
Salt Effect on the Antioxidant Activity of Red Microalgal Sulfated Polysaccharides in Soy-Bean Formula



{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
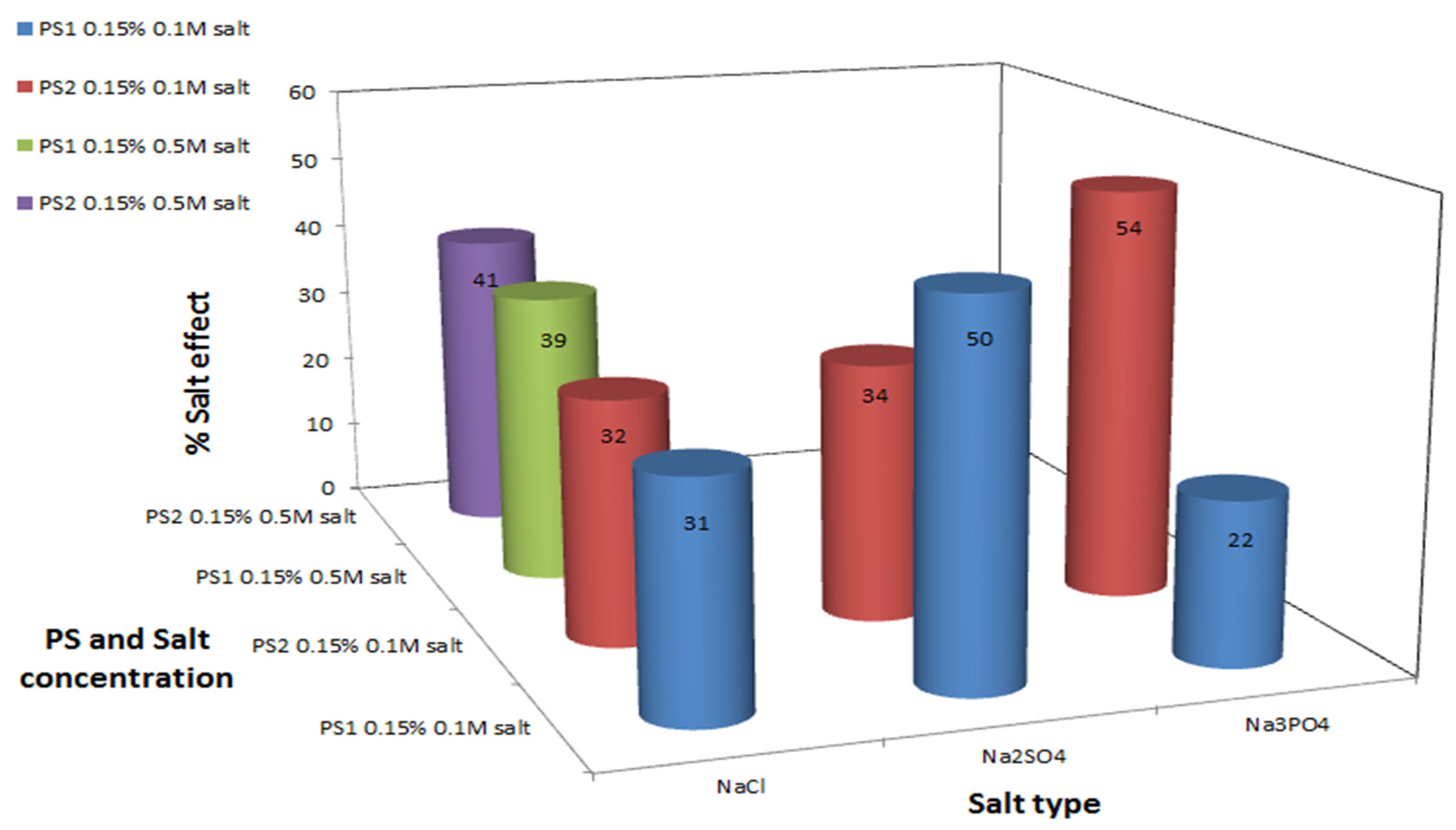
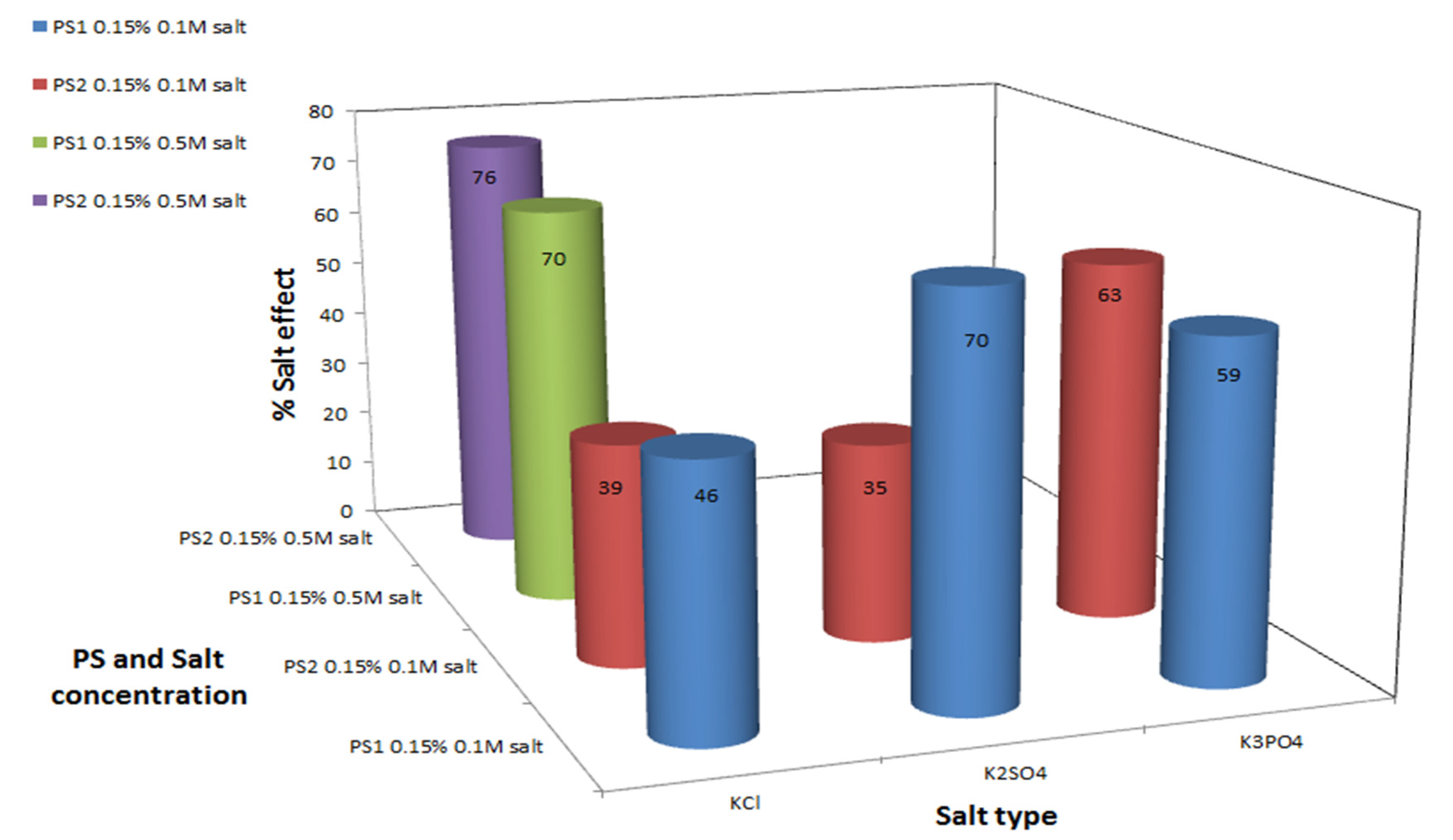
2. Results and Discussion




3. Experimental Section
3.1. Materials
3.2. Algae and Growth Conditions
3.3. Isolation and Quantitative Analysis of Soluble-Sulfated Polysaccharide Fraction
3.4. Sample Preparation
3.5. Production of Free Radicals
3.6. Malondialdehyde (MDA) Measurements
3.7. Calculation of % Salt Effect
3.8. Statistical Analyses
4. Conclusions
Supplementary Files
Supplementary File 1Author Contributions
Conflicts of Interest
References
- Angele-Martinez, C.; Goodman, C.; Brumaghim, J. Metal-mediated DNA damage and cell death: Mechanisms, detection methods, and cellular consequences. Metallomics 2014, 6, 1358–1381. [Google Scholar] [CrossRef] [PubMed]
- Bregano, J.W.; Dichi, I. Inflammatory bowel disease. In Role of Oxidative Stress Chronic Diseases; Dichi, I., Breganó, J.W, Simão, A.N.C., Cecchini, R., Eds.; CRC Press, Taylor & Francis Group: Boca Raton, FL, USA, 2014; pp. 501–520. [Google Scholar]
- Furlan, A.L.; Jobin, M.-L.; Buchoux, S.; Grelard, A.; Dufourc, E.J.; Gean, J. Membrane lipids protected from oxidation by red wine tannins: A proton NMR study. Biochimie 2014, 107, 82–90. [Google Scholar] [CrossRef] [PubMed]
- Harrison, I.P.; Selemidis, S. Understanding the biology of reactive oxygen species and their link to cancer: NADPH oxidases as novel pharmacological targets. Clin. Exp. Pharmacol. Physiol. 2014, 41, 533–542. [Google Scholar] [CrossRef] [PubMed]
- Butterfield, D.A. Amyloid β-peptide (1-42)-induced oxidative stress and neurotoxicity: Implications for neurodegeneration in Alzheimer’s disease brain. Free Radic. Res. 2002, 36, 1307–1313. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B.; Aruoma, O.I. DNA damage by oxygen-derived species. Its mechanism and measurement in mammalian systems. FEBS Lett. 1991, 281, 9–19. [Google Scholar] [CrossRef]
- Froelich, L.; Riederer, P. Free radical mechanisms in dementia of Alzheimer type and the potential for antioxidative treatment. Arzneimittelforschung 1995, 45, 443–446. [Google Scholar]
- Yang, C.S.; Landau, J.M.; Huang, M.-T.; Newmark, H.L. Inhibition of carcinogenesis by dietary polyphenolic compounds. Annu. Rev. Nutr. 2001, 21, 381–406. [Google Scholar] [CrossRef] [PubMed]
