Pufferfish Saxitoxin and Tetrodotoxin Binding Protein (PSTBP) Analogues in the Blood Plasma of the Pufferfish Arothron nigropunctatus, A. hispidus, A. manilensis, and Chelonodon patoca

Abstract
:1. Introduction
2. Results
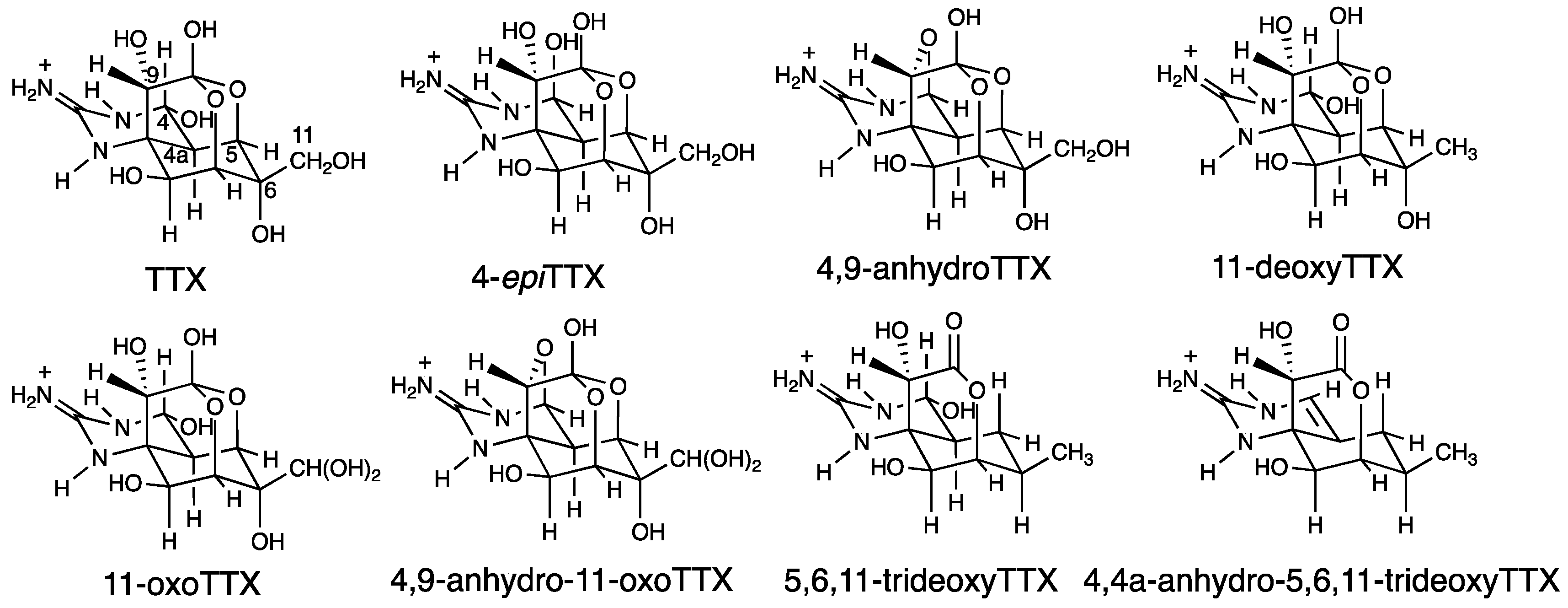
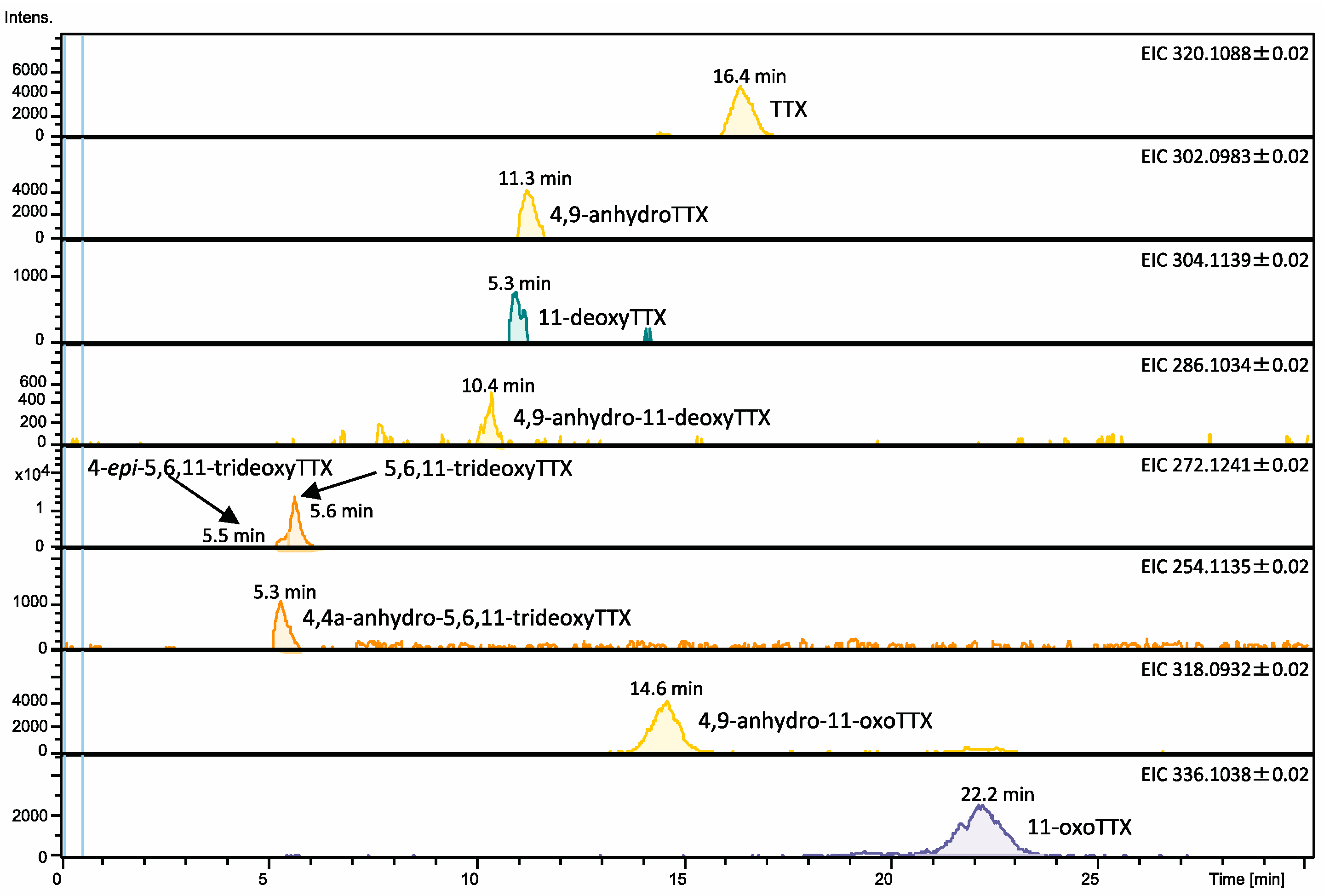
2.1. TTX and Its Analogues in Pufferfish Plasma
2.2. The Ratios of TTXs Bound to Proteins to Total TTXs in Pufferfish Plasma
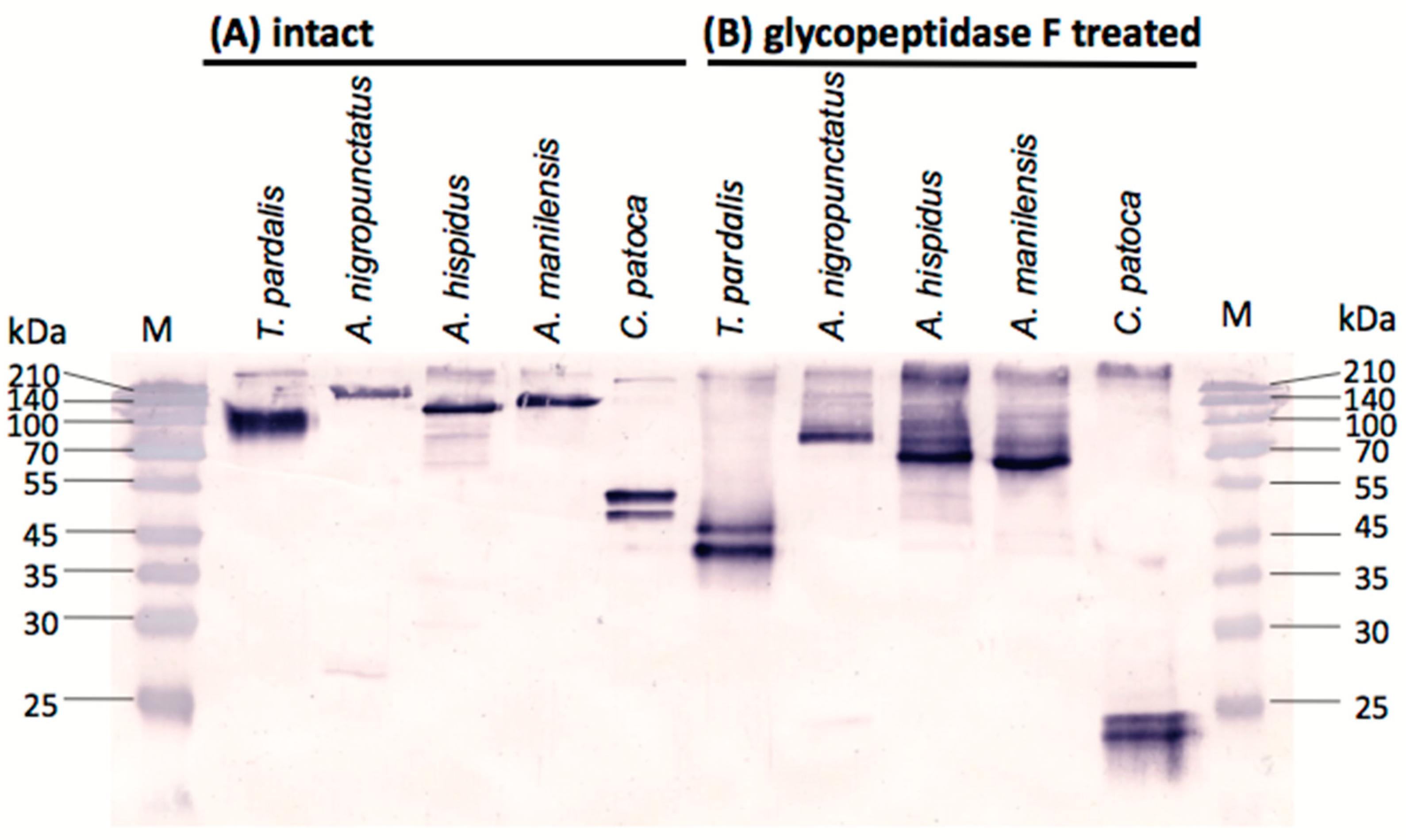
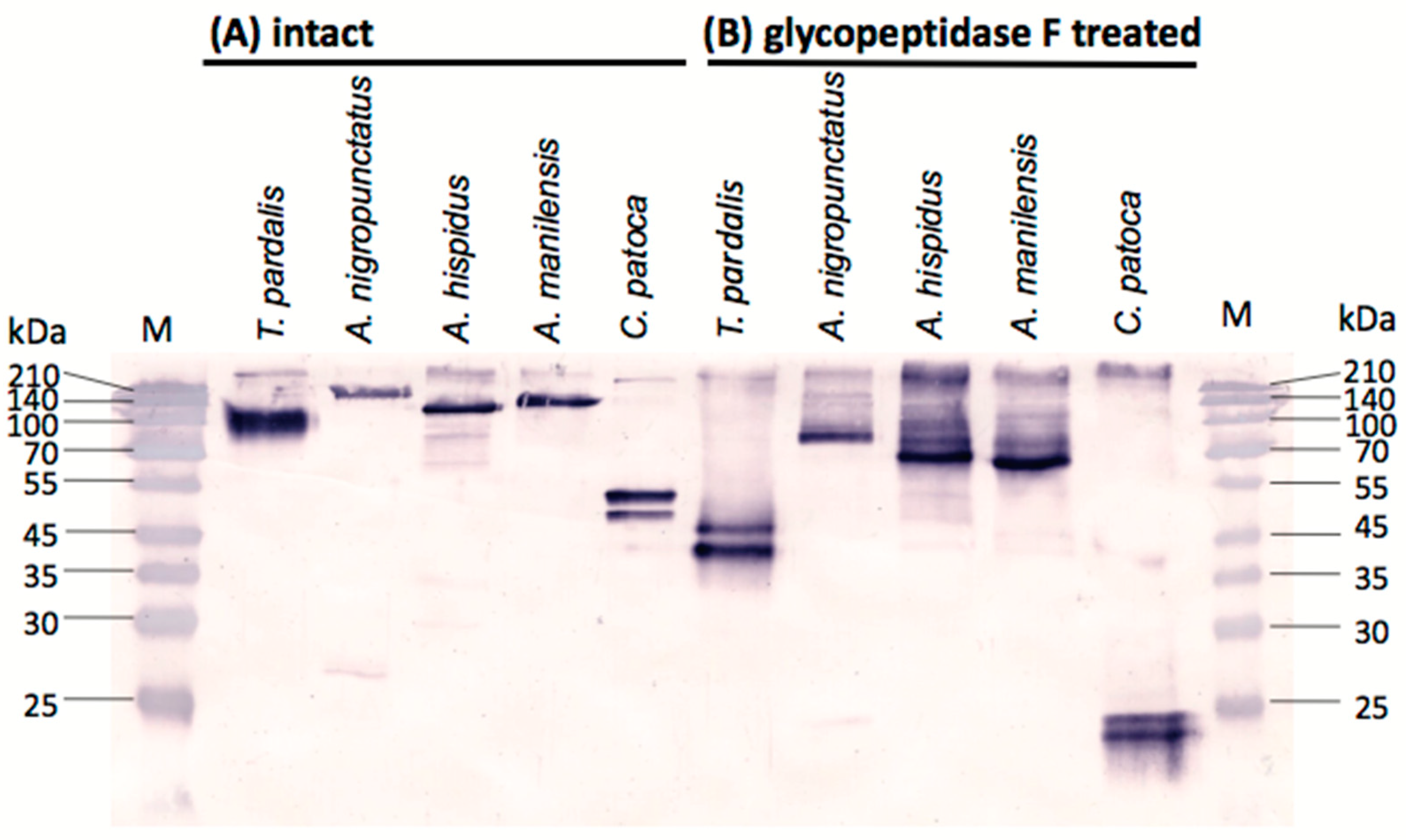
2.3. Western Blot Analyses of Pufferfish Plasma with Anti-PSTBP Antibody Detection
2.4. N-Terminal Amino Acid Sequences of PSTBP-Like Proteins
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Quantitation of the TTXs in Pufferfish Plasma
4.3. Reactions of Plasma with Glycopeptidase F
4.4. Western Blot Analysis
4.5. N-Terminal Amino Acid Sequences Analysis
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Yotsu-Yamashita, M.; Sugimoto, A.; Terakawa, T.; Shoji, Y.; Miyazawa, T.; Yasumoto, T. Purification, characterization, and cDNA cloning of a novel soluble saxitoxin and tetrodotoxin binding protein from plasma of the pufferfish, Fugu pardalis. Eur. J. Biochem. 2001, 268, 5937–5946. [Google Scholar] [CrossRef] [PubMed]
- Matsui, T.; Yamamori, K.; Furukawa, K.; Kono, M. Purification and some properties of a tetrodotoxin binding protein from the blood plasma of kusafugu. Takifugu niphobles. Toxicon 2000, 38, 463–468. [Google Scholar] [CrossRef]
