
Genome-Guided Identification of Surfactin-Producing Bacillus halotolerans AQ11M9 with Anti-Candida auris Potential

 , , , , , , ,
, , , , , , ,  and
and
Abstract
:1. Introduction
2. Results
2.1. Bacterial Strains with Anti-C. auris Activity
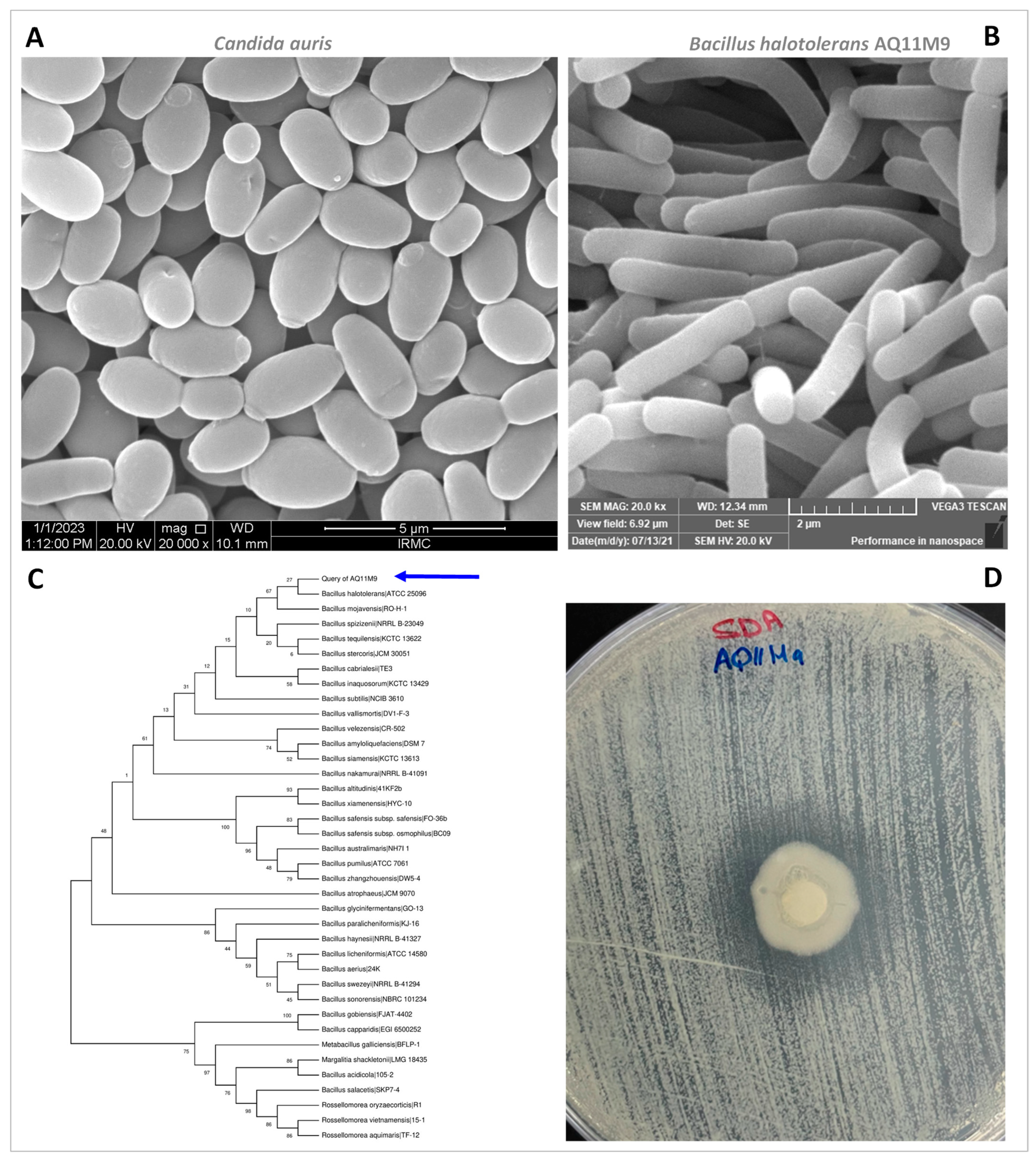
2.2. Structural Examination of AQ11M9 and C. auris Utilizing SEM
2.3. 16S rRNA Sequencing of AQ11M9
2.4. In Vitro Cytotoxicity
2.5. Identification of AQ11M9 by Whole Genome Sequence
2.6. Genome Analysis and Comparative Study
2.7. Secondary Metabolite and Biosynthetic Gene Detection
2.8. LC/MS Analysis of AQ11M9’s Secondary Metabolites
3. Discussion
4. Materials and Methods
4.1. Isolation of Bacteria
4.2. Anti-Candida auris Activity Screening
4.3. In Vitro Cytotoxic Study
4.4. Preparation of Bacillus Halotolerans for Scanning Electron Microscopy (SEM)
4.5. Preparation of Candida auris for SEM
4.6. 16S rRNA Gene Sequencing
4.7. Long-Read Whole Genome Sequencing
4.8. Genome Analysis
4.9. Biosynthetic Gene Cluster (BGC) Detection
4.10. Liquid Chromatography–Mass Spectrometry (LC/MS) Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Borgio, J.F.; Rasdan, A.S.; Sonbol, B.; Alhamid, G.; Almandil, N.B.; AbdulAzeez, S. Emerging Status of Multidrug-Resistant Bacteria and Fungi in the Arabian Peninsula. Biology 2021, 10, 1144. [Google Scholar] [CrossRef]
- Du, H.; Bing, J.; Nobile, C.J.; Huang, G. Candida auris infections in China. Virulence 2022, 13, 589–591. [Google Scholar] [CrossRef] [PubMed]
- Sanyaolu, A.; Okorie, C.; Marinkovic, A.; Abbasi, A.F.; Prakash, S.; Mangat, J.; Hosein, Z.; Haider, N.; Chan, J. Candida auris: An Overview of the Emerging Drug-Resistant Fungal Infection. Infect. Chemother. 2022, 54, 236–246. [Google Scholar] [CrossRef] [PubMed]
- Satoh, K.; Makimura, K.; Hasumi, Y.; Nishiyama, Y.; Uchida, K.; Yamaguchi, H. Candida auris sp. nov., a novel ascomycetous yeast isolated from the external ear canal of an inpatient in a Japanese hospital. Microbiol. Immunol. 2009, 53, 41–44, Erratum in Microbiol. Immunol. 2018, 62, 205. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, G.B.; Lorenz, A. What do we know about the biology of the emerging fungal pathogen of humans Candida auris? Microbiol. Res. 2021, 242, 126621. [Google Scholar] [CrossRef]
- Du, H.; Bing, J.; Hu, T.; Ennis, C.L.; Nobile, C.J.; Huang, G. Candida auris: Epidemiology, biology, antifungal resistance, and virulence. PLoS Pathog. 2020, 16, e1008921. [Google Scholar] [CrossRef]
- Thatchanamoorthy, N.; Devi, V.R.; Chandramathi, S.; Tay, S.T. Candida auris: A Mini Review on Epidemiology in Healthcare Facilities in Asia. J. Fungi 2022, 8, 1126. [Google Scholar] [CrossRef]
- Watkins, R.R.; Gowen, R.; Lionakis, M.; Ghannoum, M. Update on the Pathogenesis, Virulence, and Treatment of Candida auris. Pathog. Immun. 2022, 7, 46–65. [Google Scholar] [CrossRef]
- Aldossary, H.A.; Rehman, S.; Jermy, B.R.; AlJindan, R.; Aldayel, A.; AbdulAzeez, S.; Akhtar, S.; Alam Khan, F.; Borgio, J.F.; Al-Suhaimi, E.A. Therapeutic Intervention for Various Hospital Setting Strains of Biofilm Forming Candida auris with Multiple Drug Resistance Mutations Using Nanomaterial Ag-Silicalite-1 Zeolite. Pharmaceutics 2022, 14, 2251. [Google Scholar] [CrossRef]
