
Development of a Dunaliella tertiolecta Strain with Increased Zeaxanthin Content Using Random Mutagenesis

Abstract
:1. Introduction
2. Results
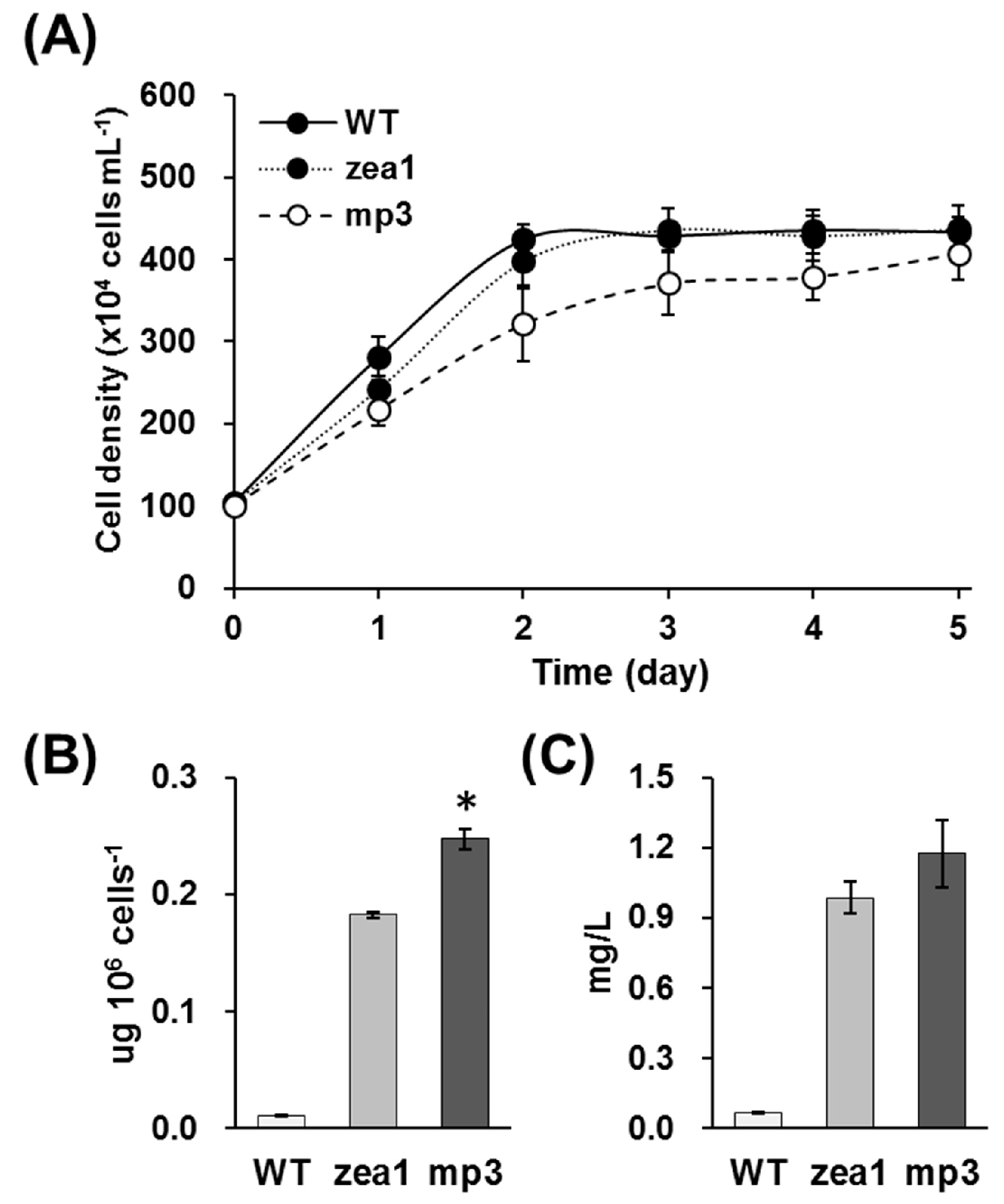
2.1. Selection and Characterization of the Dunaliella Tertiolecta mp3 Strain
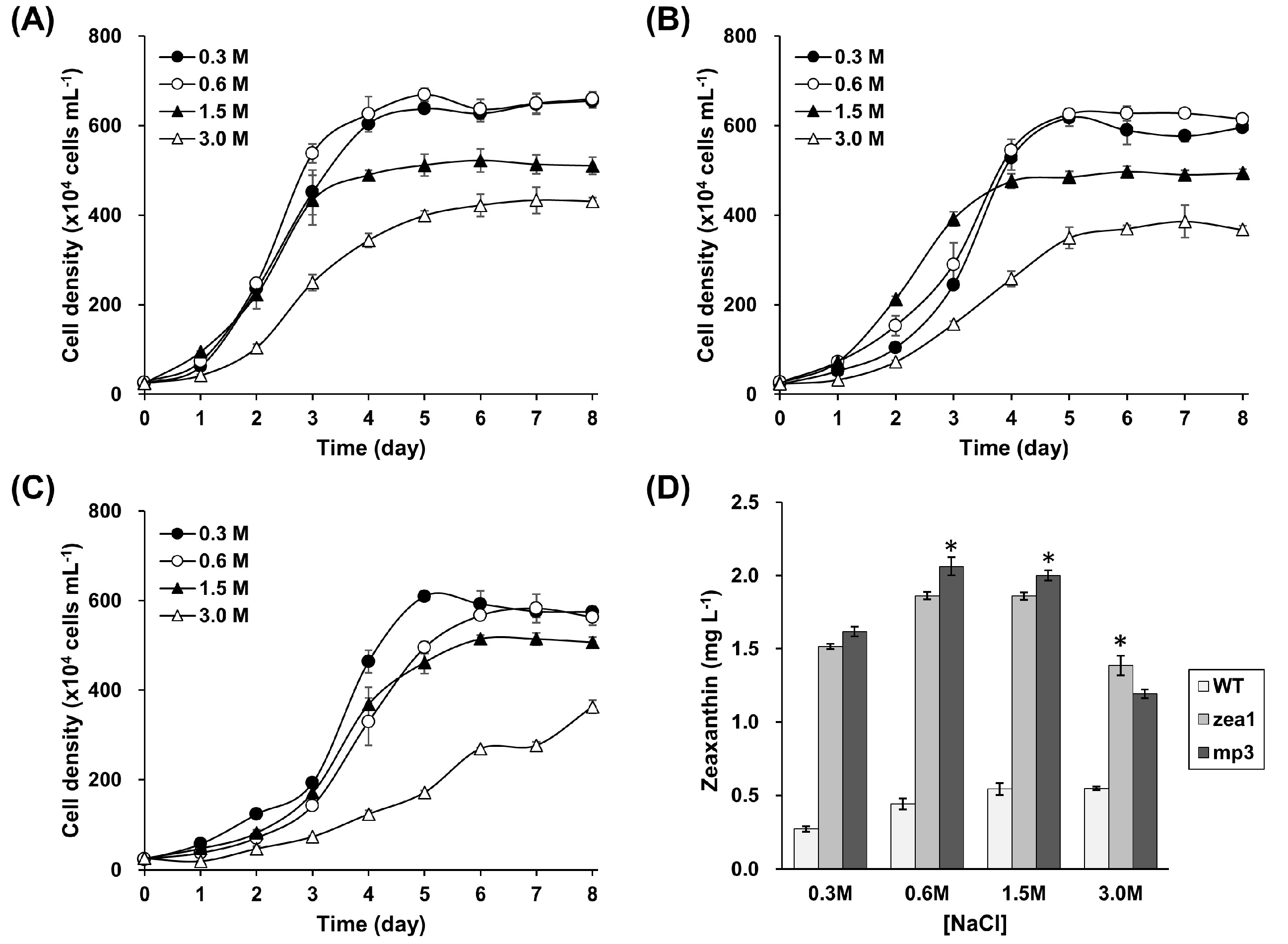
2.2. Effect of NaCl Concentration on Cell Density and Zeaxanthin Yield
2.3. Effect of Light Intensity on Growth Rate and Zeaxanthin Productivity
2.4. Zeaxanthin Yield in Two Different Batch Culture Systems
3. Discussion
4. Materials and Methods
4.1. Strains and Culture Conditions
4.2. Mutagenesis and Mutant Selection
4.3. Experimental Culture Conditions
4.3.1. Culture Medium Optimization: NaCl Concentration
4.3.2. Culture Condition Optimization: Irradiance
4.3.3. Repeated Fed-Batch Culture
4.4. Analytical Methods
4.4.1. Specific Growth Rate and Biomass
4.4.2. Pigment Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Mulders, K.J.; Lamers, P.P.; Martens, D.E.; Wijffels, R.H. Phototrophic pigment production with microalgae: Biological constraints and opportunities. J. Phycol. 2014, 50, 229–242. [Google Scholar] [CrossRef] [PubMed]
- Lemoine, Y.; Schoefs, B. Secondary ketocarotenoid astaxanthin biosynthesis in algae: A multifunctional response to stress. Photosynth. Res. 2010, 106, 155–177. [Google Scholar] [CrossRef] [PubMed]
- Bernstein, P.S.; Li, B.; Vachali, P.P.; Gorusupudi, A.; Shyam, R.; Henriksen, B.S.; Nolan, J.M. Lutein, zeaxanthin, and meso-zeaxanthin: The basic and clinical science underlying carotenoid-based nutritional interventions against ocular disease. Prog. Retin. Eye Res. 2016, 50, 34–66. [Google Scholar] [CrossRef] [PubMed]
- Nwachukwu, I.D.; Udenigwe, C.C.; Aluko, R.E. Lutein and zeaxanthin: Production technology, bioavailability, mechanisms of action, visual function, and health claim status. Trends Food Sci. Technol. 2016, 49, 74–84. [Google Scholar] [CrossRef]
