Occurrence and Properties of Thiosilvatins




Abstract
:1. Introduction
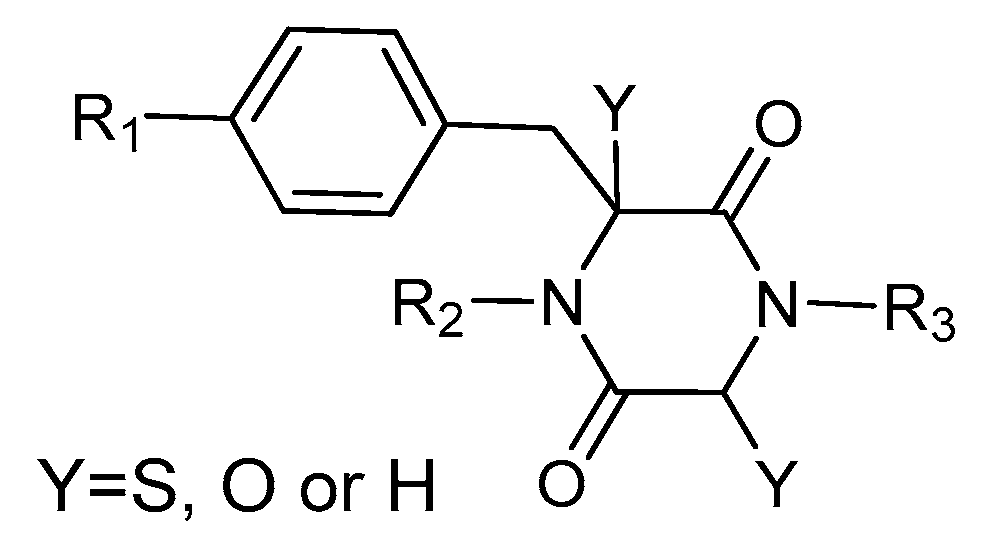
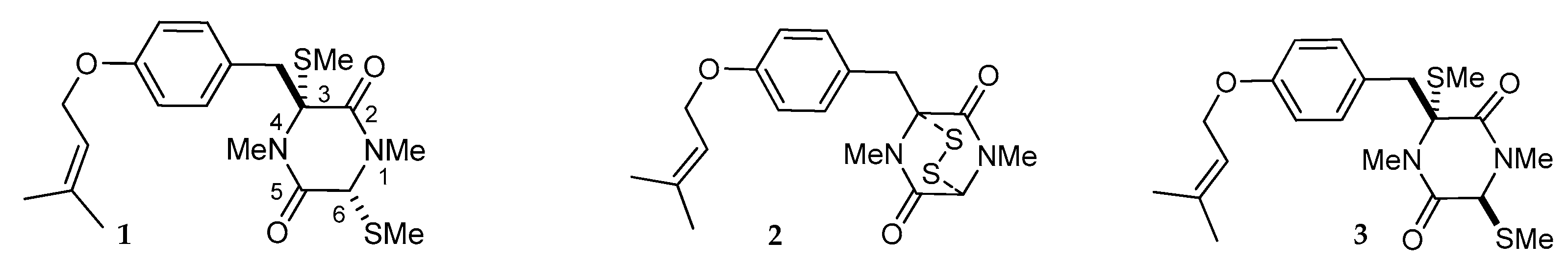
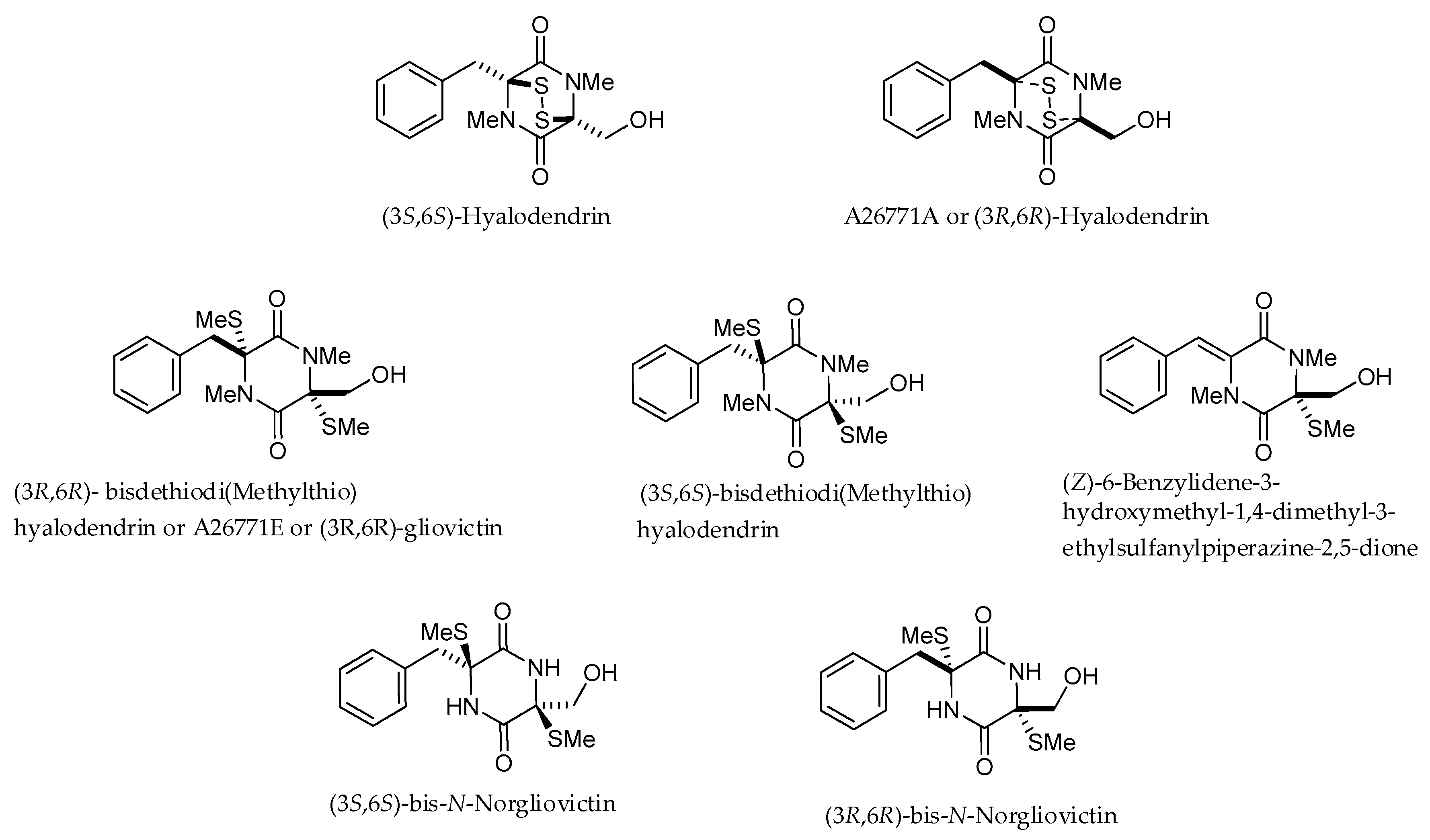
2. Structures and Chemical Properties
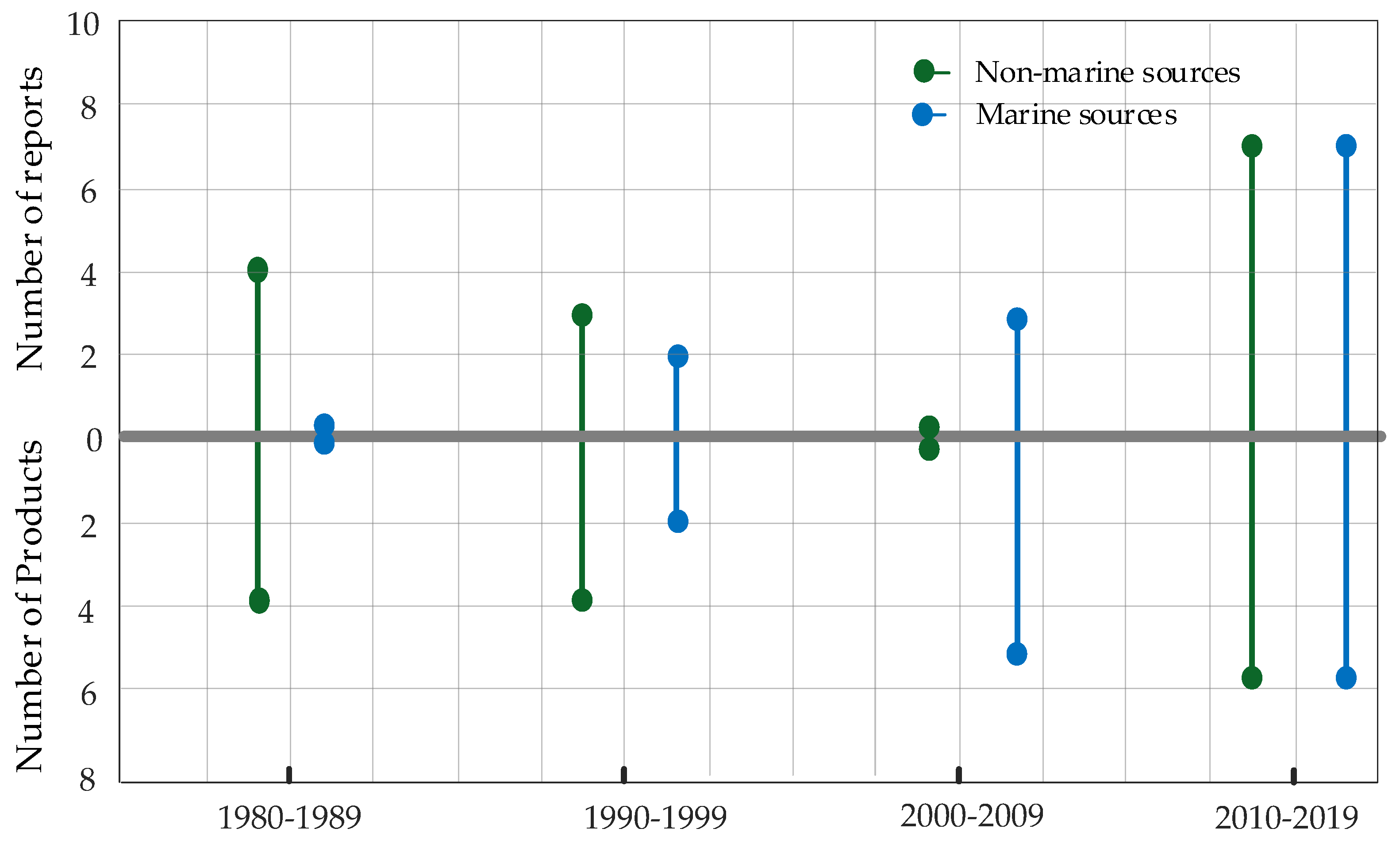
3. Fungal Sources
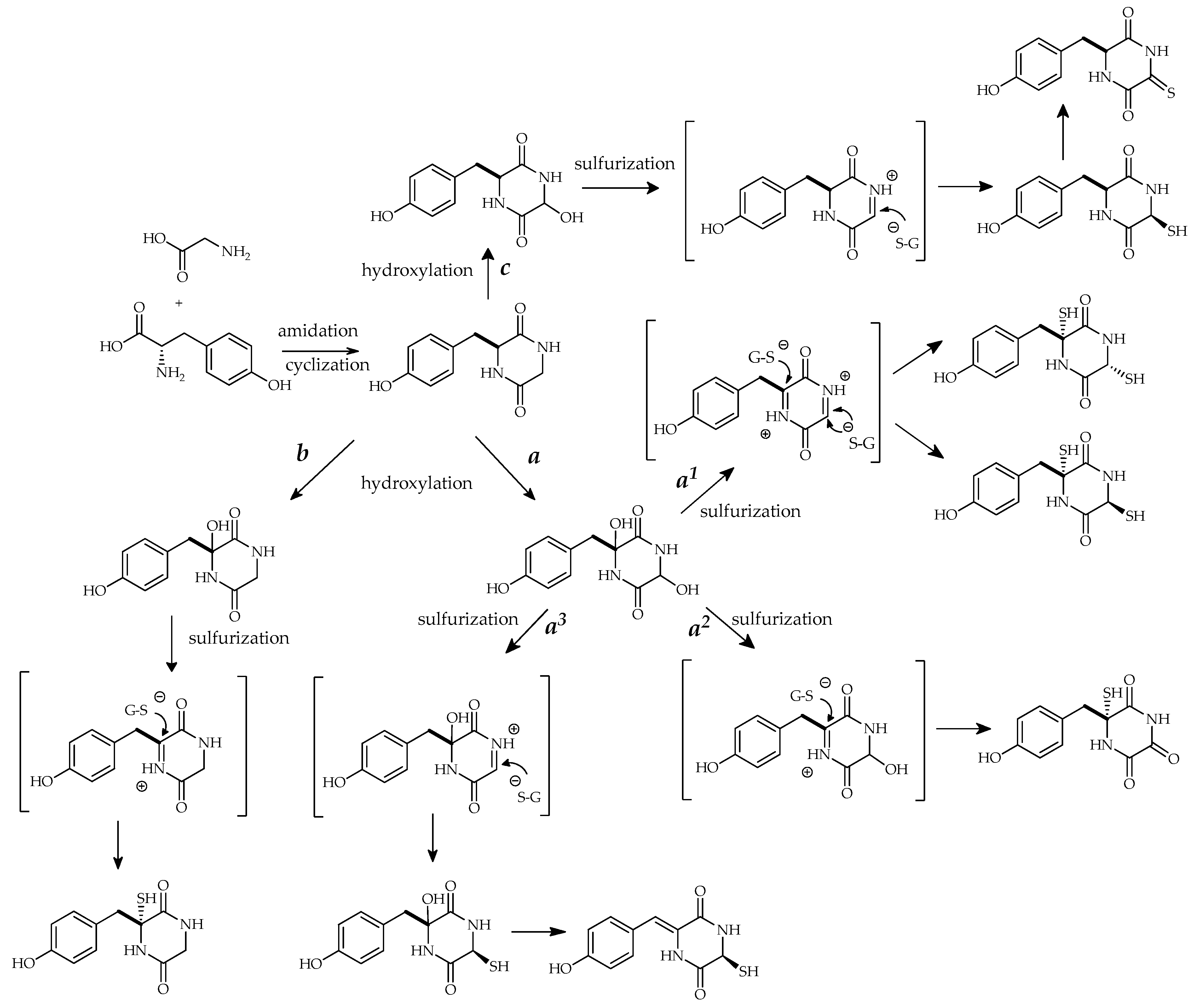
4. Proposed Biosynthetic Pathways for Thiosilvatins
5. Biological Activities
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Fox, E.M.; Howlett, B.J. Biosynthetic gene clusters for epipolythiodioxopiperazines in filamentous fungi. Mycol. Res. 2008, 112, 162–169. [Google Scholar] [CrossRef]
- Welch, T.R.; Williams, R.M. Epidithiodioxopiperazines. occurrence, synthesis and biogenesis. Nat. Prod. Rep. 2014, 31, 1376–1404. [Google Scholar] [CrossRef]
- Hanson, J.R.; O’Leary, M.A. New piperazinedione metabolites of Gliocladium deliquescens. J. Chem. Soc. Perkin Trans. 1 1981, 0, 218–220. [Google Scholar] [CrossRef]
- Kawahara, N.; Nozawa, K.; Nakajima, S.; Kawai, K.I. Studies on fungal products. Part 13. Studies on fungal products. Part 13. Isolation and structures of dithiosilvatin and silvathione, novel dioxopiperazine derivatives from Aspergillus silvaticus. J. Chem. Soc. Perkin Trans. 1 1987, 0, 2099–2101. [Google Scholar] [CrossRef]
- Chu, M.; Mierzwa, R.; Truumees, I.; Gentile, F.; Patel, M.; Gullo, V.; Chan, T.-M.; Puar, M.S. Two novel diketopiperazines isolated from the fungus Tolypocladium sp. Tetrahedron Lett. 1993, 34, 7537–7540. [Google Scholar] [CrossRef]