- Yotsu-Yamashita, M.; Shoji, Y.; Terakawa, T.; Yamada, S.; Miyazawa, T.; Yasumoto, T. Mutual binding inhibition of tetrodotoxin and saxitoxin to their binding protein from the plasma of the pufferfish. Fugu pardalis. Biosci. Biotechnol. Biochem. 2002, 66, 2520–2524. [Google Scholar] [CrossRef] [PubMed]
- Yotsu-Yamashita, M.; Yamaki, H.; Okoshi, N.; Araki, N. Distribution of homologous proteins to pufferfish saxitoxin and tetrodotoxin binding protein in the plasma of pufferfish and among the tissues of Fugu pardalis examined by Western blot analysis. Toxicon 2010, 55, 1119–1124. [Google Scholar] [CrossRef] [PubMed]
- Yotsu-Yamashita, M.; Okoshi, N.; Watanabe, K.; Araki, N.; Yamaki, H.; Shoji, Y.; Terakawa, T. Localization of pufferfish saxitoxin and tetrodotoxin binding protein (PSTBP) in the tissues of the pufferfish, Takifugu pardalis, analyzed by immunohistochemical staining. Toxicon 2013, 72, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Oba, Y.; Shimasaki, Y.; Oshima, Y.; Satone, H.; Kitano, T.; Nakao, M.; Kawabata, S.; Honjo, T. Purification and characterization of tributyltin-binding protein type 2 from plasma of Japanese flounder, Paralichthys olivaceus. J. Biochem. 2007, 142, 229–238. [Google Scholar] [CrossRef] [PubMed]
- Satone, H.; Akahoshi, E.; Nakamura, A.; Lee, J.M.; Honda, M.; Shimasaki, Y.; Kawabata, S.; Kusakabe, T.; Oshima, Y. Expression and functional characterization of recombinant tributyltin-binding protein type 2. J. Toxicol. Sci. 2013, 38, 885–890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashiguchi, Y.; Lee, J.M.; Shiraishi, M.; Komatsu, S.; Miki, S.; Shimasaki, Y.; Mochioka, N.; Kusakabe, T.; Oshima, Y. Characterization and