- Lin, J.-H.; Lee, D.-J.; Chang, J.-S. Lutein production from biomass: Marigold flowers versus microalgae. Bioresour. Technol. 2015, 184, 421–428. [Google Scholar] [CrossRef] [PubMed]
- Sajilata, M.; Singhal, R.; Kamat, M. The carotenoid pigment zeaxanthin—A review. Compr. Rev. Food Sci. Food Saf. 2008, 7, 29–49. [Google Scholar] [CrossRef]
- Johnson-Down, L.; Saudny-Unterberger, H.; Gray-Donald, K. Food habits of Canadians: Lutein and lycopene intake in the Canadian population. J. Am. Diet. Assoc. 2002, 102, 988–991. [Google Scholar] [CrossRef]
- Hadden, W.L.; Watkins, R.H.; Levy, L.W.; Regalado, E.; Rivadeneira, D.M.; Van Breemen, R.B.; Schwartz, S.J. Carotenoid composition of marigold (Tagetes erecta) flower extract used as nutritional supplement. J. Agric. Food Chem. 1999, 47, 4189–4194. [Google Scholar] [CrossRef] [PubMed]
- Del Campo, J.; Rodriguez, H.; Moreno, J.; Vargas, M.; Rivas, J.; Guerrero, M. Accumulation of astaxanthin and lutein in Chlorella zofingiensis (Chlorophyta). Appl. Microbiol. Biotechnol. 2004, 64, 848–854. [Google Scholar] [CrossRef] [PubMed]
- Ho, S.-H.; Chan, M.-C.; Liu, C.-C.; Chen, C.-Y.; Lee, W.-L.; Lee, D.-J.; Chang, J.-S. Enhancing lutein productivity of an indigenous microalga Scenedesmus obliquus FSP-3 using light-related strategies. Bioresour. Technol. 2014, 152, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Del Campo, J.A.; Moreno, J.; Rodrı́guez, H.; Vargas, M.A.; Rivas, J.; Guerrero, M.G. Carotenoid content of chlorophycean microalgae: Factors determining lutein accumulation in Muriellopsis sp. (Chlorophyta). J. Biotechnol. 2000, 76, 51–59. [Google Scholar] [CrossRef]
- Chen, C.-R.; Hong, S.-E.; Wang, Y.-C.; Hsu, S.-L.; Hsiang, D.; Chang, C.-M.J. Preparation of highly pure zeaxanthin particles from sea water-cultivated microalgae using supercritical anti-solvent recrystallization. Bioresour. Technol. 2012, 104, 828–831. [Google Scholar] [CrossRef] [PubMed]
