000, 14, 112–119. [Google Scholar] [CrossRef]
- Harris, R.A.; Tindale, L.; Lone, A.; Singh, O.; Macauley, S.L.; Stanley, M.; Holtzman, D.M.; Bartha, R.; Cumming, R.C. Aerobic Glycolysis in the Frontal Cortex Correlates with Memory Performance in Wild-Type Mice But Not the APP/PS1 Mouse Model of Cerebral Amyloidosis. J. Neurosci. 2016, 36, 1871–1878. [Google Scholar] [CrossRef]
- Le Page, L.M.; Guglielmetti, C.; Taglang, C.; Chaumeil, M.M. Imaging Brain Metabolism Using Hyperpolarized (13)C Magnetic Resonance Spectroscopy. Trends Neurosci. 2020, 43, 343–354. [Google Scholar] [CrossRef]
- Lee, C.Y.; Soliman, H.; Geraghty, B.J.; Chen, A.P.; Connelly, K.A.; Endre, R.; Perks, W.J.; Heyn, C.; Black, S.E.; Cunningham, C.H. Lactate topography of the human brain using hyperpolarized (13)C-MRI. Neuroimage 2020, 204, 116202. [Google Scholar] [CrossRef]
- Wyss, M.; Kaddurah-Daouk, R. Creatine and creatinine metabolism. Physiol. Rev. 2000, 80, 1107–1213. [Google Scholar] [CrossRef]
- Rackayova, V.; Cudalbu, C.; Pouwels, P.J.W.; Braissant, O. Creatine in the central nervous system: From magnetic resonance spectroscopy to creatine deficiencies. Anal. Biochem. 2017, 529, 144–157. [Google Scholar] [CrossRef]
- Chang, L.; Ernst, T.; Poland, R.E.; Jenden, D.J. In vivo proton magnetic resonance spectroscopy of the normal aging human brain. Life Sci. 1996, 58, 2049–2056. [Google Scholar] [CrossRef]
- Maudsley, A.A.; Domenig, C.; Govind, V.; Darkazanli, A.; Studholme, C.; Arheart, K.; Bloomer, C. Mapping of brain metabolite distributions by volumetric proton MR spectroscopic imaging (MRSI). Magn. Reson. Med. 2009, 61, 548–559. [Google Scholar] [CrossRef]
- Saunders, D.E.; Howe, F.A.; van den Boogaart, A.; Griffiths, J.R.; Brown, M.M. Aging of the adult human brain: In vivo quantitation of metabolite content with proton magnetic resonance spectroscopy. J. Magn. Reson. Imaging 1999, 9, 711–716. [Google Scholar] [CrossRef]
- Oppenheimer, S.M.; Bryan, R.N.; Conturo, T.E.; Soher, B.J.; Preziosi, T.J.; Barker, P.B. Proton magnetic resonance spectroscopy and gadolinium-DTPA perfusion imaging of asymptomatic MRI white matter lesions. Magn. Reson. Med. 1995, 33, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Ongür, D.; Prescot, A.P.; Jensen, J.E.; Cohen, B.M.; Renshaw, P.F. Creatine abnormalities in schizophrenia and bipolar disorder. Psychiatry Res. 2009, 172, 44–48. [Google Scholar] [CrossRef] [PubMed]
- Bai, X.; Edden, R.A.; Gao, F.; Wang, G.; Wu, L.; Zhao, B.; Wang, M.; Chan, Q.; Chen, W.; Barker, P.B. Decreased γ-aminobutyric acid levels in the parietal region of patients with Alzheimer’s disease. J. Magn. Reson. Imaging 2015, 41, 1326–1331. [Google Scholar] [CrossRef] [PubMed]
- Mielke, M.M.; Dage, J.L.; Frank, R.D.; Algeciras-Schimnich, A.; Knopman, D.S.; Lowe, V.J.; Bu, G.; Vemuri, P.; Graff-Radford, J.; Jack, C.R., Jr.; et al. Performance of plasma phosphorylated tau 181 and 217 in the community. Nat. Med. 2022, 28, 1398–1405. [Google Scholar] [CrossRef] [PubMed]
- Jack, C.R., Jr.; Wiste, H.J.; Algeciras-Schimnich, A.; Weigand, S.D.; Figdore, D.J.; Lowe, V.J.; Vemuri, P.; Graff-Radford, J.; Ramanan, V.K.; Knopman, D.S.; et al. Comparison of plasma biomarkers and amyloid PET for predicting memory decline in cognitively unimpaired individuals. Alzheimer’s Dement. 2024, 20, 2143–2154. [Google Scholar] [CrossRef]
- Pais, M.V.; Forlenza, O.V.; Diniz, B.S. Plasma Biomarkers of Alzheimer’s Disease: A Review of Available Assays, Recent Developments, and Implications for Clinical Practice. J. Alzheimers Dis. Rep. 2023, 7, 355–380. [Google Scholar] [CrossRef]