evolutionary analysis of tributyltin-binding protein and pufferfish saxitoxin and tetrodotoxin-binding protein genes in toxic and nontoxic pufferfishes. J. Evol. Biol. 2015, 28, 1103–1118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Satone, H.; Nonaka, S.; Lee, J.M.; Shimasaki, Y.; Kusakabe, T.; Kawabata, S.; Oshima, Y. Tetrodotoxin- and tributyltin-binding abilities of recombinant pufferfish saxitoxin and tetrodotoxin binding proteins of Takifugu rubripes. Toxicon 2017, 125, 50–52. [Google Scholar] [CrossRef] [PubMed]
- Noguchi, T.; Arakawa, O. Tetrodotoxin—Distribution and accumulation in aquatic organisms, and cases of human intoxication. Mar. Drugs 2008, 6, 220–242. [Google Scholar] [CrossRef] [PubMed]
- Teruya, N.; Oshiro, N.; Tamanaha, K. Toxicity of puffers from Okinawa. Annu. Rep. Okinawa Prefect. Inst. Health Environ. 2006, 40, 93–97. (In Japanese) [Google Scholar]
- Puilingi, C.G.; Kudo, Y.; Cho, Y.; Konoki, K.; Yotsu-Yamashita, M. Tetrodotoxin and its analogues in the pufferfish Arothron hispidus, and A. nigropunctatus from the Solomon Islands: A comparison of their toxin profiles with the same species from Okinawa, Japan. Toxins 2015, 7, 3436–3454. [Google Scholar] [CrossRef] [PubMed]
- Mahmud, Y.; Okada, K.; Takatani, T.; Kawatsu, K.; Hamano, Y.; Arakawa, O.; Noguchi, T. Intra-tissue distribution of tetrodotoxin in two marine puffers Takifugu vermicularis and Chelonodon patoca. Toxicon 2003, 41, 13–18. [Google Scholar] [CrossRef]
- Yotsu-Yamashita, M. Chemistry of puffer fish toxin. J. Toxicol. Toxin Rev. 2001, 20, 51–66. [Google Scholar] [CrossRef]