- Shi, Z.Z.; Miao, F.P.; Fang, S.T.; Yin, X.L.; Ji, N.Y. Sulfurated diketopiperazines from an algicolous isolate of Trichoderma virens. Phytochem. Let. 2018, 27, 101–104. [Google Scholar] [CrossRef]
- He, X.; Yang, J.; Qiu, L.; Feng, D.; Ju, F.; Tan, L.; Li, Y.-Z.; Gu, Y.-C.; Zhang, Z.; Guo, D.-L.; et al. Thiodiketopiperazines produced by Penicillium crustosum and their activities to promote gastrointestinal motility. Molecules 2019, 24, 299. [Google Scholar] [CrossRef] [PubMed]
- Usami, Y.; Aoki, S.; Hara, T.; Numata, A. New dioxopiperazine metabolites from a Fusarium species separated from a marine alga. J. Antibiot. 2002, 55, 655–659. [Google Scholar] [CrossRef] [PubMed]
- Amagata, T.; Minoura, K.; Numata, A. Cytotoxic metabolites produced by a fungal strain from a Sargassum alga. J. Antibiot. 1998, 51, 432–434. [Google Scholar] [CrossRef] [PubMed]
- Ayer, W.A.; Altena, I.V.; Browne, L.M. Three piperazinediones and a drimane diterpenoid from Penicillium brevi-compactum. Phytochemistry 1990, 29, 1661–1665. [Google Scholar] [CrossRef]
- Kirby, G.W.; Rao, G.V.; Robins, D.J. New co-metabolites of gliotoxin in Gliocladium virens. J. Chem. Soc. Perkin Trans. 1 1988, 2, 301–304. [Google Scholar] [CrossRef]
- DeVault, R.L.; Rosenbrook, W. A novel class of diketopiperazines. J. Antibiot. 1973, 26, 532–534. [Google Scholar] [CrossRef] [PubMed]