- Izadi, A.; Gharehbolagh, S.A.; Sadeghi, F.; Talebi, M.; Darmiani, K.; Zarrinnia, A.; Zarei, F.; Peymaeei, F.; Khojasteh, S.; Borman, A.M.; et al. Drug repurposing against Candida auris: A systematic review. Mycoses 2022, 65, 784–793. [Google Scholar] [CrossRef]
- Borgio, J.F.; Almandil, N.B.; Selvaraj, P.; John, J.S.; Alquwaie, R.; AlHasani, E.; Alhur, N.F.; Aldahhan, R.; AlJindan, R.; Almohazey, D.; et al. The Potential of Dutasteride for Treating Multidrug-Resistant Candida auris Infection. Pharmaceutics 2024, 16, 810. [Google Scholar] [CrossRef] [PubMed]
- Berkow, E.L.; Angulo, D.; Lockhart, S.R. In Vitro Activity of a Novel Glucan Synthase Inhibitor, SCY-078, against Clinical Isolates of Candida auris. Antimicrob. Agents Chemother. 2017, 61. [Google Scholar] [CrossRef] [PubMed]
- Chu, S.; Long, L.; Sherif, R.; McCormick, T.S.; Borroto-Esoda, K.; Barat, S.; Ghannoum, M.A. A Second-Generation Fungerp Analog, SCY-247, Shows Potent In Vitro Activity against Candida auris and Other Clinically Relevant Fungal Isolates. Antimicrob. Agents Chemother. 2021, 65. [Google Scholar] [CrossRef] [PubMed]
- Shibata, T.; Takahashi, T.; Yamada, E.; Kimura, A.; Nishikawa, H.; Hayakawa, H.; Nomura, N.; Mitsuyama, J. T-2307 Causes Collapse of Mitochondrial Membrane Potential in Yeast. Antimicrob. Agents Chemother. 2012, 56, 5892–5897. [Google Scholar] [CrossRef] [PubMed]
- Wiederhold, N.P.; Najvar, L.K.; Jaramillo, R.; Olivo, M.; Patterson, H.; Connell, A.; Fukuda, Y.; Mitsuyama, J.; Catano, G.; Patterson, T.F. The Novel Arylamidine T-2307 Demonstrates In Vitro and In Vivo Activity against Candida auris. Antimicrob. Agents Chemother. 2020, 64. [Google Scholar] [CrossRef]
- Larkin, E.; Long, L.; Hager, C.; Shaw, K.J.; Ghannoum, M. Evaluation of the In Vitro and In Vivo Antifungal Activity of APX001A/APX001 against Candida auris. Open Forum Infect. Dis. 2017, 4, S471–S472. [Google Scholar] [CrossRef]
- Wiederhold, N.P.; Lockhart, S.R.; Najvar, L.K.; Berkow, E.L.; Jaramillo, R.; Olivo, M.; Garvey, E.P.; Yates, C.M.; Schotzinger, R.J.; Catano, G.; et al. The Fungal Cyp51-Specific Inhibitor VT-1598 Demonstrates In Vitro and In Vivo Activity against Candida auris. Antimicrob. Agents Chemother. 2019, 63. [Google Scholar] [CrossRef]
- Dong, J.; Liang, G.; Zheng, H.; Kan, S.; Song, N.; Zhang, M.; Liu, W. In Vitro activity of ravuconazole against Candida auris and vaginal candida isolates. Mycoses 2021, 64, 651–655. [Google Scholar] [CrossRef]
- Fernandes, L.; Ribeiro, R.; Henriques, M.; Rodrigues, M.E. Candida auris, a singular emergent pathogenic yeast: Its resistance and new therapeutic alternatives. Eur. J. Clin. Microbiol. Infect. Dis. 2022, 41, 1371–1385. [Google Scholar] [CrossRef]
- Tran, H.N.H.; Graham, L.; Adukwu, E.C. In vitro antifungal activity of Cinnamomum zeylanicum bark and leaf essential oils against Candida albicans and Candida auris. Appl. Microbiol. Biotechnol. 2020, 104, 8911–8924. [Google Scholar] [CrossRef]