- Saruhashi, S.; Konoki, K.; Yotsu-Yamashita, M. The voltage-gated sodium ion channel inhibitory activities of a new tetrodotoxin analogue, 4,4a-anhydrotetrodotoxin, and three other analogues evaluated by colorimetric cell-based assay. Toxicon 2016, 119, 72–76. [Google Scholar] [CrossRef] [PubMed]
- Yasumoto, T.; Michishita, T. Fluorometric determination of tetrodotoxin by high performance liquid chromatography. Agric. Biol. Chem. 1985, 49, 3077–3080. [Google Scholar]
- Shoji, Y.; Yotsu-Yamashita, M.; Miyazawa, T.; Yasumoto, T. Electrospray ionization mass spectrometry of tetrodotoxin and its analogs: Liquid chromatography/mass spectrometry, tandem mass spectrometry, and liquid chromatography/tandem mass spectrometry. Anal. Biochem. 2001, 290, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Yotsu-Yamashita, M.; Abe, Y.; Kudo, Y.; Ritson-Williams, R.; Paul, V.J.; Konoki, K.; Cho, Y.; Adachi, M.; Imazu, T.; Nishikawa, T. First identification of 5,11-dideoxytetrodotoxin in marine animals, and characterization of major fragment ions of tetrodotoxin and its analogs by high resolution ESI-MS/MS. Mar. Drugs 2013, 11, 2799–2813. [Google Scholar] [CrossRef] [PubMed]
- Yasumoto, T.; Yasumura, D.; Yotsu, M.; Michishita, T.; Endo, A.; Kotaki, Y. Bacterial production of tetrodotoxin and anhydrotetrodotoxin. Agric. Biol. Chem. 1986, 50, 793–795. [Google Scholar]
- Noguchi, T.; Jeo, J.; Arakawa, O.; Sugita, H.; Deguchi, Y.; Shida, Y.; Hashimoto, K. Occurrence of tetrodotoxin and anhydrotetrodotoxin in Vibrio sp. Isolated from the intestines of a xanthidcrab. Atergatis floridus. J. Biochem. 1986, 99, 311–314. [Google Scholar] [CrossRef] [PubMed]
