

Transcriptomic Insights into the Atrial Fibrillation Susceptibility Locus near the MYOZ1 and SYNPO2L Genes
 , , , , and
, , , , and
Abstract
:1. Introduction
2. Results
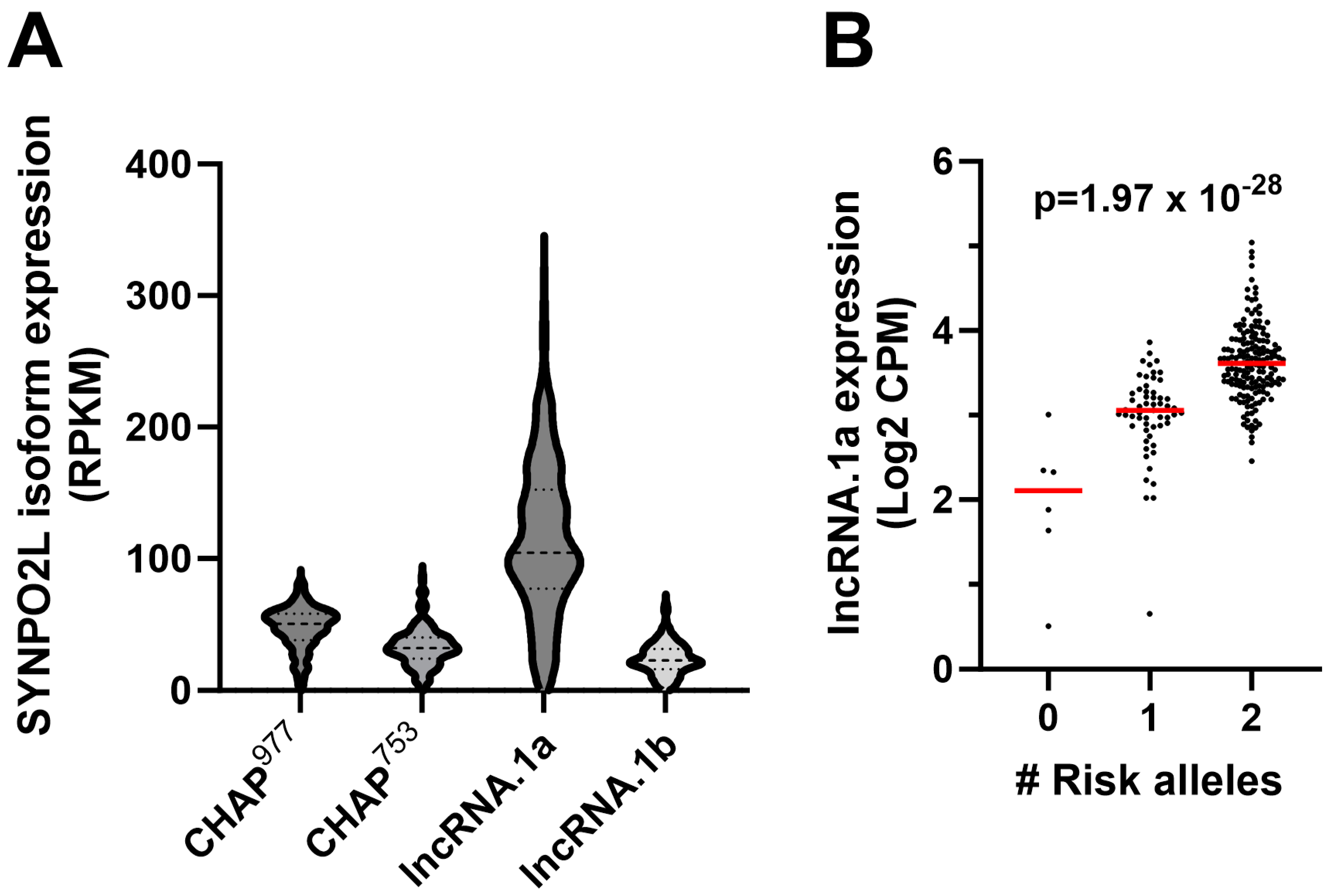
2.1. RNAseq Analysis and Identification of Transcript Isoform-Specific eQTLs
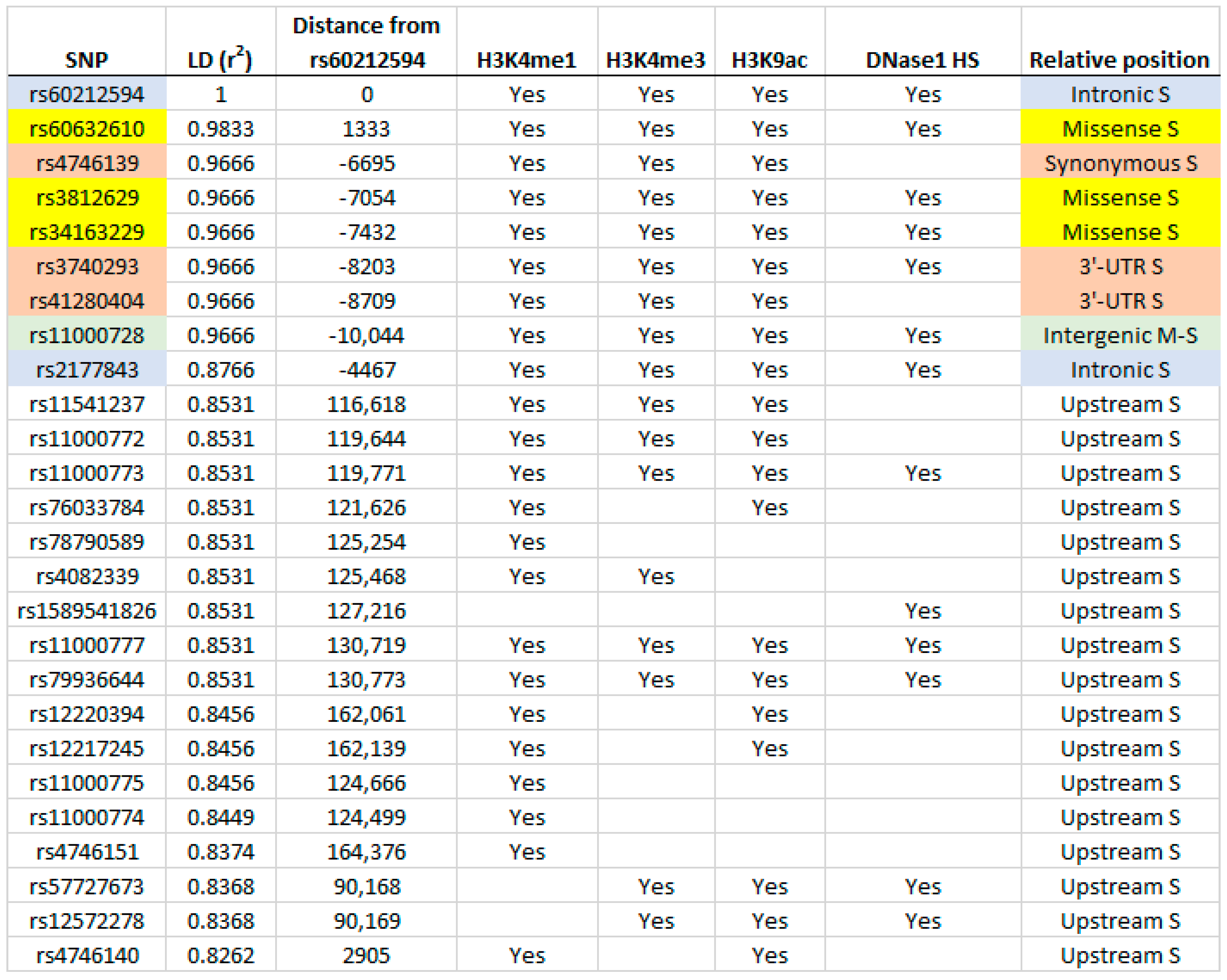
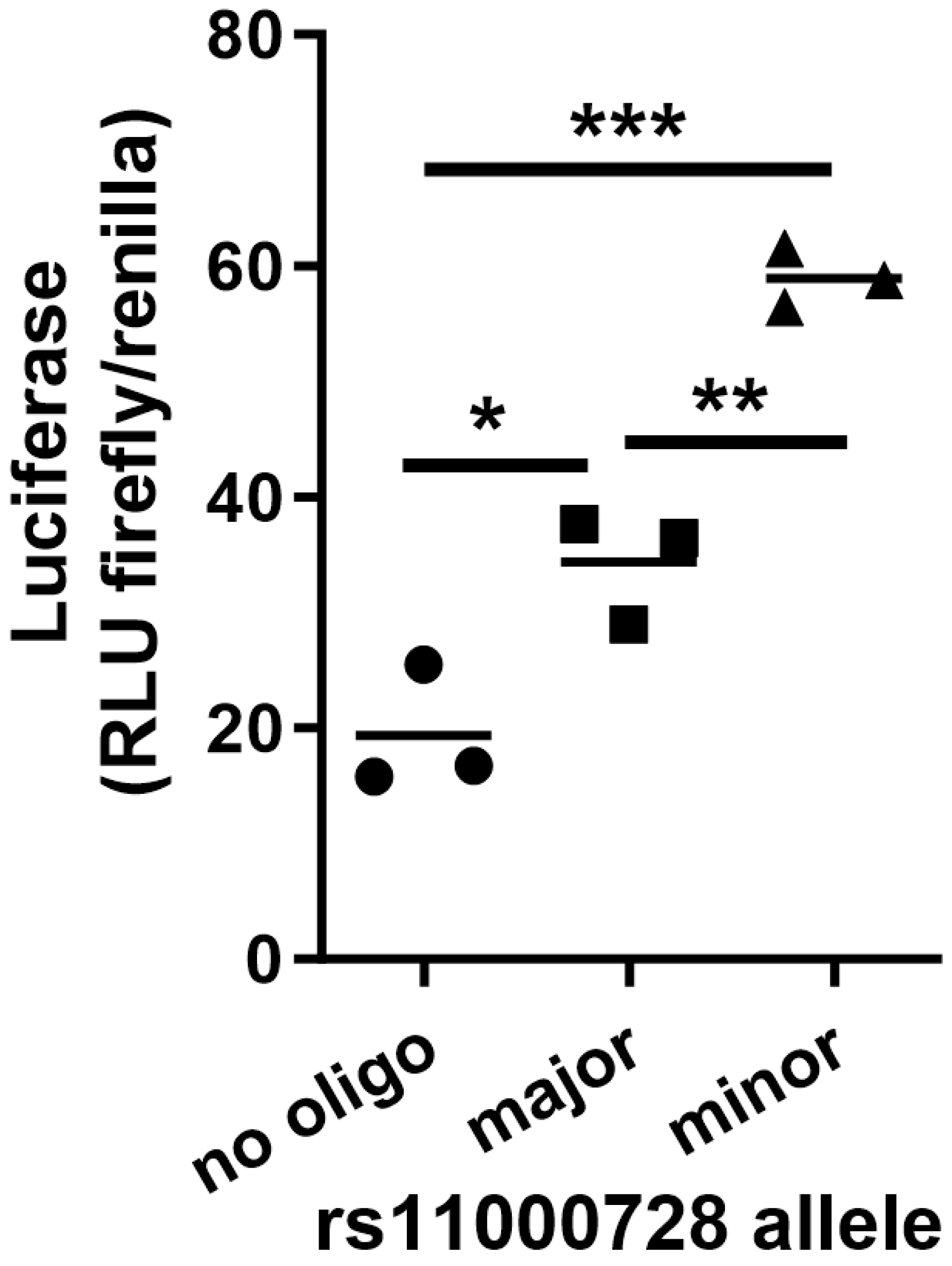
2.2. Identification of Putative Regulatory SNPs Controlling MYOZ1 and SYNPO2L Expression
2.3. Transient Expression of Fluorescently Tagged MYOZ1 and SYNPO2L Isoforms in iCMs
2.4. Left Atrial Coexpression with SYNPO2L or MYOZ1
3. Discussion
Limitations and Clinical Implications
4. Materials and Methods
4.1. RNAseq and Identification of Transcript Isoform-Specific eQTL Analysis
4.2. Protein Isoform Analysis
4.3. Identification of Putative Regulatory Regions
4.4. Reporter Gene Transfection of Human Pluripotent Stem Cell-Derived Cardiomyocytes
4.5. Protein Localization of Tagged MYOZ1 and CHAP Isoforms
4.6. Statistical Methods
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wolowacz, S.E.; Samuel, M.; Brennan, V.K.; Jasso-Mosqueda, J.G.; Van Gelder, I.C. The cost of illness of atrial fibrillation: A systematic review of the recent literature. Europace 2011, 13, 1375–1385. [Google Scholar] [CrossRef] [PubMed]
- Lippi, G.; Sanchis-Gomar, F.; Cervellin, G. Global epidemiology of atrial fibrillation: An increasing epidemic and public health challenge. Int. J. Stroke 2021, 16, 217–221. [Google Scholar] [CrossRef] [PubMed]
- Lavie, C.J.; Pandey, A.; Lau, D.H.; Alpert, M.A.; Sanders, P. Obesity and Atrial Fibrillation Prevalence, Pathogenesis, and Prognosis: Effects of Weight Loss and Exercise. J. Am. Coll. Cardiol. 2017, 70, 2022–2035. [Google Scholar] [CrossRef] [PubMed]
- Valembois, L.; Audureau, E.; Takeda, A.; Jarzebowski, W.; Belmin, J.; Lafuente-Lafuente, C. Antiarrhythmics for maintaining sinus rhythm after cardioversion of atrial fibrillation. Cochrane Database Syst. Rev. 2019, 9, CD005049. [Google Scholar] [CrossRef]