- de Alteriis, E.; Maione, A.; Falanga, A.; Bellavita, R.; Galdiero, S.; Albarano, L.; Salvatore, M.M.; Galdiero, E.; Guida, M. Activity of Free and Liposome-Encapsulated Essential Oil from Lavandula angustifolia against Persister-Derived Biofilm of Candida auris. Antibiotics 2021, 11, 26. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Zhao, M.; Braun, D.R.; Ericksen, S.S.; Piotrowski, J.S.; Nelson, J.; Peng, J.; Ananiev, G.E.; Chanana, S.; Barns, K.; et al. A marine microbiome antifungal targets urgent-threat drug-resistant fungi. Science 2020, 370, 974–978. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Zhang, F.; Zarnowski, R.; Barns, K.J.; Jones, R.; Fossen, J.L.; Sanchez, H.; Rajski, S.R.; Audhya, A.; Bugni, T.S.; et al. Turbinmicin inhibits Candida biofilm growth by disrupting fungal vesicle–mediated trafficking. J. Clin. Investig. 2021, 131. [Google Scholar] [CrossRef] [PubMed]
- Barreto, T.L.; Rossato, L.; de Freitas, A.L.D.; Meis, J.F.; Lopes, L.B.; Colombo, A.L.; Ishida, K. Miltefosine as an alternative strategy in the treatment of the emerging fungus Candida auris. Int. J. Antimicrob. Agents 2020, 56, 106049. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.; Briffa, S.M.; Swingler, S.; Gibson, H.; Kannappan, V.; Adamus, G.; Kowalczuk, M.M.; Martin, C.; Radecka, I. Synthesis of Silver Nanoparticles Using Curcumin-Cyclodextrins Loaded into Bacterial Cellulose-Based Hydrogels for Wound Dressing Applications. Biomacromolecules 2020, 21, 1802–1811. [Google Scholar] [CrossRef]
- Piedrahita, C.T.; Cadnum, J.L.; Jencson, A.L.; Shaikh, A.A.; Ghannoum, M.A.; Donskey, C.J. Environmental Surfaces in Healthcare Facilities are a Potential Source for Transmission of Candida auris and Other Candida Species. Infect. Control. Hosp. Epidemiol. 2017, 38, 1107–1109. [Google Scholar] [CrossRef]
- Fu, L.; Le, T.; Liu, Z.; Wang, L.; Guo, H.; Yang, J.; Chen, Q.; Hu, J. Different efficacies of common disinfection methods against Candida auris and other candida species. J. Infect. Public Health 2020, 13, 730–736. [Google Scholar] [CrossRef]
- dos Santos, C.I.; Campos, C.D.L.; Nunes-Neto, W.R.; Carmo, M.S.D.; Nogueira, F.A.B.; Ferreira, R.M.; Costa, E.P.S.; Gonzaga, L.F.; Araújo, J.M.M.; Monteiro, J.M.; et al. Genomic Analysis of Limosilactobacillus fermentum ATCC 23271, a Potential Probiotic Strain with Anti-Candida Activity. J. Fungi 2021, 7, 794. [Google Scholar] [CrossRef]
- Parolin, C.; Marangoni, A.; Laghi, L.; Foschi, C.; Ñahui Palomino, R.A.; Calonghi, N.; Cevenini, R.; Vitali, B. Isolation of Vaginal Lactobacilli and Characterization of Anti-Candida Activity. PLoS ONE 2015, 10, e0131220. [Google Scholar] [CrossRef]
- Li, Y.; Li, X.; Jia, D.; Liu, J.; Wang, J.; Liu, A.; Liu, Z.; Guan, G.; Liu, G.; Luo, J.; et al. Complete genome sequence and antimicrobial activity of Bacillus velezensis JT3-1, a microbial germicide isolated from yak feces. 3 Biotech 2020, 10, 1–10. [Google Scholar] [CrossRef]
- Siupka, P.; Hansen, F.T.; Schier, A.; Rocco, S.; Sørensen, T.; Piotrowska-Seget, Z. Antifungal Activity and Biosynthetic Potential of New Streptomyces sp. MW-W600-10 Strain Isolated from Coal Mine Water. Int. J. Mol. Sci. 2021, 22, 7441. [Google Scholar] [CrossRef] [PubMed]
- Gosse, J.T.; Ghosh, S.; Sproule, A.; Overy, D.; Cheeptham, N.; Boddy, C.N. Whole Genome Sequencing and Metabolomic Study of Cave Streptomyces Isolates ICC1 and ICC. Front. Microbiol. 2019, 10, 1020. [Google Scholar] [CrossRef] [PubMed]
