

Identification of Key Immune and Cell Cycle Modules and Prognostic Genes for Glioma Patients through Transcriptome Analysis

 ,
,
Abstract
:1. Introduction
2. Results
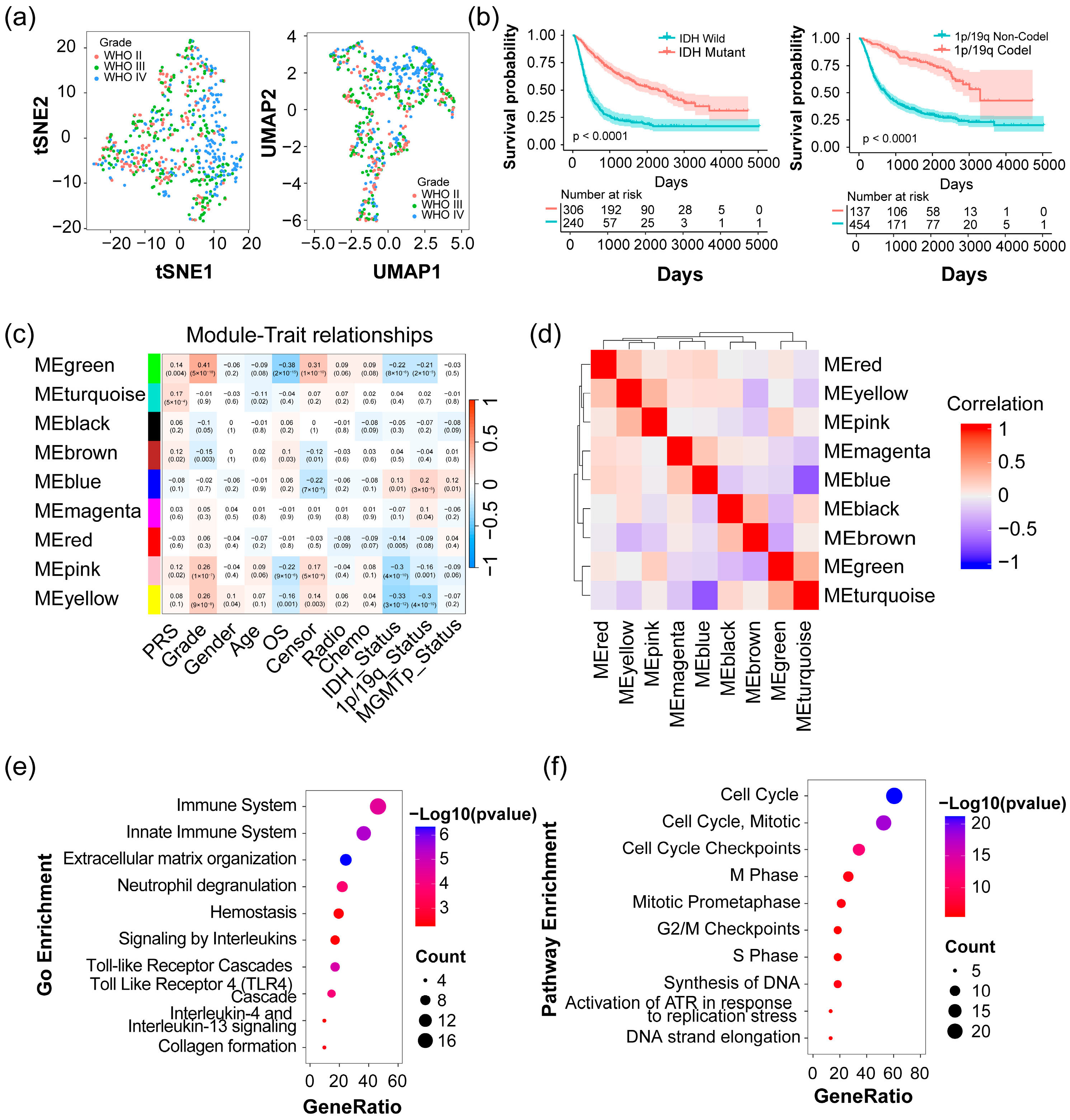
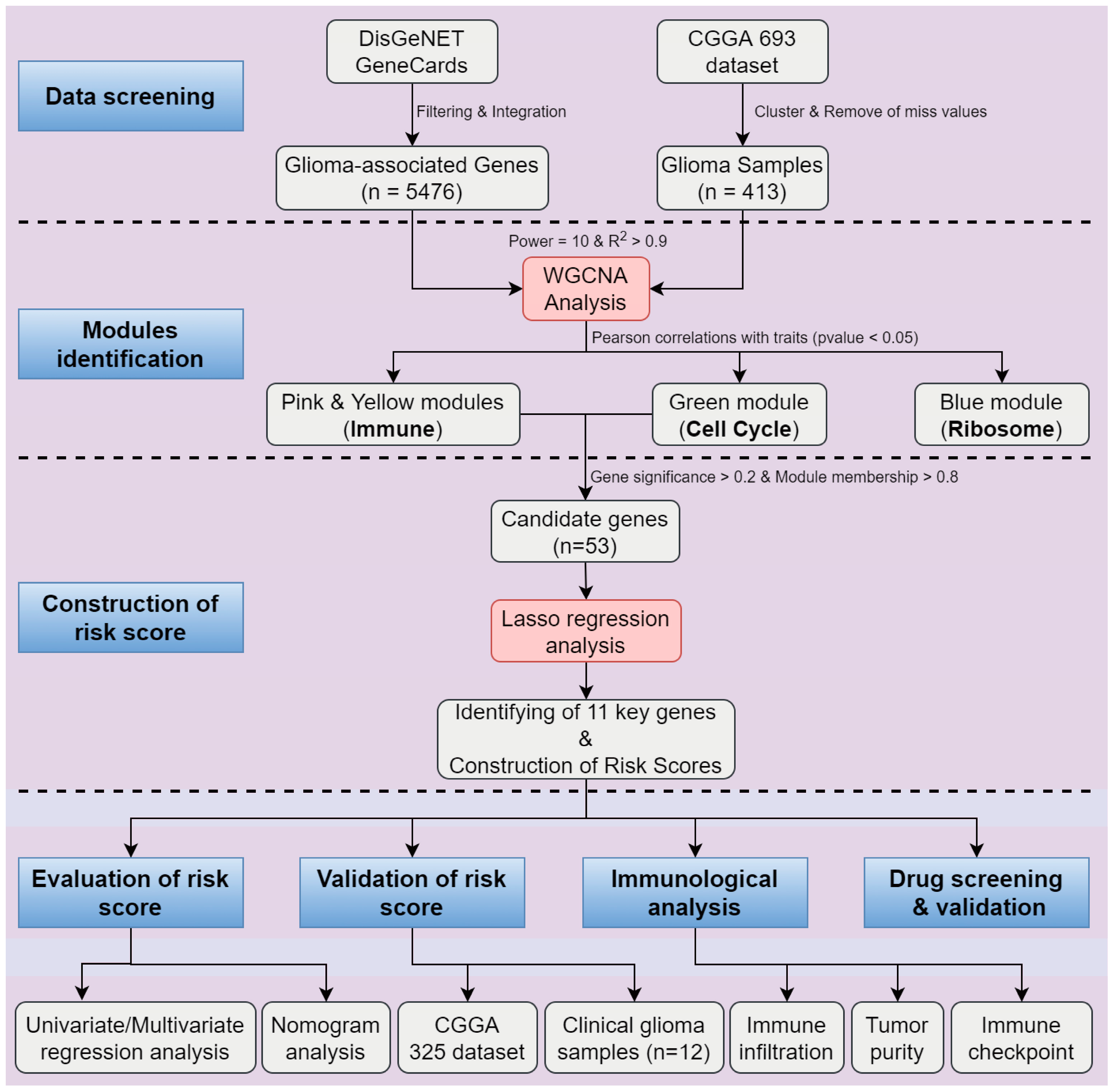
2.1. Identification of Four Modules Significantly Associated with Clinical Traits in Glioma
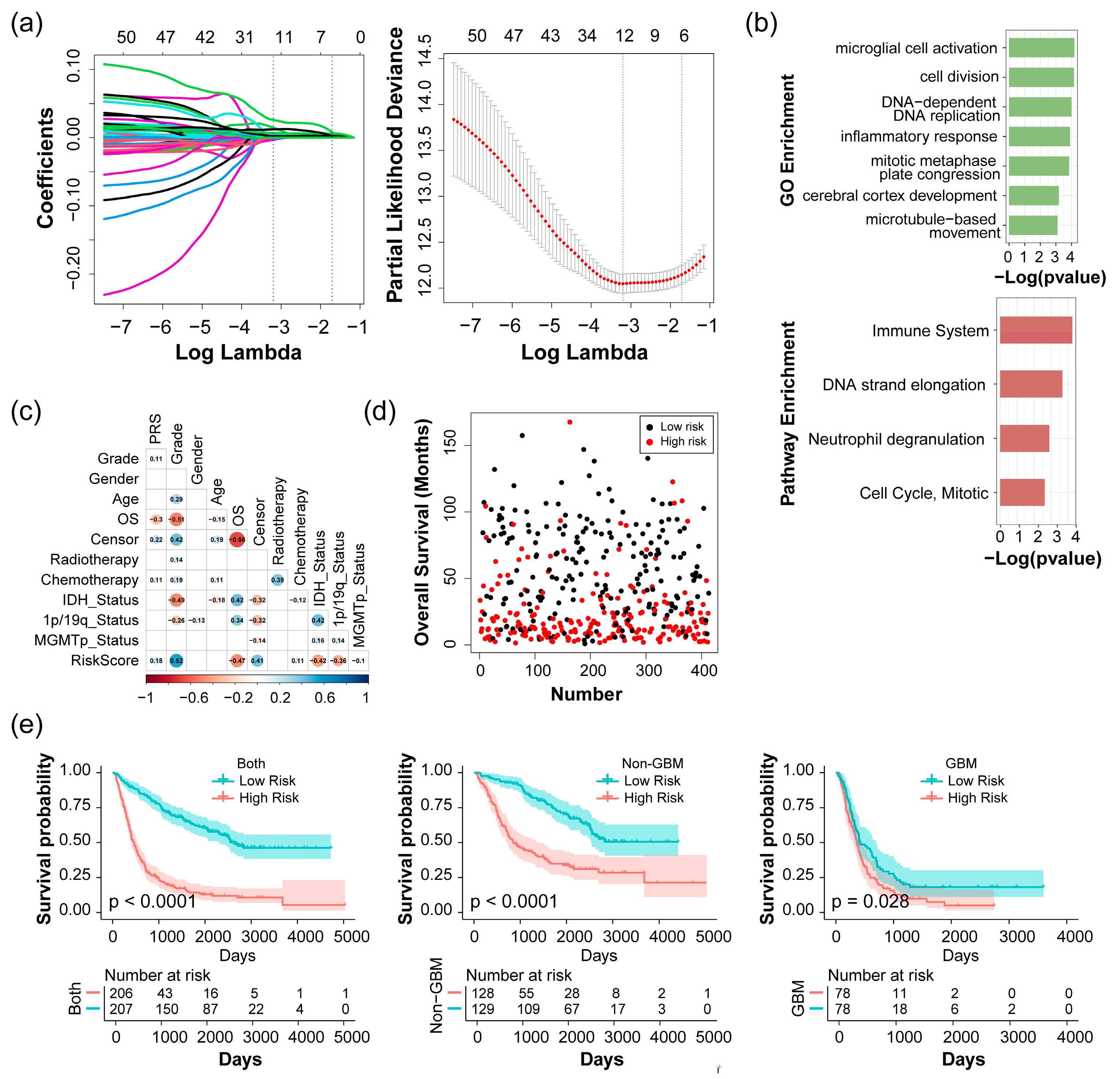
2.2. Construction and Evaluation of Risk Score with the CGGA 693 Dataset
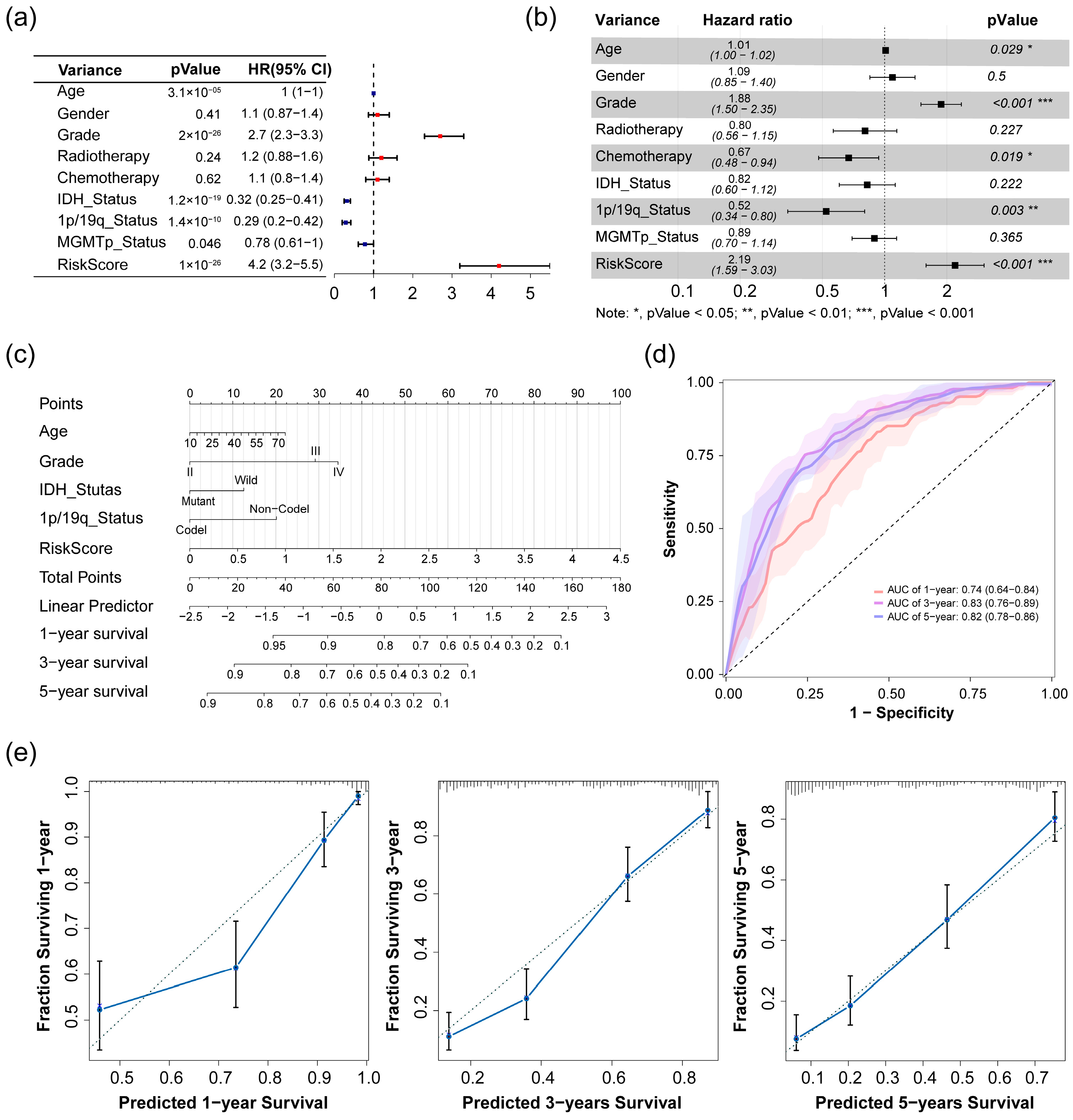
2.3. Analysis of Risk Score as an Independent Prognostic Signature
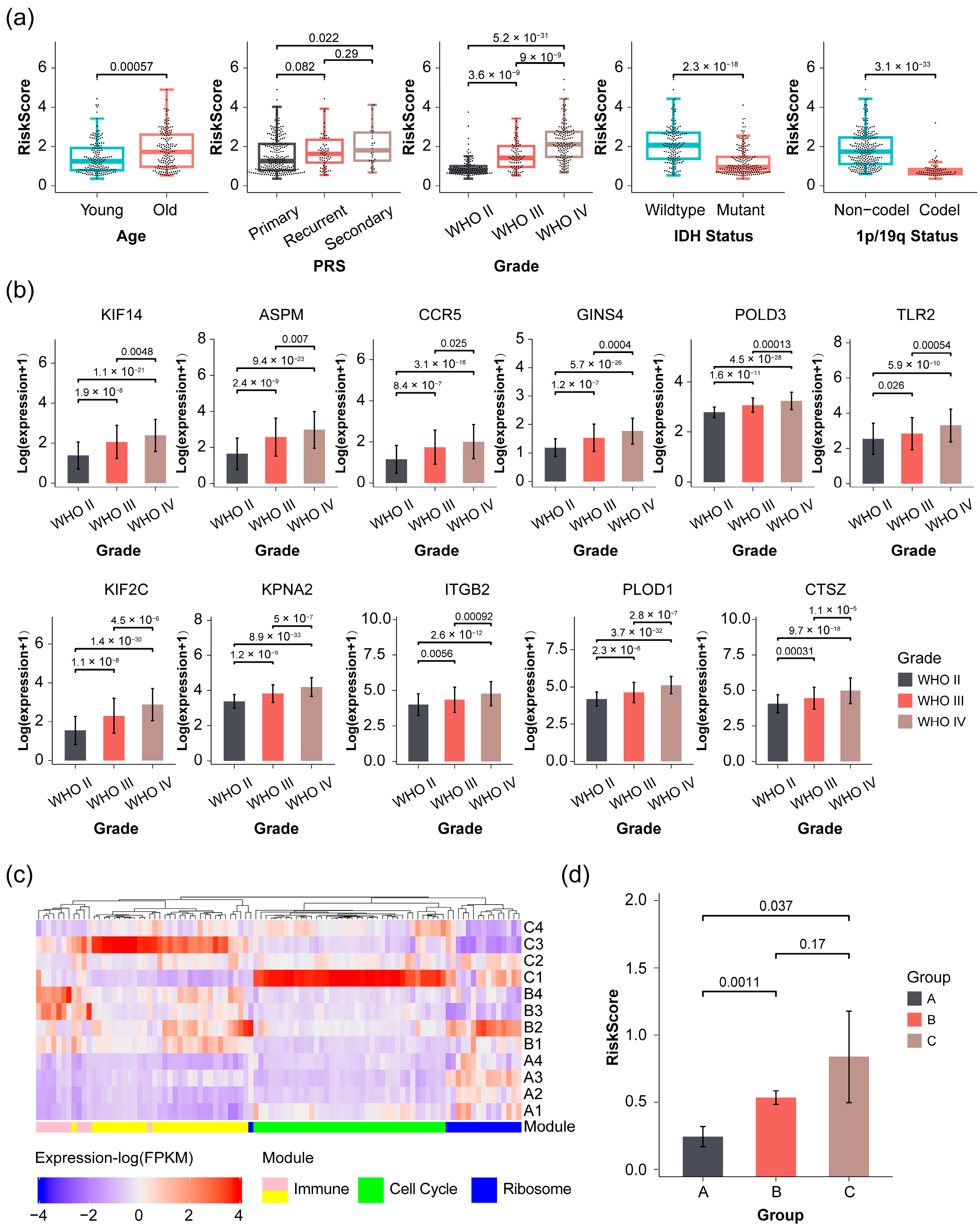
2.4. Validation of Prognostic Signature in Glioma Patients with Varying Severity
2.5. Correlation of Risk Score with Immunological Function Analysis in Glioma Patients
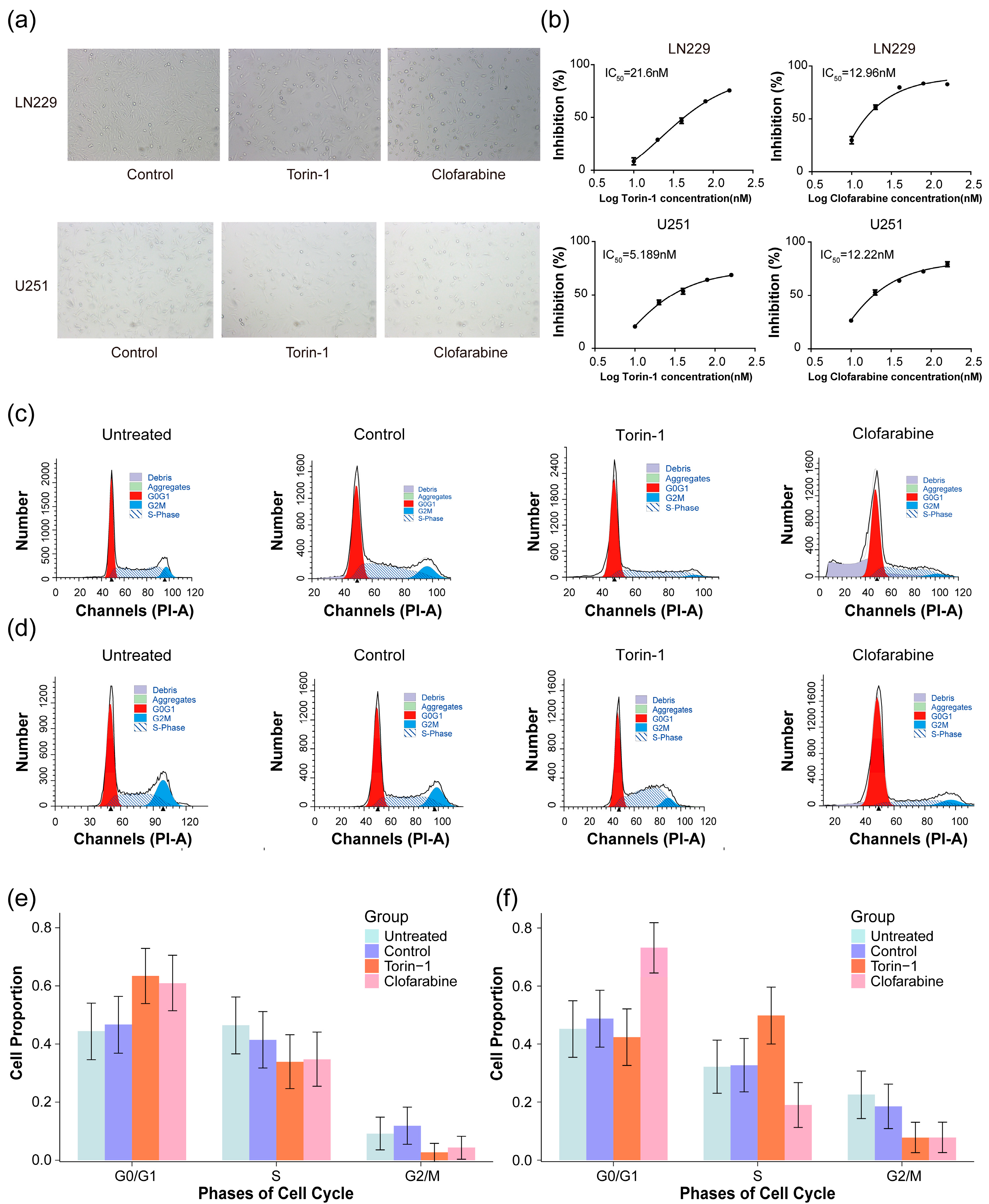
2.6. Screening for Potentially Effective Molecules Targeting Prognostic Genes of Glioma
3. Discussion
4. Materials and Methods
4.1. Data Acquisition and Reprocessing
4.2. Collection of Genes Associated with Glioma
4.3. Weighted Correlation Network Analysis (WGCNA) and Identification of Modules
4.4. Gene Function Enrichment Analysis
4.5. Construction of Glioma-Related Prognostic Signature
4.6. Protein-Protein Interaction (PPI) Analysis
4.7. Prognostic Model Based on Clinical Traits and Risk Scores
4.8. Glioma Tissues and RNA Sequencing
4.9. Immunohistochemical Staining
4.10. Immune Cell Infiltration Analysis
4.11. Estimation of Stromal and Immune Cells
4.12. Drug Screening Based on Prognostic Genes
4.13. Cell Culture and Drug Perturbation
4.14. Experiment and Analysis of Cell Cycle
4.15. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| WGCNA | Weighted Correlation Network Analysis |
| CGGA | Chinese Glioma Genome Atlas |
| TCGA | The Cancer Genome Atlas |
| LASSO | Least Absolute Shrinkage and Selection Operator |
| OS | Overall Survival |
| LINCS | The Library of Integrated Network-Based Cellular Signatures |
| GBM | Glioblastoma Multiforme |
| WHO | World Health Organization |
| LGG | Lower-grade Gliomas |
| HGG | High-grade Gliomas |
| MGMT | O6-methylguanine-DNA Methyltransferase |