- Ramsaha, S.; Aumjaud, B.E.; Neergheen-Bhujun, V.S.; Bahorun, T. Polyphenolic rich traditional plants and teas improve lipid stability in food test systems. J. Food Sci. Technol. 2015, 52, 773–782. [Google Scholar] [CrossRef] [PubMed]
- Park, P.-J.; Jung, W.-K.; Nam, K.-S.; Shahidi, F.; Kim, S.-K. Purification and characterization of antioxidative peptides from protein hydrolyzate of lecithin-free egg yolk. J. Am. Oil Chem. Soc. 2001, 78, 651–656. [Google Scholar] [CrossRef]
- Kazi, A.; Deshmukh, S.; Mirza, R.; Murtadak, S. A comprehensive review on flavonoids. World J. Pharm. Res. 2015, 4, 560–572. [Google Scholar]
- Mao, G.; Zhang, S.; Song, H.; Ding, S.; Zhu, P.; Wang, X.; Liang, C. Synthesis, biological activities and therapeutic properties of esculetin and its derivatives. J. Chem. Pharm. Res. 2015, 7, 122–130. [Google Scholar]
- Pena-Ramos, E.A.; Xiong, Y.L. Antioxidative activity of whey protein hydrolysates in a liposomal system. J. Dairy Sci. 2001, 84, 2577–2583. [Google Scholar] [CrossRef]
- Sharma, R.; Rani, D.; Jain, N.; Kantwa, S.M.; Jaitawat, A. Cinnamon—A natural replacement for synthetic drugs: A review. Int. J. Curr. Res. Biosci. Plant Biol. 2015, 2, 69–78. [Google Scholar]
- Jiao, G.; Yu, G.; Zhang, J.; Ewart, H.S. Chemical structures and bioactivities of sulfated polysaccharides from marine algae. Mar. Drugs 2011, 9, 196–223. [Google Scholar] [CrossRef] [PubMed]
- Ruperez, P.; Ahrazem, O.; Leal, J.A. Potential Antioxidant Capacity of Sulfated Polysaccharides from the Edible Marine Brown Seaweed Fucus vesiculosus. J. Agric. Food Chem. 2002, 50, 840–845. [Google Scholar] [CrossRef] [PubMed]
- Rocha de Souza, M.C.; Marques, C.T.; Guerra Dore, C.M.; Ferreira da Silva, F.R.; Oliveira Rocha, H.A.; Leite, E.L. Antioxidant activities of sulfated polysaccharides from brown and red seaweeds. J. Appl. Phycol. 2007, 19, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhang, Q.; Zhang, Z.; Song, H.; Li, P. Potential antioxidant and anticoagulant capacity of low molecular weight fucoidan fractions extracted from Laminaria japonica. Int. J. Biol. Macromol. 2010, 46, 6–12. [Google Scholar] [CrossRef] [PubMed]
- Dore, C.M.; das C Faustino Alves, M.G.; Will, L.S.; Costa, T.G.; Sabry, D.A.; de Souza Rêgo, L.A.; Accardo, C.M.; Rocha, H.A.; Filgueira, L.G.; Leite, E.L. A sulfated polysaccharide, fucans, isolated from brown algae Sargassum vulgare with anticoagulant, antithrombotic, antioxidant and anti-inflammatory effects. Carbohydr. Polym. 2013, 91, 467–475. [Google Scholar] [CrossRef] [PubMed]