- Turner, A.D.; Dhanji-Rapkova, M.; Coates, L.; Bickerstaff, L.; Milligan, S.; O’Neill, A.; Faulkner, D.; McEneny, H.; Baker-Austin, C.; Lees, D.N.; et al. Detection of tetrodotoxin shellfish poisoning (TSP) toxins and causative factors in bivalve molluscs from the UK. Mar. Drugs 2017, 15, 277. [Google Scholar] [CrossRef] [PubMed]
- Yasumoto, T.; Nagai, H.; Yasumura, D.; Michishita, T.; Endo, A.; Yotsu, M.; Kotaki, Y. Interspecies distribution and possible origin of tetrodotoxin. Ann. N. Y. Acad. Sci. 1986, 479, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Kono, M.; Matsui, T.; Furukawa, K.; Yotsu-Yamashita, M.; Yamamori, K. Accumulation of tetrodotoxin and 4,9-anhydrotetrodotoxin in cultured juvenile kusafugu Fugu niphobles by dietary administration of natural toxic komonfugu Fugu poecilonotus liver. Toxicon 2008, 51, 1269–1273. [Google Scholar] [CrossRef]
- Tatsuno, R.; Shikina, M.; Shirai, Y.; Wang, J.; Soyano, K.; Nishihara, J.N.; Takatani, T.; Arakawa, O. Change in the transfer profile of orally administered tetrodotoxin to non-toxic cultured pufferfish Takifugu rubripes depending of its development stage. Toxicon 2013, 65, 76–80. [Google Scholar] [CrossRef] [PubMed]
- Tatsuno, R.; Gao, W.; Ibi, K.; Mine, T.; Okita, K.; Nishihara, G.N.; Takatani, T.; Arakawa, O. Profile differences in tetrodotoxin transfer to skin and liver in the pufferfish Takifugu rubripes. Toxicon 2017, 130, 73–78. [Google Scholar] [CrossRef]