- Wass, S.Y.; Offerman, E.J.; Sun, H.; Hsu, J.; Rennison, J.H.; Cantlay, C.C.; McHale, M.L.; Gillinov, A.M.; Moravec, C.; Smith, J.D.; et al. Novel functional atrial fibrillation risk genes and pathways identified from coexpression analyses in human left atria. Heart Rhythm. 2023, 20, 1219–1226. [Google Scholar] [CrossRef]
- Roselli, C.; Chaffin, M.D.; Weng, L.C.; Aeschbacher, S.; Ahlberg, G.; Albert, C.M.; Almgren, P.; Alonso, A.; Anderson, C.D.; Aragam, K.G.; et al. Multi-ethnic genome-wide association study for atrial fibrillation. Nat. Genet. 2018, 50, 1225–1233. [Google Scholar] [CrossRef]
- Nielsen, J.B.; Thorolfsdottir, R.B.; Fritsche, L.G.; Zhou, W.; Skov, M.W.; Graham, S.E.; Herron, T.J.; McCarthy, S.; Schmidt, E.M.; Sveinbjornsson, G.; et al. Biobank-driven genomic discovery yields new insight into atrial fibrillation biology. Nat. Genet. 2018, 50, 1234–1239. [Google Scholar] [CrossRef]
- Hsu, J.; Gore-Panter, S.; Tchou, G.; Castel, L.; Lovano, B.; Moravec, C.S.; Pettersson, G.B.; Roselli, E.E.; Gillinov, A.M.; McCurry, K.R.; et al. Genetic Control of Left Atrial Gene Expression Yields Insights into the Genetic Susceptibility for Atrial Fibrillation. Circ. Genom. Precis. Med. 2018, 11, e002107. [Google Scholar] [CrossRef]
- Gontier, Y.; Taivainen, A.; Fontao, L.; Sonnenberg, A.; van der Flier, A.; Carpen, O.; Faulkner, G.; Borradori, L. The Z-disc proteins myotilin and FATZ-1 interact with each other and are connected to the sarcolemma via muscle-specific filamins. J. Cell Sci. 2005, 118, 3739–3749. [Google Scholar] [CrossRef]
- van Eldik, W.; Beqqali, A.; Monshouwer-Kloots, J.; Mummery, C.; Passier, R. Cytoskeletal heart-enriched actin-associated protein (CHAP) is expressed in striated and smooth muscle cells in chick and mouse during embryonic and adult stages. Int. J. Dev. Biol. 2011, 55, 649–655. [Google Scholar] [CrossRef]