- Michel, K.H.; Chaney, M.O.; Jones, N.D.; Hoehn, M.M.; Nagarajan, R. Epipolythiopiperazinedione antibiotics from Penicillium turbatum. J. Antibiot. 1974, 27, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Strunz, G.M.; Kakushima, M.; Stillwell, M.A.; Heissner, C.J. Hyalodendrin: A new fungitoxic epidithiodioxopiperazine produced by a Hyalodendron species. J. Chem. Soc. Perkin Trans. 1 1973, 0, 2600–2602. [Google Scholar] [CrossRef]
- Isaka, M.; Palasarn, S.; Rachtawee, P.; Vimuttipong, S.; Kongsaeree, P. Unique diketopiperazine dimers from the insect pathogenic fungus Verticillium hemipterigenum BCC 1449. Org. Lett. 2005, 7, 2257–2260. [Google Scholar] [CrossRef] [PubMed]
- Nicoletti, R.; Trincone, A. Bioactive compounds produced by strains of Penicillium and Talaromyces of marine origin. Mar. Drugs 2016, 14, 37. [Google Scholar] [CrossRef] [PubMed]
- Rahbæk, L.; Sperry, S.; Piper, J.E.; Crews, P. Deoxynortrichoharzin, a new polyketide from the saltwater culture of a sponge-derived Paecilomyces fungus. J. Nat. Prod. 1998, 61, 1571–1573. [Google Scholar] [CrossRef]
- Rukachaisirikul, V.; Khamthong, N.; Sukpondma, Y.; Phongpaichit, S.; Hutadilok-Towatana, N.; Graidist, P.; Sakayaroj, J.; Kirtikara, K. Cyclohexene, diketopiperazine, lactone and phenol derivatives from the sea fan-derived fungi Nigrospora sp. PSU-F11 and PSU-F12. Arch. Pharm. Res. 2010, 33, 375–380. [Google Scholar] [CrossRef]