- Janelidze, S.; Teunissen, C.E.; Zetterberg, H.; Allué, J.A.; Sarasa, L.; Eichenlaub, U.; Bittner, T.; Ovod, V.; Verberk, I.M.W.; Toba, K.; et al. Head-to-Head Comparison of 8 Plasma Amyloid-β 42/40 Assays in Alzheimer Disease. JAMA Neurol. 2021, 78, 1375–1382. [Google Scholar] [CrossRef]
- Nabers, A.; Perna, L.; Lange, J.; Mons, U.; Schartner, J.; Güldenhaupt, J.; Saum, K.U.; Janelidze, S.; Holleczek, B.; Rujescu, D.; et al. Amyloid blood biomarker detects Alzheimer’s disease. EMBO Mol. Med. 2018, 10, e8763. [Google Scholar] [CrossRef]
- Nakamura, A.; Kaneko, N.; Villemagne, V.L.; Kato, T.; Doecke, J.; Doré, V.; Fowler, C.; Li, Q.-X.; Martins, R.; Rowe, C.; et al. High performance plasma amyloid-β biomarkers for Alzheimer’s disease. Nature 2018, 554, 249–254. [Google Scholar] [CrossRef]
- Janelidze, S.; Mattsson, N.; Palmqvist, S.; Smith, R.; Beach, T.G.; Serrano, G.E.; Chai, X.; Proctor, N.K.; Eichenlaub, U.; Zetterberg, H.; et al. Plasma P-tau181 in Alzheimer’s disease: Relationship to other biomarkers, differential diagnosis, neuropathology and longitudinal progression to Alzheimer’s dementia. Nat. Med. 2020, 26, 379–386. [Google Scholar] [CrossRef]
- Karikari, T.K.; Pascoal, T.A.; Ashton, N.J.; Janelidze, S.; Benedet, A.L.; Rodriguez, J.L.; Chamoun, M.; Savard, M.; Kang, M.S.; Therriault, J.; et al. Blood phosphorylated tau 181 as a biomarker for Alzheimer’s disease: A diagnostic performance and prediction modelling study using data from four prospective cohorts. Lancet Neurol. 2020, 19, 422–433. [Google Scholar] [CrossRef]
- Janelidze, S.; Berron, D.; Smith, R.; Strandberg, O.; Proctor, N.K.; Dage, J.L.; Stomrud, E.; Palmqvist, S.; Mattsson-Carlgren, N.; Hansson, O. Associations of Plasma Phospho-Tau217 Levels With Tau Positron Emission Tomography in Early Alzheimer Disease. JAMA Neurol. 2021, 78, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Mielke, M.M.; Hagen, C.E.; Xu, J.; Chai, X.; Vemuri, P.; Lowe, V.J.; Airey, D.C.; Knopman, D.S.; Roberts, R.O.; Machulda, M.M.; et al. Plasma phospho-tau181 increases with Alzheimer’s disease clinical severity and is associated with tau- and amyloid-positron emission tomography. Alzheimer’s Dement. 2018, 14, 989–997. [Google Scholar] [CrossRef] [PubMed]
- Gafson, A.R.; Barthélemy, N.R.; Bomont, P.; Carare, R.O.; Durham, H.D.; Julien, J.P.; Kuhle, J.; Leppert, D.; Nixon, R.A.; Weller, R.O.; et al. Neurofilaments: Neurobiological foundations for biomarker applications. Brain 2020, 143, 1975–1998. [Google Scholar] [CrossRef] [PubMed]
- Mattsson, N.; Cullen, N.C.; Andreasson, U.; Zetterberg, H.; Blennow, K. Association Between Longitudinal Plasma Neurofilament Light and Neurodegeneration in Patients With Alzheimer Disease. JAMA Neurol. 2019, 76, 791–799. [Google Scholar] [CrossRef]
- Mielke, M.M.; Syrjanen, J.A.; Blennow, K.; Zetterberg, H.; Vemuri, P.; Skoog, I.; Machulda, M.M.; Kremers, W.K.; Knopman, D.S.; Jack, C., Jr.; et al. Plasma and CSF neurofilament light: Relation to longitudinal neuroimaging and cognitive measures. Neurology 2019, 93, e252–e260. [Google Scholar] [CrossRef]
- Verberk, I.M.W.; Laarhuis, M.B.; van den Bosch, K.A.; Ebenau, J.L.; van Leeuwenstijn, M.; Prins, N.D.; Scheltens, P.; Teunissen, C.E.; van der Flier, W.M. Serum markers glial fibrillary acidic protein and neurofilament light for prognosis and monitoring in cognitively normal older people: A prospective memory clinic-based cohort study. Lancet Healthy Longev. 2021, 2, e87–e95. [Google Scholar] [CrossRef]