- Beqqali, A.; Monshouwer-Kloots, J.; Monteiro, R.; Welling, M.; Bakkers, J.; Ehler, E.; Verkleij, A.; Mummery, C.; Passier, R. CHAP is a newly identified Z-disc protein essential for heart and skeletal muscle function. J. Cell Sci. 2010, 123, 1141–1150. [Google Scholar] [CrossRef] [PubMed]
- van Eldik, W.; den Adel, B.; Monshouwer-Kloots, J.; Salvatori, D.; Maas, S.; van der Made, I.; Creemers, E.E.; Frank, D.; Frey, N.; Boontje, N.; et al. Z-disc protein CHAPb induces cardiomyopathy and contractile dysfunction in the postnatal heart. PLoS ONE 2017, 12, e0189139. [Google Scholar] [CrossRef] [PubMed]
- Robinson, J.T.; Thorvaldsdóttir, H.; Winckler, W.; Guttman, M.; Lander, E.S.; Getz, G.; Mesirov, J.P. Integrative genomics viewer. Nat. Biotechnol. 2011, 29, 24–26. [Google Scholar] [CrossRef] [PubMed]
- Ward, L.D.; Kellis, M. HaploReg v4: Systematic mining of putative causal variants, cell types, regulators and target genes for human complex traits and disease. Nucleic Acids Res. 2016, 44, D877–D881. [Google Scholar] [CrossRef] [PubMed]
- Kundaje, A.; Meuleman, W.; Ernst, J.; Bilenky, M.; Yen, A.; Heravi-Moussavi, A.; Kheradpour, P.; Zhang, Z.; Wang, J.; Ziller, M.J.; et al. Integrative analysis of 111 reference human epigenomes. Nature 2015, 518, 317–330. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, W.; Zhu, W.; Dong, J.; Cheng, Y.; Yin, Z.; Shen, F. Mechanisms and Functions of Long Non-Coding RNAs at Multiple Regulatory Levels. Int. J. Mol. Sci. 2019, 20, 5573. [Google Scholar] [CrossRef]
- Gore-Panter, S.R.; Hsu, J.; Barnard, J.; Moravec, C.S.; Van Wagoner, D.R.; Chung, M.K.; Smith, J.D. PANCR, the PITX2 Adjacent Noncoding RNA, Is Expressed in Human Left Atria and Regulates PITX2c Expression. Circ. Arrhythm. Electrophysiol. 2016, 9, e003197. [Google Scholar] [CrossRef]
- Zhou, T.; Wu, Y.; Bi, Y.; Bai, H.; Jiang, Y.; Chen, G.; Chang, G.; Wang, Z. Gene Promotes Muscle Growth and Development in Meat Ducks. Genes 2022, 13, 1574. [Google Scholar] [CrossRef]