- Granado-Lorencio, F.; Herrero-Barbudo, C.; Acién-Fernández, G.; Molina-Grima, E.; Fernández-Sevilla, J.; Pérez-Sacristán, B.; Blanco-Navarro, I. In vitro bioaccesibility of lutein and zeaxanthin from the microalgae Scenedesmus almeriensis. Food Chem. 2009, 114, 747–752. [Google Scholar] [CrossRef]
- Chen, F.; Li, H.-B.; Wong, R.N.S.; Ji, B.; Jiang, Y. Isolation and purification of the bioactive carotenoid zeaxanthin from the microalga Microcystis aeruginosa by high-speed counter-current chromatography. J. Chromatogr. A 2005, 1064, 183–186. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Sevilla, J.M.; Fernández, F.A.; Grima, E.M. Biotechnological production of lutein and its applications. Appl. Microbiol. Biotechnol. 2010, 86, 27–40. [Google Scholar] [CrossRef] [PubMed]
- Manke Natchigal, A.; Oliveira Stringheta, A.; Corrêa Bertoldi, M.; Stringheta, P. Quantification and characterization of lutein from Tagetes (Tagetes patula L.) and Calendula (Calendula officinalis L.) flowers. In Proceedings of the XXVIII International Horticultural Congress on Science and Horticulture for People (IHC2010): International Symposium on Emerging Health Topics in Fruits and Vegetables, Lisborn, Portugal, 22–27 August 2010; pp. 309–314. [Google Scholar]
- Ben-Amotz, A.; Avron, M. Photosynthetic activities of the halophilic alga Dunaliella parva. Plant Physiol. 1972, 49, 240–243. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Kim, M.; Lee, S.-G.; Lee, Y.; Choi, H.-K.; Jin, E. Contrasting photoadaptive strategies of two morphologically distinct Dunaliella species under various salinities. J. Appl. Phycol. 2015, 27, 1053–1062. [Google Scholar] [CrossRef]
- Borowitzka, M.A.; Borowitzka, L.J.; Kessly, D. Effects of salinity increase on carotenoid accumulation in the green alga Dunaliella salina. J. Appl. Phycol. 1990, 2, 111–119. [Google Scholar] [CrossRef]