- Capon, R.J.; Stewart, M.; Ratnayake, R.; Lacey, E.; Gill, J.H. Citromycetins and bilains A–C: New aromatic polyketides and diketopiperazines from Australian marine-derived and terrestrial Penicillium spp. J. Nat. Prod. 2007, 70, 1746–1752. [Google Scholar] [CrossRef]
- Wang, J.; Liu, P.; Wang, Y.; Wang, H.; Li, J.; Zhuang, Y.; Zhu, W. Antimicrobial aromatic polyketides from gorgonian-associated fungus, Penicillium commune 518. Chin. J. Chem. 2012, 30, 1236–1242. [Google Scholar] [CrossRef]
- Liu, C.C.; Zhang, Z.Z.; Feng, Y.Y.; Gu, Q.Q.; Li, D.H.; Zhu, T.J. Secondary metabolites from Antarctic marine-derived fungus Penicillium crustosum HDN153086. Nat. Prod. Res. 2019, 33, 414–419. [Google Scholar] [CrossRef] [PubMed]
- Yurchenko, A.N.; Berdyshev, D.V.; Smetanina, O.F.; Ivanets, E.V.; Zhuravleva, O.I.; Rasin, A.B.; Khudyakova, Y.V.; Popov, R.S.; Dyshlovoy, S.A.; Amsberg, G.V.; et al. Citriperazines AD produced by a marine algae-derived fungus Penicillium sp. KMM 4672. Nat. Prod. Res. 2019, 1–6. [Google Scholar]
- Li, C.Y.; Ding, W.J.; Shao, C.L.; She, Z.G.; Lin, Y.C. Secondary metabolites of a marine mangrove fungus (Penicillium sp. No. 2556) from South China sea. J. Chin. Med. Mater. 2008, 31, 960–962. [Google Scholar]
- Li, F.; Guo, W.; Wu, L.; Zhu, T.; Gu, Q.; Li, D.; Che, Q. Saroclazines A–C, thio-diketopiperazines from mangrove-derived fungi Sarocladium kiliense HDN11-84. Arch. Pharm. Res. 2018, 41, 30–34. [Google Scholar] [CrossRef]