| PTEN | Phosphatase and Tensin Homolog |
| TP53 | Tumor Protein p53 |
| EGFR | Epidermal Growth Factor Receptor |
| MET | MET Tyrosine Kinase |
| IDH | Isocitrate Dehydrogenase (gene) |
| H3K27M | Lysine 27-to-Methionine Mutation in Histone H3 |
| KPS | Karnofsky Performance Status |
| TMM | Trimmed Mean of M-values |
| tSNE | t-Distributed Stochastic Neighbor Embedding |
| UMAP | Uniform Manifold Approximation and Projection |
| PRS | Primary-Recurrent-Secondary |
| MM | Module Membership |
| GS | Gene Significance |
| MCODE | Molecular Complex Detection |
| GO | Gene Ontology |
| BP | Biological Process |
| HR | Hazard Ratio |
| ROC | Receiver Operating Characteristic Curve |
| GSEA | Gene Set Enrichment Analysis |
| ssGSEA | Single Sample Gene Set Enrichment Analysis |
| IC50 | Half-maximal Inhibitory Concentration |
| ICB | Immune Checkpoint Blockade |
| FPKM | Fragments per Kilobase per Million Mapped Fragments |
| TOM | Topological Overlap Matrix |
| MEs | Module Eigengenes |
| AUC | Area Under Curve |
| NES | Normalized Enrichment Score |
| TMZ | Temozolomide |
| FDR | False Discovery Rate |
References
- Ostrom, Q.T.; Cioffi, G.; Gittleman, H.; Patil, N.; Waite, K.; Kruchko, C.; Barnholtz-Sloan, J.S. CBTRUS Statistical Report: Primary Brain and Other Central Nervous System Tumors Diagnosed in the United States in 2012–2016. Neuro-Oncology 2019, 21 (Suppl. S5), v1–v100. [Google Scholar] [CrossRef] [PubMed]
- Lin, D.; Wang, M.; Chen, Y.; Gong, J.; Chen, L.; Shi, X.; Lan, F.; Chen, Z.; Xiong, T.; Sun, H.; et al. Trends in Intracranial Glioma Incidence and Mortality in the United States, 1975–2018. Front. Oncol. 2021, 11, 748061. [Google Scholar] [CrossRef] [PubMed]
- Reifenberger, G.; Wirsching, H.G.; Knobbe-Thomsen, C.B.; Weller, M. Advances in the molecular genetics of gliomas—Implications for classification and therapy. Nat. Rev. Clin. Oncol. 2017, 14, 434–452. [Google Scholar] [CrossRef] [PubMed]
- Louis, D.N.; Ohgaki, H.; Wiestler, O.D.; Cavenee, W.K.; Burger, P.C.; Jouvet, A.; Scheithauer, B.W.; Kleihues, P. The 2007 WHO classification of tumours of the central nervous system. Acta Neuropathol. 2007, 114, 97–109. [Google Scholar] [CrossRef] [PubMed]