- Pal, D.; Khozin-Goldberg, I.; Didi-Cohen, S.; Solovchenko, A.; Batushansky, A.; Kaye, Y.; Sikron, N.; Samani, T.; Fait, A.; Boussiba, S. Growth, lipid production and metabolic adjustments in the euryhaline eustigmatophyte Nannochloropsis oceanica CCALA 804 in response to osmotic downshift. Appl. Microbiol. Biotechnol. 2013, 97, 8291–8306. [Google Scholar] [CrossRef] [PubMed]
- Aburai, N.; Sumida, D.; Abe, K. Effect of light level and salinity on the composition and accumulation of free and ester-type carotenoids in the aerial microalga Scenedesmus sp. (Chlorophyceae). Algal Res. 2015, 8, 30–36. [Google Scholar] [CrossRef]
- Jin, E.; Feth, B.; Melis, A. A mutant of the green alga Dunaliella salina constitutively accumulates zeaxanthin under all growth conditions. Biotechnol. Bioeng. 2003, 81, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Jin, E.; Yokthongwattana, K.; Polle, J.E.; Melis, A. Role of the reversible xanthophyll cycle in the photosystem II damage and repair cycle in Dunaliella salina. Plant Physiol. 2003, 132, 352–364. [Google Scholar] [CrossRef] [PubMed]
- Thaipratum, R.; Melis, A.; Svasti, J.; Yokthongwattana, K. Analysis of non-photochemical energy dissipating processes in wild type Dunaliella salina (green algae) and in zea1, a mutant constitutively accumulating zeaxanthin. J. Plant Res. 2009, 122, 465–476. [Google Scholar] [CrossRef] [PubMed]
- Hieber, A.D.; Bugos, R.C.; Yamamoto, H.Y. Plant lipocalins: Violaxanthin de-epoxidase and zeaxanthin epoxidase. Biochim. Biophys. Acta Protein Struct. Mol. Enzymol. 2000, 1482, 84–91. [Google Scholar] [CrossRef]
- Cebollero, E.; Gonzalez-Ramos, D.; Tabera, L.; Gonzalez, R. Transgenic wine yeast technology comes of age: Is it time for transgenic wine? Biotechnol. Lett. 2007, 29, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Haughn, G.; Somerville, C. Selection for herbicide resistance at the whole-plant level. In Biotechnology in Agricultural Chemistry; Oxford University Press: Oxford, UK, 1987; pp. 98–107. [Google Scholar]
- Ashburner, M. Drosophila: A Laboratory Handbook; Cold Spring Harbor Laboratory: Englewood Cliffs, NJ, USA, 1989.