- Williams, A.N.; Sorout, N.; Cameron, A.J.; Stavrinides, J. The Integration of Genome Mining, Comparative Genomics, and Functional Genetics for Biosynthetic Gene Cluster Identification. Front. Genet. 2020, 11, 600116. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Saitou, A.; Wang, C.-M.; Toyoda, A.; Minakuchi, Y.; Sekiguchi, Y.; Ueda, K.; Takano, H.; Sakai, Y.; Abe, K.; et al. Genome Features and Secondary Metabolites Biosynthetic Potential of the Class Ktedonobacteria. Front. Microbiol. 2019, 10, 893. [Google Scholar] [CrossRef]
- Yadav, A.; Singh, A.; Chowdhary, A. Isolation of Candida auris in Clinical Specimens. Methods Mol. Biol. 2022, 2517, 3–20. [Google Scholar] [CrossRef] [PubMed]
- Hager, C.L.; Larkin, E.L.; Long, L.; Abidi, F.Z.; Shaw, K.J.; Ghannoum, M.A. In Vitro and In Vivo Evaluation of the Antifungal Activity of APX001A/APX001 against Candida auris. Antimicrob. Agents Chemother. 2018, 62. [Google Scholar] [CrossRef]
- Rossatto, F.C.P.; Tharmalingam, N.; Escobar, I.E.; D’azevedo, P.A.; Zimmer, K.R.; Mylonakis, E. Antifungal Activity of the Phenolic Compounds Ellagic Acid (EA) and Caffeic Acid Phenethyl Ester (CAPE) against Drug-Resistant Candida auris. J. Fungi 2021, 7, 763. [Google Scholar] [CrossRef]
- Marena, G.D.; Carvalho, G.C.; Ramos, M.A.d.S.; Chorilli, M.; Bauab, T.M. Anti-Candida auris activity in vitro and in vivo of micafungin loaded nanoemulsions. Med Mycol. 2022, 61, myac090. [Google Scholar] [CrossRef]
- Marena, G.D.; Ramos, M.A.D.S.; Lima, L.C.; Chorilli, M.; Bauab, T.M. Galleria mellonella for systemic assessment of anti-Candida auris using amphotericin B loaded in nanoemulsion. Sci. Total Environ. 2021, 807, 151023. [Google Scholar] [CrossRef]
- Cruz, A.L.; de Souza, F.S.; Abegg, M.A. Anti-Candida and anti-quorum sensing activity of airborne microorganisms detected by a rapid method. Rev. Soc. Bras. Med. Trop. 2019, 52, e20180152. [Google Scholar] [CrossRef]
- Pajor, M.; Worobo, R.W.; Milewski, S.; Szweda, P. The Antimicrobial Potential of Bacteria Isolated from Honey Samples Produced in the Apiaries Located in Pomeranian Voivodeship in Northern Poland. Int. J. Environ. Res. Public Health 2018, 15, 2002. [Google Scholar] [CrossRef] [PubMed]
- Parolin, C.; Croatti, V.; Laghi, L.; Giordani, B.; Tondi, M.R.; De Gregorio, P.R.; Foschi, C.; Vitali, B. Lactobacillus Biofilms Influence Anti-Candida Activity. Front. Microbiol. 2021, 12, 750368. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Srivastava, S. Anti- Candida activity of spent culture filtrate of Lactobacillus plantarum strain LR/14. J. Med. Mycol. 2013, 24, e25–e34. [Google Scholar] [CrossRef]
- Srivastava, V.; Dubey, A.K. Anti-biofilm activity of the metabolites of Streptomyces chrestomyceticus strain ADP4 against Candida albicans. J. Biosci. Bioeng. 2016, 122, 434–440. [Google Scholar] [CrossRef] [PubMed]
- Lakhtin, M.; Alyoshkin, V.; Lakhtin, V.; Afanasyev, S.; Pozhalostina, L.; Pospelova, V. Probiotic Lactobacillus and Bifidobacterial Lectins against Candida albicans and Staphylococcus aureus Clinical Strains: New Class of the Pathogen Biofilm Destructors. Probiotics Antimicrob. Proteins 2010, 2, 186–196. [Google Scholar] [CrossRef]