- Yin, X.; Kiriake, A.; Ohta, A.; Kitani, A.; Ishizaki, S.; Nagashima, Y. A novel function of vitellogenin subdomain, vWF type D, as a toxin-binding protein in the pufferfish Takifugu pardalis ovary. Toxicon 2017, 136, 56–66. [Google Scholar] [CrossRef] [PubMed]
- Nagashima, Y.; Ohta, A.; Yin, X.; Ishizaki, S.; Matsumoto, T.; Doi, H.; Ishibashi, T. Difference in uptake of tetrodotoxin and saxitoxins into liver tissue slices among pufferfish, boxfish and porcupinefish. Mar. Drugs 2018, 16, 17. [Google Scholar] [CrossRef] [PubMed]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Towbin, H.; Staehelin, T.; Gordon, J. Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: Procedure and some applications. Proc. Natl. Acad. Sci. USA 1979, 76, 4350–4354. [Google Scholar] [CrossRef] [PubMed]
- Kenmochi, E.; Kabir, S.R.; Ogawa, T.; Naude, R.; Tateno, H.; Hirabayashi, J.; Muramoto, K. Isolation and biochemical characterization of Apios tuber lectin. Molecules 2015, 20, 987–1002. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| A. nigropunctatus | A. hispidus | A. manilensis | C. patoca | |
|---|---|---|---|---|
| TTX | 150 | 91 | 890 | 1000 |
| 4-epiTTX | 78 | 12 | 60 | 110 |
| 4,9-anhydroTTX | 19 | 15 | 850 | 95 |
| 11-oxoTTX | 45 | ND | 1100 | 28 |
| 4,9-anhydro-11-oxoTTX | 96 | ND | 2800 | ND |
| 11-deoxyTTX | 120 | ND | 74 | 170 |
| 5,6,11-trideoxyTTX | 95 | 22 | 810 | 59 |
| 4,4a-anhydro-trideoxyTTX | 100 | 32 | 75 | 30 |
| A. nigropunctatus | A. hispidus | A. manilensis | C. patoca | |
|---|---|---|---|---|
| TTX | 32 | 26 | 53 | 12 |
| 4-epiTTX | ND | ND | ND | 17 |
| 4,9-anhydroTTX | ND | ND | 51 | ND |