- Qi, H.; Zhang, Q.; Zhao, T.; Hu, R.; Zhang, K.; Li, Z. In vitro antioxidant activity of acetylated and benzoylated derivatives of polysaccharide extracted from Ulva pertusa (Chlorophyta). Bioorg. Med. Chem. Lett. 2006, 19, 2441–2445. [Google Scholar] [CrossRef] [PubMed]
- Xue, Z.; Xue, C.; Cai, Y.; Wang, D.; Fang, Y. The study of antioxidant activities of fucoidan from Laminaria japonica. High Technol. Lett. 2005, 11, 91–94. [Google Scholar]
- Costa, L.S.; Fidelis, G.P.; Cordeiro, S.L.; Oliveira, R.M.; Sabry, D.A.; Camara, R.B.; Nobre, L.T.; Costa, M.S.; Almeida-Lima, J.; Farias, E.H.; et al. Biological activities of sulfated polysaccharides from tropical seaweeds. Biomed. Pharmacother. 2010, 64, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Wang, C.; Shi, Q.; Ma, C. Preparation of different molecular weight polysaccharides from Porphyridium cruentum and their antioxidant activities. Int. J. Biol. Macromol. 2009, 45, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Tannin-Spitz, T.; Bergman, M.; van-Moppes, D.; Grossman, S.; Arad, S. Antioxidant activity of the polysaccharide of the red microalga Porphyridium sp. J. Appl. Phycol. 2005, 17, 215–222. [Google Scholar] [CrossRef]
- Chen, B.; You, W.; Huang, J.; Yu, Y.; Chen, W. Isolation and antioxidant property of the extracellular polysaccharide from Rhodella reticulata. World J. Microbiol. Biotechnol. 2010, 26, 833–840. [Google Scholar] [CrossRef]
- Wang, J.; Hu, S.; Nie, S.; Yu, Q.; Xie, M. Reviews on Mechanisms of in Vitro Antioxidant Activity of Polysaccharides. Oxid. Medi. Cell. Longev. 2015; in press. [Google Scholar]
- Bold, H.C.; Wynne, M.J. Introduction to the Algae: Structure and Reproduction, 2nd ed.; Prentice Hall Inc.: Englewood Cliiffs, NJ, USA, 1985. [Google Scholar]
- Ramus, J. Rhodophyte unicells, biopolymer, physiology and production. In Algal Biomass Technologies—An Interdisciplinary Perspective; Barclay, W.R., McIntosh, R.P., Eds.; Cramer: Berlin, Germany, 1986; pp. 51–55. [Google Scholar]
- Arad, M.S. Production of sulfated polysaccharides from red unicellular algae. In Algal Biotechnology—An Interdisciplinary Perspective; Stadler, T., Mollion, J., Verduset, M.C., Eds.; Elsevier Applied Science: London, UK, 1988; pp. 65–87. [Google Scholar]
- Arad, S.; Levy-Ontman, O. Red microalgal cell wall polysaccharides: Biotechnological aspects. Curr. Opin. Biotechnol. 2010, 21, 358–364. [Google Scholar] [CrossRef] [PubMed]