- Lipinska-Zubrycka, L.; Klewicki, R.; Sojka, M.; Bonikowski, R.; Milczarek, A.; Klewicka, E. Anticandidal activity of Lactobacillus spp. in the presence of galactosyl polyols. Microbiol. Res. 2020, 240, 126540. [Google Scholar] [CrossRef]
- Okkers, D.J.; Dicks, L.M.T.; Silvester, M.; Joubert, J.J.; Odendaal, H.J. Characterization of pentocin TV35b, a bacteriocin-like peptide isolated from Lactobacillus pentosus with a fungistatic effect on Candida albicans. J. Appl. Microbiol. 1999, 87, 726–734. [Google Scholar] [CrossRef]
- Vazquez-Munoz, R.; Dongari-Bagtzoglou, A. Anticandidal Activities by Lactobacillus Species: An Update on Mechanisms of Action. Front. Oral Health 2021, 2, 689382. [Google Scholar] [CrossRef]
- Dunlap, C.A.; Bowman, M.J.; Schisler, D.A.; Rooney, A.P. Genome analysis shows Bacillus axarquiensis is not a later heterotypic synonym of Bacillus mojavensis; reclassification of Bacillus malacitensis and Brevibacterium halotolerans as heterotypic synonyms of Bacillus axarquiensis. Int. J. Syst. Evol. Microbiol. 2016, 66, 2438–2443. [Google Scholar] [CrossRef]
- Sagredo-Beltrán, J.; De La Cruz-Rodríguez, Y.; Alvarado-Rodríguez, M.; Vega-Arreguín, J.; Rodríguez-Guerra, R.; Alvarado-Gutiérrez, A.; Fraire-Velázquez, S. Genome Sequence of Bacillus halotolerans Strain MS50-18A with Antifungal Activity against Phytopathogens, Isolated from Saline Soil in San Luis Potosi, Mexico. Genome Announc. 2018, 6. [Google Scholar] [CrossRef]
- Ben Slama, H.; Cherif-Silini, H.; Bouket, A.C.; Qader, M.; Silini, A.; Yahiaoui, B.; Alenezi, F.N.; Luptakova, L.; Triki, M.A.; Vallat, A.; et al. Screening for Fusarium Antagonistic Bacteria from Contrasting Niches Designated the Endophyte Bacillus halotolerans as Plant Warden against Fusarium. Front. Microbiol. 2019, 9, 3236. [Google Scholar] [CrossRef]
- Thomloudi, E.-E.; Tsalgatidou, P.C.; Baira, E.; Papadimitriou, K.; Venieraki, A.; Katinakis, P. Genomic and Metabolomic Insights into Secondary Metabolites of the Novel Bacillus halotolerans Hil4, an Endophyte with Promising Antagonistic Activity against Gray Mold and Plant Growth Promoting Potential. Microorganisms 2021, 9, 2508. [Google Scholar] [CrossRef] [PubMed]
- Tsalgatidou, P.C.; Thomloudi, E.-E.; Baira, E.; Papadimitriou, K.; Skagia, A.; Venieraki, A.; Katinakis, P. Integrated Genomic and Metabolomic Analysis Illuminates Key Secreted Metabolites Produced by the Novel Endophyte Bacillus halotolerans Cal.l.30 Involved in Diverse Biological Control Activities. Microorganisms 2022, 10, 399. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Xiao, J.; Zhang, Y.; Li, R.; Liu, L.; Deng, J. Biocontrol ability and action mechanism of Bacillus halotolerans against Botrytis cinerea causing grey mould in postharvest strawberry fruit. Postharvest Biol. Technol. 2021, 174, 111456. [Google Scholar] [CrossRef]
- Wu, X.; Wu, H.; Wang, R.; Wang, Z.; Zhang, Y.; Gu, Q.; Farzand, A.; Yang, X.; Semenov, M.; Borriss, R.; et al. Genomic Features and Molecular Function of a Novel Stress-Tolerant Bacillus halotolerans Strain Isolated from an Extreme Environment. Biology 2021, 10, 1030. [Google Scholar] [CrossRef]