- Louis, D.N.; Perry, A.; Reifenberger, G.; von Deimling, A.; Figarella-Branger, D.; Cavenee, W.K.; Ohgaki, H.; Wiestler, O.D.; Kleihues, P.; Ellison, D.W. The 2016 World Health Organization Classification of Tumors of the Central Nervous System: A summary. Acta Neuropathol. 2016, 131, 803–820. [Google Scholar] [CrossRef]
- Brat, D.J.; Aldape, K.; Colman, H.; Holland, E.C.; Louis, D.N.; Jenkins, R.B.; Kleinschmidt-DeMasters, B.K.; Perry, A.; Reifenberger, G.; Stupp, R.; et al. cIMPACT-NOW update 3: Recommended diagnostic criteria for “Diffuse astrocytic glioma, IDH-wildtype, with molecular features of glioblastoma, WHO grade IV”. Acta Neuropathol. 2018, 136, 805–810. [Google Scholar] [CrossRef]
- Hu, H.; Mu, Q.; Bao, Z.; Chen, Y.; Liu, Y.; Chen, J.; Wang, K.; Wang, Z.; Nam, Y.; Jiang, B.; et al. Mutational Landscape of Secondary Glioblastoma Guides MET-Targeted Trial in Brain Tumor. Cell 2018, 175, 1665–1678.e18. [Google Scholar] [CrossRef]
- Yang, K.; Liu, Y.; Liu, Z.; Liu, J.; Liu, X.; Chen, X.; Li, C.; Zeng, Y. p38gamma overexpression in gliomas and its role in proliferation and apoptosis. Sci. Rep. 2013, 3, 2089. [Google Scholar] [CrossRef]
- Huang, J.; Liu, F.; Liu, Z.; Tang, H.; Wu, H.; Gong, Q.; Chen, J. Immune Checkpoint in Glioblastoma: Promising and Challenging. Front. Pharmacol. 2017, 8, 242. [Google Scholar] [CrossRef]
- Chen, X.; Zhang, L.; Zhang, I.Y.; Liang, J.; Wang, H.; Ouyang, M.; Wu, S.; da Fonseca, A.C.C.; Weng, L.; Yamamoto, Y.; et al. RAGE expression in tumor-associated macrophages promotes angiogenesis in glioma. Cancer Res. 2014, 74, 7285–7297. [Google Scholar] [CrossRef]
- Liang, X.; Wang, Z.; Dai, Z.; Zhang, H.; Cheng, Q.; Liu, Z. Promoting Prognostic Model Application: A Review Based on Gliomas. J. Oncol. 2021, 2021, 7840007. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Zhang, K.N.; Wang, Q.; Li, G.; Zeng, F.; Zhang, Y.; Wu, F.; Chai, R.; Wang, Z.; Zhang, C.; et al. Chinese Glioma Genome Atlas (CGGA): A Comprehensive Resource with Functional Genomic Data from Chinese Glioma Patients. Genom. Proteom. Bioinform. 