- Pérez-Torrado, R.; Querol, A.; Guillamón, J.M. Genetic improvement of non-GMO wine yeasts: Strategies, advantages and safety. Trends Food Sci. Technol. 2015, 45, 1–11. [Google Scholar] [CrossRef]
- Chaturvedi, R.; Fujita, Y. Isolation of enhanced eicosapentaenoic acid producing mutants of Nannochloropsis oculata ST-6 using ethyl methane sulfonate induced mutagenesis techniques and their characterization at mRNA transcript level. Phycol. Res. 2006, 54, 208–219. [Google Scholar] [CrossRef]
- Ong, S.-C.; Kao, C.-Y.; Chiu, S.-Y.; Tsai, M.-T.; Lin, C.-S. Characterization of the thermal-tolerant mutants of Chlorella sp. with high growth rate and application in outdoor photobioreactor cultivation. Bioresour. Technol. 2010, 101, 2880–2883. [Google Scholar] [CrossRef] [PubMed]
- Anandarajah, K.; Mahendraperumal, G.; Sommerfeld, M.; Hu, Q. Characterization of microalga Nannochloropsis sp. mutants for improved production of biofuels. Appl. Energy 2012, 96, 371–377. [Google Scholar] [CrossRef]
- Zhang, Y.; He, M.; Zou, S.; Fei, C.; Yan, Y.; Zheng, S.; Rajper, A.A.; Wang, C. Breeding of high biomass and lipid producing Desmodesmus sp. by Ethylmethane sulfonate-induced mutation. Bioresour. Technol. 2016, 207, 268–275. [Google Scholar] [CrossRef] [PubMed]
- Melis, A. Solar energy conversion efficiencies in photosynthesis: Minimizing the chlorophyll antennae to maximize efficiency. Plant Sci. 2009, 177, 272–280. [Google Scholar] [CrossRef]
- Jeong, J.; Baek, K.; Kirst, H.; Melis, A.; Jin, E. Loss of CpSRP54 function leads to a truncated light-harvesting antenna size in Chlamydomonas reinhardtii. Biochim. Biophys. Acta Bioenerg. 2017, 1858, 45–55. [Google Scholar] [CrossRef] [PubMed]
- Ahrazem, O.; Rubio-Moraga, A.; López, R.C.; Gómez-Gómez, L. The expression of a chromoplast-specific lycopene beta cyclase gene is involved in the high production of saffron’s apocarotenoid precursors. J. Exp. Bot. 2010, 61, 105–119. [Google Scholar] [CrossRef] [PubMed]
- Davison, P.; Hunter, C.; Horton, P. Overexpression of β-carotene hydroxylase enhances stress tolerance in Arabidopsis. Nature 2002, 418, 203–206. [Google Scholar] [CrossRef] [PubMed]
- Baek, K.; Kim, D.H.; Jeong, J.; Sim, S.J.; Melis, A.; Kim, J.-S.; Jin, E.; Bae, S. DNA-free two-gene knockout in Chlamydomonas reinhardtii via CRISPR-Cas9 ribonucleoproteins. Sci. Rep. 2016, 6, 30620. [Google Scholar] [CrossRef] [PubMed]
- Minhas, A.K.; Hodgson, P.; Barrow, C.J.; Adholeya, A. A review on the assessment of stress conditions for simultaneous production of microalgal lipids and carotenoids. Front. Microbiol. 2016, 7, 546. [Google Scholar] [CrossRef] [PubMed]
- Kirst, G. Salinity tolerance of eukaryotic marine algae. Annu. Rev. Plant Biol. 1990, 41, 21–53. [Google Scholar] [CrossRef]
- Müller, P.; Li, X.-P.; Niyogi, K.K. Non-photochemical quenching. A response to excess light energy. Plant Physiol. 2001, 125, 1558–1566. [Google Scholar] [CrossRef] [PubMed]
- Pulz, O. Photobioreactors: Production systems for phototrophic microorganisms. Appl. Microbiol. Biotechnol. 2001, 57, 287–293. [Google Scholar] [PubMed]
- Tang, H.; Abunasser, N.; Garcia, M.; Chen, M.; Ng, K.S.; Salley, S.O. Potential of microalgae oil from Dunaliella tertiolecta as a feedstock for biodiesel. Appl. Energy 2011, 88, 3324–3330. [Google Scholar] [CrossRef]