- Amoutzias, G.D.; Nikolaidis, M.; Hesketh, A. The Notable Achievements and the Prospects of Bacterial Pathogen Genomics. Microorganisms 2022, 10, 1040. [Google Scholar] [CrossRef]
- Deamer, D.; Akeson, M.; Branton, D. Three decades of nanopore sequencing. Nat. Biotechnol. 2016, 34, 518–524. [Google Scholar] [CrossRef] [PubMed]
- Jain, M.; Olsen, H.E.; Paten, B.; Akeson, M. The Oxford Nanopore MinION: Delivery of nanopore sequencing to the genomics community. Genome Biol. 2016, 17, 1–11. [Google Scholar] [CrossRef]
- Wang, Y.; Zhao, Y.; Bollas, A.; Wang, Y.; Au, K.F. Nanopore sequencing technology, bioinformatics and applications. Nat. Biotechnol. 2021, 39, 1348–1365. [Google Scholar] [CrossRef]
- Tedersoo, L.; Albertsen, M.; Anslan, S.; Callahan, B. Perspectives and Benefits of High-Throughput Long-Read Sequencing in Microbial Ecology. Appl. Environ. Microbiol. 2021, 87, e00626-21. [Google Scholar] [CrossRef]
- Method of the Year 2022: Long-read sequencing. Nat. Methods 2023, 20, 1. [CrossRef] [PubMed]
- Marshall, A.; Fuller, D.T.; Dougall, P.; Kumaragama, K.; Dhaniyala, S.; Sur, S. Application of Nanopore Sequencing for Accurate Identification of Bacterial Colonies. BioRxiv 2023. [Google Scholar] [CrossRef]
- Blin, K.; Shaw, S.; Kloosterman, A.M.; Charlop-Powers, Z.; van Wezel, G.P.; Medema, M.H.; Weber, T. antiSMASH 6.0: Improving cluster detection and comparison capabilities. Nucleic Acids Res. 2021, 49, W29–W35. [Google Scholar] [CrossRef] [PubMed]
- Honma, M.; Tanaka, K.; Konno, K.; Tsuge, K.; Okuno, T.; Hashimoto, M. Termination of the structural confusion between plipastatin A1 and fengycin IX. Bioorganic Med. Chem. 2012, 20, 3793–3798. [Google Scholar] [CrossRef]
- Hussein, W. Fengycin or plipastatin? A confusing question in Bacilli. BioTechnologia 2019, 100, 47–55. [Google Scholar] [CrossRef]
- Liu, L.; Jin, X.; Lu, X.; Guo, L.; Lu, P.; Yu, H.; Lv, B. Mechanisms of Surfactin from Bacillus subtilis SF1 against Fusarium foetens: A Novel Pathogen Inducing Potato Wilt. J. Fungi 2023, 9, 367. [Google Scholar] [CrossRef]
- AlJindan, R.; AlEraky, D.M.; Mahmoud, N.; Abdalhamid, B.; Almustafa, M.; AbdulAzeez, S.; Borgio, J.F. Drug Resistance-Associated Mutations in ERG11 of Multidrug-Resistant Candida auris in a Tertiary Care Hospital of Eastern Saudi Arabia. J. Fungi 2020, 7, 18. [Google Scholar] [CrossRef]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Collins, T.J. ImageJ for microscopy. Biotechniques 2007, 43 (Suppl. 1), S25–S30. [Google Scholar] [CrossRef]
- Borgio, J.F.; Alhujaily, R.; Alquwaie, R.; Alabdullah, M.J.; AlHasani, E.; Alothman, W.; Alaqeel, R.K.; Alfaraj, A.S.; Kaabi, A.; Alhur, N.F.; et al. Mining the nanotube-forming Bacillus amyloliquefaciens MR14M3 genome for determining anti-Candida auris and anti-Candida albicans potential by pathogenicity and comparative genomics analysis. Comput. Struct. Biotechnol. J. 2023, 21, 4261–4276. [Google Scholar] [CrossRef]