- Ucko, M.; Cohen, E.; Gordin, H.; Arad, S. Relationship between the unicellular red alga Porphyridium sp. and its predator, the dinoflagellate Gymnodinium sp. Appl. Environ. Microbiol. 1989, 55, 2990–2994. [Google Scholar] [PubMed]
- Arad, S.; Keristovesky, G.; Simon, B.; Barak, Z.E.; Geresh, S. Biodegradation of the sulfated polysaccharide of Porphyridium by soil bacteria. Phytochemistry 1993, 32, 287–290. [Google Scholar] [CrossRef]
- Percival, E.; Foyle, R.A.J. The extracellular polysaccharides of Porphyridium cruentum and Porphyridium aerugineum. Carbohydr. Res. 1979, 72, 165–176. [Google Scholar] [CrossRef]
- Geresh, S.; Arad, S. The extracellular polysaccharides of the red microalgae: Chemistry and rheology. Bioresour. Technol. 1991, 38, 195–201. [Google Scholar] [CrossRef]
- Heaney-Kieras, J.; Chapman, D.J. Structural studies on the extracellular polysaccharide of the red alga, Porphyridium cruentum. Carbohydr. Res. 1976, 52, 169–177. [Google Scholar] [CrossRef]
- Geresh, S.; Adin, I.; Yarmolinsky, E.; Karpasas, M. Characterization of the extracellular polysaccharide of Porphyridium sp.: Molecular weight determination and rheological properties. Carbohydr. Polym. 2002, 50, 183–189. [Google Scholar] [CrossRef]
- Geresh, S.; Dubinsky, O.; Arad, M.S.; Christiaen, D.; Glaser, R. Structure of 3-O-(alpha-d-glucopyranosyluronic acid)-l-galactopyranose, an aldobiouronic acid isolated from the polysaccharides of various unicellular red algae. Carbohydr. Res. 1990, 208, 301–305. [Google Scholar] [CrossRef]
- Shrestha, R.P.; Weinstein, Y.; Bar-Zui, D.; Arad, S. A glycoprotein noncovalently associated with cell-wall polysaccharide of the red microalga Porphyridium sp. (Rhodophyta). J. Phycol. 2004, 40, 568–580. [Google Scholar] [CrossRef]
- Levy-Ontman, O.; Arad, S.; Harvey, D.J.; Parsons, T.B.; Fairbanks, A.; Tekoah, Y. Unique N-glycan moieties of the 66-kDa cell wall glycoprotein from the red microalga Porphyridium sp. J. Biol. Chem. 2011, 286, 21340–21352. [Google Scholar] [CrossRef] [PubMed]
- Ucko, M.; Shrestha, R.P.; Mesika, P.; Bar-Zvi, D.; Arad, S. Glycoprotein moiety in the cell wall of the red microalga Porphyridium sp. (Rhodophyta) as the biorecognition site for the Crypthecodinium cohnii-like dinoflagellate. J. Phycol. 1999, 35, 1276–1281. [Google Scholar] [CrossRef]
- Arad, S.; Levy-Ontman, O. Sulfated polysaccharides in the cell wall of red microalgae. In Handbook of Biopolymer-Based Materials: From Blends and Composites to Gels and Complex; Sabu, T., Dominique, D., Christophe, C., Jyotishkumar, P., Eds.; Wiley-VCH Verlag: Berlin, Germany, 2013; Volume 2, pp. 351–370. [Google Scholar]