- Wang, G.-Y.-S.; Abrell, L.M.; Avelar, A.; Borgeson, B.M.; Crews, P. New hirsutane based sesquiterpenes from salt water cultures of a marine sponge-derived fungus and the terrestrial fungus Coriolus consors. Tetrahedron 1998, 54, 7335–7342. [Google Scholar] [CrossRef]
- Visagie, C.M.; Renaud, J.B.; Burgess, K.M.N.; Malloch, D.W.; Clark, D.; Ketch, L.; Urb, M.; Luis-Seize, G.; Assabguie, R.; Sumarah, M.W.; et al. Fifteen new species of Penicillium. Persoonia 2016, 36, 247–280. [Google Scholar] [CrossRef] [PubMed]
- Guimarães, D.O.; Borges, W.S.; Vieira, N.J.; De Oliveira, L.F.; Da Silva, C.H.; Lopes, N.P.; Dias, L.G.; Durán-Patrón, R.; Collado, I.G.; Pupo, M.T. Diketopiperazines produced by endophytic fungi found in association with two Asteraceae species. Phytochemistry 2010, 71, 1423–1429. [Google Scholar] [CrossRef]
- Wang, H.; Chen, G.; Liu, Y.; Hua, H.; Bai, J.; Pei, Y.H. 1H and 13C NMR assignments of two new piperazine-trione from the fungus Penicillium crustosum YN-HT-15. Magn. Reson. Chem. 2015, 53, 620–623. [Google Scholar] [CrossRef]
- Hammerl, R.; Frank, O.; Schmittnägel, T.; Ehrmann, M.A.; Hofmann, T. Functional metabolome analysis of Penicillium roqueforti by means of differential Off-Line LC–NMR. J. Agric. Food Chem. 2019, 67, 5135–5146. [Google Scholar] [CrossRef]
- Yang, M.H.; Li, T.X.; Wang, Y.; Liu, R.H.; Luo, J.; Kong, L.Y. Antimicrobial metabolites from the plant endophytic fungus Penicillium sp. Fitoterapia 2017, 116, 72–76. [Google Scholar] [CrossRef]