2021, 19, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Ma, W.; Fan, W.; Ren, C.; Xu, J.; Zeng, F.; Bao, Z.; Jiang, T.; Zhao, Z. Comprehensive transcriptomic characterization reveals core genes and module associated with immunological changes via 1619 samples of brain glioma. Cell Death Dis 2021, 12, 1140. [Google Scholar] [CrossRef] [PubMed]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.; Qiu, X.; Sun, P.; Ye, Y.; Huang, Q.; Kong, L.; Lu, J.J. Association of IDH mutation and 1p19q co-deletion with tumor immune microenvironment in lower-grade glioma. Mol. Ther. Oncolytics 2021, 21, 288–302. [Google Scholar] [CrossRef]
- Pagès, F.; Galon, J.; Dieu-Nosjean, M.C.; Tartour, E.; Sautès-Fridman, C.; Fridman, W.H. Immune infiltration in human tumors: A prognostic factor that should not be ignored. Oncogene 2010, 29, 1093–1102. [Google Scholar] [CrossRef]
- Fridman, W.H.; Galon, J.; Dieu-Nosjean, M.C.; Cremer, I.; Fisson, S.; Damotte, D.; Pagès, F.; Tartour, E.; Sautès-Fridman, C. Immune infiltration in human cancer: Prognostic significance and disease control. In Cancer Immunology and Immunotherapy; Current Topics in Microbiology and Immunology; Springer: Berlin/Heidelberg, Germany, 2011; Volume 344, pp. 1–24. [Google Scholar] [CrossRef]
- Gordon, S.R.; Maute, R.L.; Dulken, B.W.; Hutter, G.; George, B.M.; McCracken, M.N.; Gupta, R.; Tsai, J.M.; Sinha, R.; Corey, D.; et al. PD-1 expression by tumour-associated macrophages inhibits phagocytosis and tumour immunity. Nature 2017, 545, 495–499. [Google Scholar] [CrossRef]
- Leuzzi, G.; Vasciaveo, A.; Taglialatela, A.; Chen, X.; Firestone, T.M.; Hickman, A.R.; Mao, W.; Thakar, T.; Vaitsiankova, A.; Huang, J.W.; et al. SMARCAL1 is a dual regulator of innate immune signaling and PD-L1 expression that promotes tumor immune evasion. Cell 2024, 187, 861–881.e32. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, Y.; Yang, Y.; Weng, L.; Wu, Q.; Zhang, J.; Zhao, P.; Fang, L.; Shi, Y.; Wang, P. Emerging phagocytosis checkpoints in cancer immunotherapy. Signal. Transduct. Target. Ther. 2023, 8, 104. [Google Scholar] [CrossRef]