- Yaron, A.; Cohen, E.; Arad, S.M. Stabilization of aloe vera gel by interaction with sulfated polysaccharides from red microalgae and with xanthan gum. J. Agric. Food Chem. 1992, 40, 1316–1320. [Google Scholar] [CrossRef]
- Eteshola, E.; Gottlieb, M.; Arad, S. Dilute solution viscosity of red microalga exopolysaccharide. Chem. Eng. Sci. 1996, 51, 1487–1494. [Google Scholar] [CrossRef]
- Ginzberg, A.; Korin, E.; Arad, S.M. Effect of drying on the biological activities of a red microalgal polysaccharide. Biotechnol. Bioeng. 2008, 99, 411–420. [Google Scholar] [CrossRef] [PubMed]
- Savins, J.G. Oil Recovery Process Employing Thickened Aqueous Driving Fluid. U.S. Patent 4,079,544, 21 March 1978. [Google Scholar]
- Arad, S.; Weinstein, Y. Novel lubricants from red microalgae: Interplay between genes and products. Biomedic 2003, 1, 32–37. [Google Scholar]
- Arad, M.S.; Richmond, A. Industrial production of microalgal cell-mass and secondary products—Species of high potential: Porphyridium sp. In Handbook of Microalgal Culture: Biotechnology and Applied Phycology; Richmond, A., Ed.; Blackwell Publishing Ltd.: Oxford, UK, 2004; pp. 289–299. [Google Scholar]
- Arad, M.S.; Rapoport, L.; Moshkovich, A.; van-Moppes, D.; Karpasas, M.; Golan, R.; Golan, Y. Superior biolubricant from a species of red microalga. Langmuir 2006, 22, 7313–7317. [Google Scholar] [CrossRef] [PubMed]
- Eteshola, E.; Karpasas, M.; Arad, S.; Gottlieb, M. Red microalga exo polysaccharides. Part 2. Study of the rheology, morphology, and thermal gelation of aqueous preparations. Acta Polym. 1998, 49, 549–556. [Google Scholar] [CrossRef]
- Liu, F.; Ooi, V.E.C.; Chang, S.T. Free radical scavenging activities of mushroom polysaccharide extracts. Life Sci. 1997, 60, 763–771. [Google Scholar] [CrossRef]
- Liu, D.; Sheng, J.; Li, Z.; Qi, H.; Sun, Y.; Duan, Y.; Zhang, W. Antioxidant activity of polysaccharide fractions extracted from Athyrium multidentatum (Doll.) Ching. Int. J. Biol. Macromol. 2013, 56, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Leung, P.H.; Zhao, S.; Ho, K.P.; Wu, J.Y. Chemical properties and antioxidant activity of exopolysaccharides from mycelial culture of Cordyceps sinensis fungus Cs-HK1. Food Chem. 2009, 114, 1251–1256. [Google Scholar] [CrossRef]
- Cheung, Y.-C.; Siu, K.-C.; Liu, Y.-S.; Wu, J.-Y. Molecular properties and antioxidant activities of polysaccharide-protein complexes from selected mushrooms by ultrasound-assisted extraction. Process Biochem. 2012, 47, 892–895. [Google Scholar] [CrossRef]