- Heller, C.; Foiani, M.S.; Moore, K.; Convery, R.; Bocchetta, M.; Neason, M.; Cash, D.M.; Thomas, D.; Greaves, C.V.; Woollacott, I.O.; et al. Plasma glial fibrillary acidic protein is raised in progranulin-associated frontotemporal dementia. J. Neurol. Neurosurg. Psychiatry 2020, 91, 263–270. [Google Scholar] [CrossRef]
- Liu, H.; Zhang, D.; Lin, H.; Zhang, Q.; Zheng, L.; Zheng, Y.; Yin, X.; Li, Z.; Liang, S.; Huang, S. Meta-Analysis of Neurochemical Changes Estimated via Magnetic Resonance Spectroscopy in Mild Cognitive Impairment and Alzheimer’s Disease. Front. Aging Neurosci. 2021, 13, 738971. [Google Scholar] [CrossRef]
- Braak, H.; Braak, E. Neuropathological stageing of Alzheimer-related changes. Acta Neuropathol. 1991, 82, 239–259. [Google Scholar] [CrossRef]
- Thal, D.R.; Rüb, U.; Orantes, M.; Braak, H. Phases of A beta-deposition in the human brain and its relevance for the development of AD. Neurology 2002, 58, 1791–1800. [Google Scholar] [CrossRef]
- Park, Y.W.; Deelchand, D.K.; Joers, J.M.; Hanna, B.; Berrington, A.; Gillen, J.S.; Kantarci, K.; Soher, B.J.; Barker, P.B.; Park, H.; et al. AutoVOI: Real-time automatic prescription of volume-of-interest for single voxel spectroscopy. Magn. Reson. Med. 2018, 80, 1787–1798. [Google Scholar] [CrossRef] [PubMed]
- Kantarci, K.; Jicha, G.A. Development of 1H MRS biomarkers for tracking early predementia Alzheimer disease. Neurology 2019, 92, 209–210. [Google Scholar] [CrossRef] [PubMed]
- Deelchand, D.K.; Adanyeguh, I.M.; Emir, U.E.; Nguyen, T.-M.; Valabregue, R.; Henry, P.-G.; Mochel, F.; Öz, G. Two-site reproducibility of cerebellar and brainstem neurochemical profiles with short-echo, single-voxel MRS at 3T. Magn. Reson. Med. 2015, 73, 1718–1725. [Google Scholar] [CrossRef] [PubMed]
- Öz, G.; Deelchand, D.K.; Wijnen, J.P.; Mlynárik, V.; Xin, L.; Mekle, R.; Noeske, R.; Scheenen, T.W.J.; Tkáč, I. Advanced single voxel (1) H magnetic resonance spectroscopy techniques in humans: Experts’ consensus recommendations. NMR Biomed. 2020, e4236. [Google Scholar] [CrossRef] [PubMed]
- Bell, T.K.; Godfrey, K.J.; Ware, A.L.; Yeates, K.O.; Harris, A.D. Harmonization of multi-site MRS data with ComBat. NeuroImage 2022, 257, 119330. [Google Scholar] [CrossRef]
- Harris, A.D.; Amiri, H.; Bento, M.; Cohen, R.; Ching, C.R.K.; Cudalbu, C.; Dennis, E.L.; Doose, A.; Ehrlich, S.; Kirov, I.I.; et al. Harmonization of multi-scanner in vivo magnetic resonance spectroscopy: ENIGMA consortium task group considerations. Front. Neurol. 2023, 13, 1045678. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kara, F.; Kantarci, K. Understanding Proton Magnetic Resonance Spectroscopy Neurochemical Changes Using Alzheimer’s Disease Biofluid, PET, Postmortem Pathology Biomarkers, and APOE Genotype. Int. J. Mol. Sci. 2024, 25, 10064. https://doi.org/10.3390/ijms251810064
Kara F, Kantarci K. Understanding Proton Magnetic Resonance Spectroscopy Neurochemical Changes Using Alzheimer’s Disease Biofluid, PET, Postmortem Pathology Biomarkers, and APOE Genotype. International Journal of Molecular Sciences. 2024; 25(18):10064. https://doi.org/10.3390/ijms251810064
Chicago/Turabian StyleKara, Firat, and Kejal Kantarci. 2024. "Understanding Proton Magnetic Resonance Spectroscopy Neurochemical Changes Using Alzheimer’s Disease Biofluid, PET, Postmortem Pathology Biomarkers, and APOE Genotype" International Journal of Molecular Sciences 25, no. 18: 10064. https://doi.org/10.3390/ijms251810064