| 11-oxoTTX | 68 | ND | 70 | ND |
| 4,9-anhydro-11-oxoTTX | ND | ND | 61 | ND |
| 11-deoxyTTX | 14 | ND | 29 | 3 |
| 5,6,11-trideoxyTTX | 66 | 72 | 75 | 100 |
| 4,4a-anhydro-5,6,11-trideoxyTTX | 76 | 82 | 85 | 100 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yotsu-Yamashita, M.; Nagaoka, Y.; Muramoto, K.; Cho, Y.; Konoki, K. Pufferfish Saxitoxin and Tetrodotoxin Binding Protein (PSTBP) Analogues in the Blood Plasma of the Pufferfish Arothron nigropunctatus, A. hispidus, A. manilensis, and Chelonodon patoca. Mar. Drugs 2018, 16, 224. https://doi.org/10.3390/md16070224
Yotsu-Yamashita M, Nagaoka Y, Muramoto K, Cho Y, Konoki K. Pufferfish Saxitoxin and Tetrodotoxin Binding Protein (PSTBP) Analogues in the Blood Plasma of the Pufferfish Arothron nigropunctatus, A. hispidus, A. manilensis, and Chelonodon patoca. Marine Drugs. 2018; 16(7):224. https://doi.org/10.3390/md16070224
Chicago/Turabian StyleYotsu-Yamashita, Mari, Yuuma Nagaoka, Koji Muramoto, Yuko Cho, and Keiichi Konoki. 2018. "Pufferfish Saxitoxin and Tetrodotoxin Binding Protein (PSTBP) Analogues in the Blood Plasma of the Pufferfish Arothron nigropunctatus, A. hispidus, A. manilensis, and Chelonodon patoca" Marine Drugs 16, no. 7: 224. https://doi.org/10.3390/md16070224
APA StyleYotsu-Yamashita, M., Nagaoka, Y., Muramoto, K., Cho, Y., & Konoki, K. (2018). Pufferfish Saxitoxin and Tetrodotoxin Binding Protein (PSTBP) Analogues in the Blood Plasma of the Pufferfish Arothron nigropunctatus, A. hispidus, A. manilensis, and Chelonodon patoca. Marine Drugs, 16(7), 224. https://doi.org/10.3390/md16070224