- Huang, Q.-L.; Siu, K.-C.; Wang, W.-Q.; Cheung, Y.-C.; Wu, J.-Y. Fractionation, characterization and antioxidant activity of exopolysaccharides from fermentation broth of a Cordyceps sinensis fungus. Process Biochem. 2013, 48, 380–386. [Google Scholar] [CrossRef]
- Dumitriu, S.; Vidal, P.F.; Chornet, E. Hydrogels based on polysaccharides. In Polysaccharides in medical application; Dumitriu, S., Ed.; Marcel Dekker, Inc.: New York, NY, USA, 1996; pp. 125–241. [Google Scholar]
- Shao, F.; Ankur, T.; Diana, M.S.; Riccardo, L.B.; Ira, S.B.; Sachin, V.; Eric, J.M.; Lawrence, H.B. Relevance of Rheological Properties of Sodium Alginate in Solution to Calcium Alginate Gel Properties. Res. Artic. 2011, 12, 453–460. [Google Scholar]
- Seely, G.R.; Hart, R.L. Binding of alkaline earth metal ions to alginate. Macromolecules 1974, 7, 706–710. [Google Scholar] [CrossRef] [PubMed]
- Florian-Algarin, V.; Acevedo, A. Rheology and Thermotropic Gelation of Aqueous Sodium Alginate Solutions. J. Pharm. Innov. 2010, 5, 37–44. [Google Scholar] [CrossRef]
- Jones, R.H.; Speer, H.L.; Kury, W. Studies on the growth of the red alga Porphyridium cruentum. Physiol. Plant. 1963, 16, 636–643. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Halliwell, B. Free radicals and antioxidants—Quo vadis? Trends Pharmacol. Sci. 2011, 32, 125–130. [Google Scholar] [CrossRef]
- Fenaille, F.; Mottier, P.; Turesky, R.J.; Ali, S.; Guy, P.A. Comparison of analytical techniques to quantify malondialdehyde in milk powders. J. Chromatogr. A 2001, 921, 237–245. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Burg, A.; Oshrat, L.-O. Salt Effect on the Antioxidant Activity of Red Microalgal Sulfated Polysaccharides in Soy-Bean Formula. Mar. Drugs 2015, 13, 6425-6439. https://doi.org/10.3390/md13106425
Burg A, Oshrat L-O. Salt Effect on the Antioxidant Activity of Red Microalgal Sulfated Polysaccharides in Soy-Bean Formula. Marine Drugs. 2015; 13(10):6425-6439. https://doi.org/10.3390/md13106425
Chicago/Turabian StyleBurg, Ariela, and Levy-Ontman Oshrat. 2015. "Salt Effect on the Antioxidant Activity of Red Microalgal Sulfated Polysaccharides in Soy-Bean Formula" Marine Drugs 13, no. 10: 6425-6439. https://doi.org/10.3390/md13106425
APA StyleBurg, A., & Oshrat, L. -O. (2015). Salt Effect on the Antioxidant Activity of Red Microalgal Sulfated Polysaccharides in Soy-Bean Formula. Marine Drugs, 13(10), 6425-6439. https://doi.org/10.3390/md13106425