- AlEraky, D.M.; Madi, M.; El Tantawi, M.; AlHumaid, J.; Fita, S.; AbdulAzeez, S.; Borgio, J.F.; Al-Harbi, F.A.; Alagl, A.S. Predominance of non-Streptococcus mutans bacteria in dental biofilm and its relation to caries progression. Saudi J. Biol. Sci. 2021, 28, 7390–7395. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.-H.; Ha, S.-M.; Kwon, S.; Lim, J.; Kim, Y.; Seo, H.; Chun, J. Introducing EzBioCloud: A taxonomically united database of 16S rRNA gene sequences and whole-genome assemblies. Int. J. Syst. Evol. Microbiol. 2017, 67, 1613–1617. [Google Scholar] [CrossRef]
- AlJindan, R.; Mahmoud, N.; AlEraky, D.M.; Almandil, N.B.; AbdulAzeez, S.; Borgio, J.F. Phenomics and genomic features of Enterococcus avium IRMC1622a isolated from a clinical sample of hospitalized patient. J. Infect. Public Health 2024, 17, 102463. [Google Scholar] [CrossRef]
- Czmil, A.; Wronski, M.; Czmil, S.; Sochacka-Pietal, M.; Cmil, M.; Gawor, J.; Wołkowicz, T.; Plewczynski, D.; Strzalka, D.; Pietal, M. NanoForms: An integrated server for processing, analysis and assembly of raw sequencing data of microbial genomes, from Oxford Nanopore technology. PeerJ 2022, 10, e13056. [Google Scholar] [CrossRef]
- de Koning, W.; Miladi, M.; Hiltemann, S.; Heikema, A.; Hays, J.P.; Flemming, S.; Beek, M.v.D.; Mustafa, D.A.; Backofen, R.; Grüning, B.; et al. NanoGalaxy: Nanopore long-read sequencing data analysis in Galaxy. GigaScience 2020, 9, giaa105. [Google Scholar] [CrossRef]
- Meier-Kolthoff, J.P.; Göker, M. TYGS is an automated high-throughput platform for state-of-the-art genome-based taxonomy. Nat. Commun. 2019, 10, 2182. [Google Scholar] [CrossRef] [PubMed]
- Wattam, A.R.; Davis, J.J.; Assaf, R.; Boisvert, S.; Brettin, T.; Bun, C.; Conrad, N.; Dietrich, E.M.; Disz, T.; Gabbard, J.L.; et al. Improvements to PATRIC, the all-bacterial Bioinformatics Database and Analysis Resource Center. Nucleic Acids Res. 2016, 45, D535–D542. [Google Scholar] [CrossRef]
- Antonopoulos, D.A.; Assaf, R.; Aziz, R.K.; Brettin, T.; Bun, C.; Conrad, N.; Davis, J.J.; Dietrich, E.M.; Disz, T.; Gerdes, S.; et al. PATRIC as a unique resource for studying antimicrobial resistance. Briefings Bioinform. 2017, 20, 1094–1102. [Google Scholar] [CrossRef]
- Terlouw, B.R.; Blin, K.; Navarro-Muñoz, J.C.; Avalon, N.E.; Chevrette, M.G.; Egbert, S.; Lee, S.; Meijer, D.; Recchia, M.J.J.; Reitz, Z.L.; et al. MIBiG 3.0: A community-driven effort to annotate experimentally validated biosynthetic gene clusters. Nucleic Acids Res. 2022, 51, D603–D610. [Google Scholar] [CrossRef]
- Klau, L.J.; Podell, S.; Creamer, K.E.; Demko, A.M.; Singh, H.W.; Allen, E.E.; Moore, B.S.; Ziemert, N.; Letzel, A.C.; Jensen, P.R. The Natural Product Domain Seeker version 2 (NaPDoS2) webtool relates ketosynthase phylogeny to biosynthetic function. J. Biol. Chem. 2022, 298, 102480. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Features | AQ11M9 |
|---|---|
| Genome size | 4,197,347 bp |
| The quantity of protein coding sequences | 5623 |
| The quantity of rRNA genes | 30 |
| The quantity of tRNA genes | 86 |
| The quantity of tmRNA genes | 1 |
| GC content % | 43.73 |
| Hypothetical proteins | 894 |
| Region | Predicted Size (bp) | Type of BGC | Most Similar Known Cluster | No. of Genes | MIBIG ID/(Similarity %) |
|---|---|---|---|---|---|
| 1.1 | 62,838 | NRP: LIPOPEPTIDE | surfactin | 60 | BGC0000433/86% |