- Sharma, Y.C.; Singh, B.; Korstad, J. A critical review on recent methods used for economically viable and eco-friendly development of microalgae as a potential feedstock for synthesis of biodiesel. Green Chem. 2011, 13, 2993–3006. [Google Scholar] [CrossRef]
- Cheirsilp, B.; Torpee, S. Enhanced growth and lipid production of microalgae under mixotrophic culture condition: Effect of light intensity, glucose concentration and fed-batch cultivation. Bioresour. Technol. 2012, 110, 510–516. [Google Scholar] [CrossRef] [PubMed]
- De Swaaf, M.E.; Sijtsma, L.; Pronk, J.T. High-cell-density fed-batch cultivation of the docosahexaenoic acid producing marine alga Crypthecodinium cohnii. Biotechnol. Bioeng. 2003, 81, 666–672. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.M.; Chen, F. High-yield production of lutein by the green microalga Chlorella protothecoidesin heterotrophic fed-batch culture. Biotechnol. Prog. 2002, 18, 723–727. [Google Scholar] [CrossRef] [PubMed]
- Moros, E.; Darnoko, D.; Cheryan, M.; Perkins, E.; Jerrell, J. Analysis of xanthophylls in corn by HPLC. J. Agric. Food Chem. 2002, 50, 5787–5790. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Leung, I.; Tso, M.O.; Lam, K.-W. The identification of dipalmityl zeaxanthin as the major carotenoid in Gou Qi Zi by high pressure liquid chromatography and mass spectrometry. J. Ocul. Pharmacol. Ther. 1999, 15, 557–565. [Google Scholar] [CrossRef] [PubMed]
- Cordero, B.F.; Obraztsova, I.; Couso, I.; Leon, R.; Vargas, M.A.; Rodriguez, H. Enhancement of lutein production in Chlorella sorokiniana (Chorophyta) by improvement of culture conditions and random mutagenesis. Mar. Drugs 2011, 9, 1607–1624. [Google Scholar] [CrossRef] [PubMed]
- Römer, S.; Lübeck, J.; Kauder, F.; Steiger, S.; Adomat, C.; Sandmann, G. Genetic engineering of a zeaxanthin-rich potato by antisense inactivation and co-suppression of carotenoid epoxidation. Metab. Eng. 2002, 4, 263–272. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Polle, J.E.; Melis, A.; Lee, T.K.; Jin, E. Up-regulation of photoprotection and PSII-repair gene expression by irradiance in the unicellular green alga Dunaliella salina. Mar. Biotechnol. 2006, 8, 120–128. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Park, S.; Polle, J.E.; Jin, E. Gene expression profiling of Dunaliella sp. acclimated to different salinities. Phycol. Res. 2010, 58, 17–28. [Google Scholar] [CrossRef]
- Pick, U.; Karni, L.; Avron, M. Determination of ion content and ion fluxes in the halotolerant alga Dunaliella salina. Plant Physiol. 1986, 81, 92–96. [Google Scholar] [CrossRef] [PubMed]
- Levasseur, M.; Thompson, P.A.; Harrison, P.J. Physiological acclimation of marine phytoplankton to different nitrogen sources. J. Phycol. 1993, 29, 587–595. [Google Scholar] [CrossRef]
- Park, S.; Jung, G.; Hwang, Y.-S.; Jin, E. Dynamic response of the transcriptome of a psychrophilic diatom, Chaetoceros neogracile, to high irradiance. Planta 2010, 231, 349–360. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | [NaCl] | Carotenoid Pigments (μg·106·Cells−1) | |||||
|---|---|---|---|---|---|---|---|