- Faulkner, G.; Pallavicini, A.; Comelli, A.; Salamon, M.; Bortoletto, G.; Ievolella, C.; Trevisan, S.; Kojic, S.; Dalla Vecchia, F.; Laveder, P.; et al. FATZ, a filamin-, actinin-, and telethonin-binding protein of the Z-disc of skeletal muscle. J. Biol. Chem. 2000, 275, 41234–41242. [Google Scholar] [CrossRef]
- Musarò, A.; McCullagh, K.J.; Naya, F.J.; Olson, E.N.; Rosenthal, N. IGF-1 induces skeletal myocyte hypertrophy through calcineurin in association with GATA-2 and NF-ATc1. Nature 1999, 400, 581–585. [Google Scholar] [CrossRef]
- Schulz, R.A.; Yutzey, K.E. Calcineurin signaling and NFAT activation in cardiovascular and skeletal muscle development. Dev. Biol. 2004, 266, 1–16. [Google Scholar] [CrossRef]
- Fiedler, B.; Lohmann, S.M.; Smolenski, A.; Linnemuller, S.; Pieske, B.; Schroder, F.; Molkentin, J.D.; Drexler, H.; Wollert, K.C. Inhibition of calcineurin-NFAT hypertrophy signaling by cGMP-dependent protein kinase type I in cardiac myocytes. Proc. Natl. Acad. Sci. USA 2002, 99, 11363–11368. [Google Scholar] [CrossRef] [PubMed]
- Bushdid, P.B.; Osinska, H.; Waclaw, R.R.; Molkentin, J.D.; Yutzey, K.E. NFATc3 and NFATc4 are required for cardiac development and mitochondrial function. Circ. Res. 2003, 92, 1305–1313. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.Y.; Kim, E.; Douglas, C.M.; Wirianto, M.; Han, C.; Ono, K.; Kim, S.Y.; Ji, J.H.; Tran, C.K.; Chen, Z.; et al. The circadian E3 ligase FBXL21 regulates myoblast differentiation and sarcomere architecture via MYOZ1 ubiquitination and NFAT signaling. PLoS Genet. 2022, 18, e1010574. [Google Scholar] [CrossRef] [PubMed]
- Yoo, S.H. Circadian regulation of cardiac muscle function and protein degradation. Chronobiol. Int. 2023, 40, 4–12. [Google Scholar] [CrossRef] [PubMed]
- Rabinovich-Nikitin, I.; Love, M.; Kirshenbaum, L.A. Intersection of autophagy regulation and circadian rhythms in the heart. Biochim. Biophys. Acta Mol. Basis Dis. 2022, 1868, 166354. [Google Scholar] [CrossRef]