- Chen, B.; Ma, L.; Paik, H.; Sirota, M.; Wei, W.; Chua, M.-S.; So, S.; Butte, A.J. Reversal of cancer gene expression correlates with drug efficacy and reveals therapeutic targets. Nat. Commun. 2017, 8, 16022. [Google Scholar] [CrossRef]
- Wang, Q.; Wang, L.; Li, D.; Deng, J.; Zhao, Z.; He, S.; Zhang, Y.; Tu, Y. Kinesin family member 14 is a candidate prognostic marker for outcome of glioma patients. Cancer Epidemiol. 2013, 37, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Li, K.K.; Qi, Y.; Xia, T.; Chan, A.K.; Zhang, Z.Y.; Aibaidula, A.; Zhang, R.; Zhou, L.; Yao, Y.; Ng, H.K. The kinesin KIF14 is overexpressed in medulloblastoma and downregulation of KIF14 suppressed tumor proliferation and induced apoptosis. Lab. Investig. 2017, 97, 946–961. [Google Scholar] [CrossRef] [PubMed]
- Iegiani, G.; Di Cunto, F.; Pallavicini, G. Inhibiting microcephaly genes as alternative to microtubule targeting agents to treat brain tumors. Cell Death Dis. 2021, 12, 956. [Google Scholar] [CrossRef]
- Tu, B.; Xiang, H.; Li, M.; Zhong, F.; Fang, M.; Yan, W. In Silico Analysis of the Correlation of KIF2C with Prognosis and Immune Infiltration in Glioma. Comput. Math. Methods Med. 2022, 2022, 6320828. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Wu, X.; Wang, P.; Cao, S.L.; Peng, B.; Xu, X. ASPM promotes homologous recombination-mediated DNA repair by safeguarding BRCA1 stability. iScience 2021, 24, 102534. [Google Scholar] [CrossRef] [PubMed]
- Bikeye, S.N.; Colin, C.; Marie, Y.; Vampouille, R.; Ravassard, P.; Rousseau, A.; Boisselier, B.; Idbaih, A.; Calvo, C.F.; Leuraud, P.; et al. ASPM-associated stem cell proliferation is involved in malignant progression of gliomas and constitutes an attractive therapeutic target. Cancer Cell Int. 2010, 10, 1. [Google Scholar] [CrossRef]
- Kranjc, M.K.; Novak, M.; Pestell, R.G.; Lah, T.T. Cytokine CCL5 and receptor CCR5 axis in glioblastoma multiforme. Radiol. Oncol. 2019, 53, 397–406. [Google Scholar] [CrossRef]
- Shan, D.D.; Zheng, Q.X.; Chen, Z. Go-Ichi-Ni-San 2: A potential biomarker and therapeutic target in human cancers. World J. Gastrointest. Oncol. 2022, 14, 1892–1902. [Google Scholar] [CrossRef]