| 1.2 | 10,387 | RiPP-LIKE | unknown | 16 | |
| 1.3 | 41,208 | OTHER | bacilysin | 53 | BGC0001184/100% |
| 1.4 | 21,615 | Sactipeptide, RiPP: Thiopeptide | subtilosin | 29 | BGC0000602/87% |
| 1.5 | 44,322 | NRPS | bacillibactin | 43 | BGC0000309/100% |
| NRPS | paenibactin | BGC0000401/100% | |||
| 1.6 | 39,542 | T3PKS | unknown | 60 | |
| 1.7 | 20,225 | TERPENE | unknown | 22 | |
| 1.8 | 73,995 | NRPS, BETALACTONE | fengycin | 62 | BGC0001095/100% |
| NRP | plipastatin | BGC0000407/100% | |||
| NRP + polyketide | mycosubtilin | BGC0001103/90% | |||
| 1.9 | 87,945 | POLYKETIDE + NRP | bacillaene | 57 | BGC0001089/92% |
| Polyketide: Trans-AT type I | macrobrevin | BGC0001470/93% | |||
| 1.10 | 20,807 | TERPENE | unknown | 25 |
| Metabolite Type | Most Similar Known Cluster | AQ11M9 | PAMC26543 | Halotolerans v48-19 |
|---|---|---|---|---|
| NRPS: Lipopeptide | surfactin | 86% | 86% | |
| RiPP-LIKE | unknown | + | − | − |
| OTHER | bacilysin | 100% | 100% | 100% |
| SACTIPEPTIDE | Subtilosin A | 87% | 100% | 100% |
| NRPS | bacillibactin | 100% | 100% | |
| NRPS | paenibactin | 100% | 100% | |
| T3PKS | unknown | + | − | − |
| TERPENE | unknown | + | − | − |
| NRPS, BETALACTONE | fengycin | 100% | 100% | |
| NRP | plipastatin | 100% | 100% | |
| Polyketide + NRP | bacillaene | 92% | 100% | |
| Polyketide: Trans-AT type I | macrobrevin | 93% | 53% | |
| TERPENE | unknown | + | − | − |
| bacteriocin | unknown | + | − | − |
| BGC | NRP/Polyketide | Identity % | Align Length | E-Value | Domain Class |
|---|---|---|---|---|---|
| Surfactin | NRP | 92 | 426 | 2.1 × 10−231 | starter |
| 91 | 433 | 2.1 × 10−231 | DCL | ||
| 89 | 445 | 2 × 10−238 | epim | ||
| Bacilibactin | NRP | 86 | 305 | 2.5 × 10−152 | starter |
| 88 | 446 | 2 × 10−229 | LCL | ||
| Fengycin | NRP | 100 | 290 | 4.00 × 10−169 | DCL |
| 98 | 436 | 4.50 × 10−250 | starter | ||
| 98 | 429 | 8.20 × 10−247 | LCL | ||
| 97 | 456 | 1.10 × 10−264 | epim | ||
| Bacillaene | NRP/PK | 88 | 418 | 1.20 × 10−220 | Type I trans-AT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Borgio, J.F.; Alhujaily, R.; Alfaraj, A.S.; Alabdullah, M.J.; Alaqeel, R.K.; Kaabi, A.; Alquwaie, R.; Alhur, N.F.; AlJindan, R.; Almofty, S.; et al. Genome-Guided Identification of Surfactin-Producing Bacillus halotolerans AQ11M9 with Anti-Candida auris Potential. Int. J. Mol. Sci. 2024, 25, 10408. https://doi.org/10.3390/ijms251910408
Borgio JF, Alhujaily R, Alfaraj AS, Alabdullah MJ, Alaqeel RK, Kaabi A, Alquwaie R, Alhur NF, AlJindan R, Almofty S, et al. Genome-Guided Identification of Surfactin-Producing Bacillus halotolerans AQ11M9 with Anti-Candida auris Potential. International Journal of Molecular Sciences. 2024; 25(19):10408. https://doi.org/10.3390/ijms251910408
Chicago/Turabian StyleBorgio, J. Francis, Rahaf Alhujaily, Aqeelah Salman Alfaraj, Maryam Jawad Alabdullah, Rawan Khalid Alaqeel, Ayidah Kaabi, Rahaf Alquwaie, Norah F. Alhur, Reem AlJindan, Sarah Almofty, and et al. 2024. "Genome-Guided Identification of Surfactin-Producing Bacillus halotolerans AQ11M9 with Anti-Candida auris Potential" International Journal of Molecular Sciences 25, no. 19: 10408. https://doi.org/10.3390/ijms251910408