- Wirianto, M.; Yang, J.; Kim, E.; Gao, S.; Paudel, K.R.; Choi, J.M.; Choe, J.; Gloston, G.F.; Ademoji, P.; Parakramaweera, R.; et al. The GSK-3β-FBXL21 Axis Contributes to Circadian TCAP Degradation and Skeletal Muscle Function. Cell Rep. 2020, 32, 108140. [Google Scholar] [CrossRef]
- Mirzoev, T.M.; Sharlo, K.A.; Shenkman, B.S. The Role of GSK-3β in the Regulation of Protein Turnover, Myosin Phenotype, and Oxidative Capacity in Skeletal Muscle under Disuse Conditions. Int. J. Mol. Sci. 2021, 22, 5081. [Google Scholar] [CrossRef]
- Nattel, S.; Li, D. Ionic remodeling in the heart: Pathophysiological significance and new therapeutic opportunities for atrial fibrillation. Circ. Res. 2000, 87, 440–447. [Google Scholar] [CrossRef]
- Pellman, J.; Lyon, R.C.; Sheikh, F. Extracellular matrix remodeling in atrial fibrosis: Mechanisms and implications in atrial fibrillation. J. Mol. Cell Cardiol. 2010, 48, 461–467. [Google Scholar] [CrossRef]
- Varabyou, A.; Pertea, G.; Pockrandt, C.; Pertea, M. TieBrush: An efficient method for aggregating and summarizing mapped reads across large datasets. Bioinformatics 2021, 37, 3650–3651. [Google Scholar] [CrossRef] [PubMed]
- Varabyou, A.; Sommer, M.J.; Erdogdu, B.; Shinder, I.; Minkin, I.; Chao, K.-H.; Park, S.; Heinz, J.; Pockrandt, C.; Shumate, A.; et al. CHESS 3: An improved, comprehensive catalog of human genes and transcripts based on large-scale expression data, phylogenetic analysis, and protein structure. Genome Biol. 2023, 24, 249. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, M.; Huber, W.; Pagès, H.; Aboyoun, P.; Carlson, M.; Gentleman, R.; Morgan, M.T.; Carey, V.J. Software for computing and annotating genomic ranges. PLoS Comput. Biol. 2013, 9, e1003118. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, S.; Das, S.; Kretzschmar, W.; Delaneau, O.; Wood, A.R.; Teumer, A.; Kang, H.M.; Fuchsberger, C.; Danecek, P.; Sharp, K.; et al. A reference panel of 64,976 haplotypes for genotype imputation. Nat. Genet. 2016, 48, 1279–1283. [Google Scholar] [CrossRef] [PubMed]