- Bellido, F.; Pineda, M.; Aiza, G.; Valdés-Mas, R.; Navarro, M.; Puente, D.A.; Pons, T.; González, S.; Iglesias, S.; Darder, E.; et al. POLE and POLD1 mutations in 529 kindred with familial colorectal cancer and/or polyposis: Review of reported cases and recommendations for genetic testing and surveillance. Genet. Med. 2016, 18, 325–332. [Google Scholar] [CrossRef]
- Sanefuji, K.; Taketomi, A.; Iguchi, T.; Sugimachi, K.; Ikegami, T.; Yamashita, Y.; Gion, T.; Soejima, Y.; Shirabe, K.; Maehara, Y. Significance of DNA polymerase delta catalytic subunit p125 induced by mutant p53 in the invasive potential of human hepatocellular carcinoma. Oncology 2010, 79, 229–237. [Google Scholar] [CrossRef]
- Yang, C.A.; Huang, H.Y.; Lin, C.L.; Chang, J.G. G6PD as a predictive marker for glioma risk, prognosis and chemosensitivity. J. Neurooncol. 2018, 139, 661–670. [Google Scholar] [CrossRef] [PubMed]
- Bae, J.M.; Yoo, S.Y.; Kim, J.H.; Kang, G.H. Immune landscape and biomarkers for immuno-oncology in colorectal cancers. J. Pathol. Transl. Med. 2020, 54, 351–360. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Luo, Y.B.; Wu, W.; Zhang, L.; Wang, Z.; Dai, Z.; Feng, S.; Cao, H.; Cheng, Q.; Liu, Z. The molecular feature of macrophages in tumor immune microenvironment of glioma patients. Comput. Struct. Biotechnol. J. 2021, 19, 4603–4618. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Li, J.; Ma, J.; Bao, Y. ZNF143 facilitates the growth and migration of glioma cells by regulating KPNA2-mediated Hippo signalling. Sci. Rep. 2023, 13, 11097. [Google Scholar] [CrossRef]
- Zhang, Z.; Huang, X.; Li, J.; Fan, H.; Yang, F.; Zhang, R.; Yang, Y.; Feng, S.; He, D.; Sun, W.; et al. Interleukin 10 promotes growth and invasion of glioma cells by up-regulating KPNA 2 in vitro. J. Cancer Res. Ther. 2019, 15, 927–932. [Google Scholar] [CrossRef]
- Sai Krishna, A.V.S.; Ramu, A.; Hariharan, S.; Sinha, S.; Donakonda, S. Characterization of tumor microenvironment in glioblastoma multiforme identifies ITGB2 as a key immune and stromal related regulator in glial cell types. Comput. Biol. Med. 2023, 165, 107433. [Google Scholar] [CrossRef]