- Pang, K.L.; Overy, D.P.; Jones, E.G.; da Luz Calado, M.; Burgaud, G.; Walker, A.K.; Johnson, J.A.; Kerr, R.G.; Cha, H.-J.; Bills, G.F. ‘Marine fungi’ and ‘marine-derived fungi’ in natural product chemistry research: Toward a new consensual definition. Fungal Biol. Rev. 2016, 30, 163–175. [Google Scholar] [CrossRef]
- Nicoletti, R.; Andolfi, A. The Marine-Derived Filamentous Fungi in Biotechnology. In Grand Challenges in Marine Biotechnology; Springer: Cham, Switzerland, 2018; pp. 157–189. [Google Scholar]
- Davis, C.; Carberry, S.; Schrettl, M.; Singh, I.; Stephens, J.C.; Barry, S.M.; Kavanagh, K.; Challis, G.L.; Brougham, D.; Doyle, S. The role of glutathione S-transferase GliG in gliotoxin biosynthesis in Aspergillus fumigatus. Chem. Biol. 2011, 18, 542–552. [Google Scholar] [CrossRef] [PubMed]
- Stachelhaus, T.; Mootz, H.D.; Bergendahl, V.; Marahiel, M.A. Peptide bond formation in nonribosomal peptide biosynthesis catalytic role of the condensation domain. J. Biol. Chem. 1998, 273, 22773–22781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brakhage, A.A. Regulation of fungal secondary metabolism. Nat. Rev. Microbiol. 2013, 11, 21–32. [Google Scholar] [CrossRef] [PubMed]
- Khaldi, N.; Seifuddin, F.T.; Turner, G.; Haft, D.; Nierman, W.C.; Wolfe, K.H.; Fedorova, N.D. SMURF: Genomic mapping of fungal secondary metabolite clusters. Fungal Genet. Biol. 2010, 47, 736–741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gardiner, D.M.; Cozijnsen, A.J.; Wilson, L.M.; Pedras, M.S.C.; Howlett, B.J. The sirodesmin biosynthetic gene cluster of the plant pathogenic fungus Leptosphaeria maculans. Mol. Microbiol. 2004, 53, 1307–1318. [Google Scholar] [CrossRef]