| Neoxanthin | Violaxanthin | Antheraxanthin | Lutein | Zeaxanthin | β-Carotene | ||
| wild type | 0.3 M | 0.112 (±0.008) | 0.157 (±0.006) | 0.061 (±0.005) | 0.507 (±0.022) | 0.079 (±0.005) | 0.172 (±0.011) |
| 0.6 M | 0.101 (±0.018) | 0.112 (±0.014) | 0.061 (±0.007) | 0.414 (±0.012) | 0.083 (±0.007) | 0.133 (±0.013) | |
| 1.5 M | 0.096 (±0.018) | 0.074 (±0.005) | 0.057 (±0.003) | 0.505 (±0.059) | 0.117 (±0.013) | 0.143 (±0.010) | |
| 3.0 M | 0.122 (±0.010) | 0.086 (±0.004) | 0.074 (±0.004) | 0.433 (±0.027) | 0.131 (±0.006) | 0.149 (±0.013) | |
| zea1 | 0.3 M | N.D. | N.D. | N.D. | 0.358 (±0.017) | 0.246 (±0.011) | 0.186 (±0.008) |
| 0.6 M | N.D. | N.D. | N.D. | 0.459 (±0.025) | 0.303 (±0.038) | 0.184 (±0.004) | |
| 1.5 M | N.D. | N.D. | N.D. | 0.502 (±0.058) | 0.374 (±0.012) | 0.185 (±0.010) | |
| 3.0 M | N.D. | N.D. | N.D. | 0.402 (±0.025) | 0.359 (±0.038) | 0.167 (±0.010) | |
| mp3 | 0.3 M | N.D. | N.D. | N.D. | 0.272 (±0.014) | 0.262 (±0.019) | 0.130 (±0.001) |
| 0.6 M | N.D. | N.D. | N.D. | 0.329 (±0.016) | 0.368 (±0.013) | 0.171 (±0.015) | |
| 1.5 M | N.D. | N.D. | N.D. | 0.346 (±0.035) | 0.389 (±0.012) | 0.173 (±0.008) | |
| 3.0 M | N.D. | N.D. | N.D. | 0.268 (±0.024) | 0.312 (±0.029) | 0.133 (±0.013) | |
| Strains | Irradiance | Carotenoid Pigments (μg·106·Cells−1) | |||||
|---|---|---|---|---|---|---|---|
| Neoxanthin | Violaxanthin | Antheraxanthin | Lutein | Zeaxanthin | β-Carotene | ||
| wild type | Low-light | 0.101 (±0.018) | 0.112 (±0.014) | 0.061 (±0.007) | 0.414 (±0.012) | 0.083 (±0.007) | 0.133 (±0.013) |
| Mid-light | 0.069 (±0.005) | 0.091 (±0.003) | 0.056 (±0.004) | 0.324 (±0.023) | 0.066 (±0.006) | 0.119 (±0.006) | |
| High-light | 0.051 (±0.002) | 0.072 (±0.004) | 0.041 (±0.002) | 0.194 (±0.016) | 0.043 (±0.001) | 0.088 (±0.003) | |
| zea1 | Low-light | N.D. | N.D. | N.D. | 0.459 (±0.025) | 0.303 (±0.038) | 0.184 (±0.004) |
| Mid-light | N.D. | N.D. | N.D. | 0.336 (±0.016) | 0.253 (±0.012) | 0.159 (±0.013) | |
| High-light | N.D. | N.D. | N.D. | 0.236 (±0.006) | 0.149 (±0.004) | 0.079 (±0.003) | |
| mp3 | Low-light | N.D. | N.D. | N.D. | 0.329 (±0.016) | 0.368 (±0.013) | 0.171 (±0.015) |
| Mid-light | N.D. | N.D. | N.D. | 0.240 (±0.003) | 0.267 (±0.005) | 0.174 (±0.023) | |
| High-light | N.D. | N.D. | N.D. | 0.173 (±0.005) | 0.209 (±0.002) | 0.080 (±0.014) | |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, M.; Ahn, J.; Jeon, H.; Jin, E. Development of a Dunaliella tertiolecta Strain with Increased Zeaxanthin Content Using Random Mutagenesis. Mar. Drugs 2017, 15, 189. https://doi.org/10.3390/md15060189
Kim M, Ahn J, Jeon H, Jin E. Development of a Dunaliella tertiolecta Strain with Increased Zeaxanthin Content Using Random Mutagenesis. Marine Drugs. 2017; 15(6):189. https://doi.org/10.3390/md15060189
Chicago/Turabian StyleKim, Minjae, Junhak Ahn, Hancheol Jeon, and EonSeon Jin. 2017. "Development of a Dunaliella tertiolecta Strain with Increased Zeaxanthin Content Using Random Mutagenesis" Marine Drugs 15, no. 6: 189. https://doi.org/10.3390/md15060189
APA StyleKim, M., Ahn, J., Jeon, H., & Jin, E. (2017). Development of a Dunaliella tertiolecta Strain with Increased Zeaxanthin Content Using Random Mutagenesis. Marine Drugs, 15(6), 189. https://doi.org/10.3390/md15060189