- Seton-Rogers, S. Microenvironment: Source influences function. Nat. Rev. Cancer 2014, 14, 704–705. [Google Scholar] [CrossRef]
- Guo, X.; Wang, G. Advances in research on immune escape mechanism of glioma. CNS Neurosci. Ther. 2023, 29, 1709–1720. [Google Scholar] [CrossRef]
- Xu, S.; Tang, L.; Li, X.; Fan, F.; Liu, Z. Immunotherapy for glioma: Current management and future application. Cancer Lett. 2020, 476, 1–12. [Google Scholar] [CrossRef]
- Sathornsumetee, S.; Reardon, D.A.; Desjardins, A.; Quinn, J.A.; Vredenburgh, J.J.; Rich, J.N. Molecularly targeted therapy for malignant glioma. Cancer 2007, 110, 13–24. [Google Scholar] [CrossRef]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [PubMed]
- Piñero, J.; Bravo, À.; Queralt-Rosinach, N.; Gutiérrez-Sacristán, A.; Deu-Pons, J.; Centeno, E.; García-García, J.; Sanz, F.; Furlong, L.I. DisGeNET: A comprehensive platform integrating information on human disease-associated genes and variants. Nucleic Acids Res. 2017, 45, D833–D839. [Google Scholar] [CrossRef] [PubMed]
- Langfelder, P.; Horvath, S. WGCNA: An R package for weighted correlation network analysis. BMC Bioinform. 2008, 9, 559. [Google Scholar] [CrossRef]
- Sherman, B.T.; Hao, M.; Qiu, J.; Jiao, X.; Baseler, M.W.; Lane, H.C.; Imamichi, T.; Chang, W. DAVID: A web server for functional enrichment analysis and functional annotation of gene lists (2021 update). Nucleic Acids Res. 2022, 50, W216–W221. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [PubMed]
- Mootha, V.K.; Lindgren, C.M.; Eriksson, K.-F.; Subramanian, A.; Sihag, S.; Lehar, J.; Puigserver, P.; Carlsson, E.; Ridderstråle, M.; Laurila, E.; et al. PGC-1α-responsive genes involved in oxidative phosphorylation are coordinately downregulated in human diabetes. Nat. Genet. 2003, 34, 267–273. [Google Scholar] [CrossRef]
- Tibshirani, R. The lasso method for variable selection in the Cox model. Stat. Med. 1997, 16, 385–395. [Google Scholar] [CrossRef]
- Hänzelmann, S.; Castelo, R.; Guinney, J. GSVA: Gene set variation analysis for microarray and RNA-seq data. BMC Bioinform. 2013, 14, 7. [Google Scholar] [CrossRef]