- Law, C.W.; Chen, Y.; Shi, W.; Smyth, G.K. voom: Precision weights unlock linear model analysis tools for RNA-seq read counts. Genome Biol. 2014, 15, R29. [Google Scholar] [CrossRef]
- Robinson, M.D.; Oshlack, A. A scaling normalization method for differential expression analysis of RNA-seq data. Genome Biol. 2010, 11, R25. [Google Scholar] [CrossRef]
- Leek, J.T. svaseq: Removing batch effects and other unwanted noise from sequencing data. Nucleic Acids Res. 2014, 42, e161. [Google Scholar] [CrossRef]
- Adzhubei, I.; Jordan, D.M.; Sunyaev, S.R. Predicting functional effect of human missense mutations using PolyPhen-2. Curr. Protoc. Hum. Genet. 2013, 7, 7.20.1–7.20.41. [Google Scholar] [CrossRef]
- Ward, L.D.; Kellis, M. HaploReg: A resource for exploring chromatin states, conservation, and regulatory motif alterations within sets of genetically linked variants. Nucleic Acids Res. 2012, 40, D930–D934. [Google Scholar] [CrossRef]
- Burridge, P.W.; Matsa, E.; Shukla, P.; Lin, Z.C.; Churko, J.M.; Ebert, A.D.; Lan, F.; Diecke, S.; Huber, B.; Mordwinkin, N.M.; et al. Chemically defined generation of human cardiomyocytes. Nat. Methods 2014, 11, 855–860. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Positive association with MYOZ1 coexpression | ||
| Rank | Hallmark | p-value |
| 1 | HALLMARK_CHOLESTEROL_HOMEOSTASIS | 2.24 × 10−2 |
| 2 | HALLMARK_UV_RESPONSE_UP | 3.09 × 10−2 |
| 3 | HALLMARK_MTORC1_SIGNALING | 3.39 × 10−2 |
| 4 | HALLMARK_PI3K_AKT_MTOR_SIGNALING | 3.55 × 10−2 |
| 5 | HALLMARK_DNA_REPAIR | 4.07 × 10−2 |
| Negative association with MYOZ1 coexpression | ||
| Rank | Hallmark | p-value |
| 1 | HALLMARK_OXIDATIVE_PHOSPHORYLATION | 8.13 × 10−3 |
| 2 | HALLMARK_APICAL_SURFACE | 2.95 × 10−2 |
| 3 | HALLMARK_EPITHELIAL_MESENCHYMAL_TRANSITION | 3.39 × 10−2 |
| 4 | HALLMARK_UV_RESPONSE_DN | 4.79 × 10−2 |
| Positive association with SYNPO2L coexpression | ||
| Rank | Hallmark | p-value |
| 1 | HALLMARK_OXIDATIVE_PHOSPHORYLATION | 1.20 × 10−3 |
| 2 | HALLMARK_MYOGENESIS | 2.69 × 10−3 |