- Zhang, C.-B.; Cheng, W.; Ren, X.; Liu, X.; Li, G.; Han, S.; Jiang, T.; Wu, A. Tumor Purity As an Underlying Key Factor in Glioma. Clin. Cancer Res. 2017, 23, 6279–6291. [Google Scholar] [CrossRef]
- Yoshihara, K.; Shahmoradgoli, M.; Martínez, E.; Vegesna, R.; Kim, H.; Torres-Garcia, W.; Treviño, V.; Shen, H.; Laird, P.W.; Levine, D.A.; et al. Inferring tumour purity and stromal and immune cell admixture from expression data. Nat. Commun. 2013, 4, 2612. [Google Scholar] [CrossRef]
- Subramanian, A.; Narayan, R.; Corsello, S.M.; Peck, D.D.; Natoli, T.E.; Lu, X.; Gould, J.; Davis, J.F.; Tubelli, A.A.; Asiedu, J.K.; et al. A Next Generation Connectivity Map: L1000 Platform and the First 1,000,000 Profiles. Cell 2017, 171, 1437–1452.e17. [Google Scholar] [CrossRef] [PubMed]
- JASP Team. JASP (Version 0.19.0) [Computer Software]. 2024. Available online: https://jasp-stats.org/ (accessed on 15 July 2024).




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| LN229 Cells | U251 Cells | |||
|---|---|---|---|---|
| Control and Torin-1 | Control and Clofarabine | Control and Torin-1 | Control and Clofarabine | |
| Chi-square statistic | 9.110 | 6.042 | 8.342 | 12.849 |
| Degree of freedom | 2 | 2 | 2 | 2 |
| p-value | 0.011 | 0.049 | 0.015 | 0.002 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, K.; Yang, J.; Jiang, R.; Ren, X.; Liu, P.; Wang, W.; Zhou, S.; Wang, X.; Ma, L.; Hu, Y. Identification of Key Immune and Cell Cycle Modules and Prognostic Genes for Glioma Patients through Transcriptome Analysis. Pharmaceuticals 2024, 17, 1295. https://doi.org/10.3390/ph17101295
Guo K, Yang J, Jiang R, Ren X, Liu P, Wang W, Zhou S, Wang X, Ma L, Hu Y. Identification of Key Immune and Cell Cycle Modules and Prognostic Genes for Glioma Patients through Transcriptome Analysis. Pharmaceuticals. 2024; 17(10):1295. https://doi.org/10.3390/ph17101295
Chicago/Turabian StyleGuo, Kaimin, Jinna Yang, Ruonan Jiang, Xiaxia Ren, Peng Liu, Wenjia Wang, Shuiping Zhou, Xiaoguang Wang, Li Ma, and Yunhui Hu. 2024. "Identification of Key Immune and Cell Cycle Modules and Prognostic Genes for Glioma Patients through Transcriptome Analysis" Pharmaceuticals 17, no. 10: 1295. https://doi.org/10.3390/ph17101295