| 3 | HALLMARK_ADIPOGENESIS | 1.55 × 10−2 |
| 4 | HALLMARK_FATTY_ACID_METABOLISM | 2.82 × 10−2 |
| 5 | HALLMARK_APICAL_JUNCTION | 3.39 × 10−2 |
| 6 | HALLMARK_UV_RESPONSE_DN | 3.47 × 10−2 |
| 7 | HALLMARK_EPITHELIAL_MESENCHYMAL_TRANSITION | 3.98 × 10−2 |
| 8 | HALLMARK_NOTCH_SIGNALING | 4.37 × 10−2 |
| 9 | HALLMARK_COAGULATION | 4.47 × 10−2 |
| 10 | HALLMARK_KRAS_SIGNALING_DN | 4.79 × 10−2 |
| Negative association with SYNPO2L coexpression | ||
| Rank | Hallmark | p-value |
| 1 | HALLMARK_CHOLESTEROL_HOMEOSTASIS | 1.20 × 10−2 |
| 2 | HALLMARK_INTERFERON_ALPHA_RESPONSE | 1.48 × 10−2 |
| 3 | HALLMARK_MTORC1_SIGNALING | 1.66 × 10−2 |
| 4 | HALLMARK_INTERFERON_GAMMA_RESPONSE | 2.57 × 10−2 |
| 5 | HALLMARK_SPERMATOGENESIS | 3.16 × 10−2 |
| 7 | HALLMARK_P53_PATHWAY | 3.55 × 10−2 |
| 6 | HALLMARK_DNA_REPAIR | 3.55 × 10−2 |
| 8 | HALLMARK_TNFA_SIGNALING_VIA_NFKB | 3.55 × 10−2 |
| 9 | HALLMARK_ALLOGRAFT_REJECTION | 3.63 × 10−2 |
| 10 | HALLMARK_INFLAMMATORY_RESPONSE | 3.80 × 10−2 |
| 11 | HALLMARK_MYC_TARGETS_V1 | 3.89 × 10−2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wass, S.Y.; Sun, H.; Tchou, G.; Liu, N.; Van Wagoner, D.R.; Chung, M.K.; Barnard, J.; Smith, J.D. Transcriptomic Insights into the Atrial Fibrillation Susceptibility Locus near the MYOZ1 and SYNPO2L Genes. Int. J. Mol. Sci. 2024, 25, 10309. https://doi.org/10.3390/ijms251910309
Wass SY, Sun H, Tchou G, Liu N, Van Wagoner DR, Chung MK, Barnard J, Smith JD. Transcriptomic Insights into the Atrial Fibrillation Susceptibility Locus near the MYOZ1 and SYNPO2L Genes. International Journal of Molecular Sciences. 2024; 25(19):10309. https://doi.org/10.3390/ijms251910309
Chicago/Turabian StyleWass, Sojin Y., Han Sun, Gregory Tchou, Nana Liu, David R. Van Wagoner, Mina K. Chung, John Barnard, and Jonathan D. Smith. 2024. "Transcriptomic Insights into the Atrial Fibrillation Susceptibility Locus near the MYOZ1 and SYNPO2L Genes" International Journal of Molecular Sciences 25, no. 19: 10309. https://doi.org/10.3390/ijms251910309