- Gardiner, D.M.; Waring, P.; Howlett, B.J. The epipolythiodioxopiperazine (ETP) class of fungal toxins: Distribution, mode of action, functions and biosynthesis. Microbiology 2005, 151, 1021–1032. [Google Scholar] [CrossRef] [Green Version]
- Jiang, C.S.; Guo, Y.W. Epipolythiodioxopiperazines from fungi: Chemistry and bioactivities. Mini Rev. Med. Chem. 2011, 11, 728–745. [Google Scholar] [CrossRef]
- Gu, B.; He, S.; Yan, X.; Zhang, L. Tentative biosynthetic pathways of some microbial diketopiperazines. Appl. Microbiol. Biotechnol. 2013, 97, 8439–8453. [Google Scholar] [CrossRef]
- Scharf, D.H.; Brakhage, A.A.; Mukherjee, P.K. Gliotoxin–bane or boon? Environ. Microbiol. 2016, 18, 1096–1109. [Google Scholar] [CrossRef]
- Suhadolnik, R.J.; Chenoweth, R.G. Biosynthesis of Gliotoxin. I. 1 Incorporation of Phenylalanine-1-and-2-C14. J. ACS 1958, 80, 4391–4392. [Google Scholar]
- Winstead, J.A.; Suhadolnik, R.J. Biosnthesis of gliotoxin. II. 1, 2 Further studies on the incorporation of carbon-14 and tritium-labeled precursors. J. ACS 1960, 82, 1644–1647. [Google Scholar]
- Balibar, C.J.; Walsh, C.T. GliP, a multimodular nonribosomal peptide synthetase in Aspergillus fumigatus, makes the diketopiperazine scaffold of gliotoxin. Biochemistry 2006, 45, 15029–15038. [Google Scholar] [CrossRef] [PubMed]
- Stillwell, M.A.; Magasi, L.P.; Strunz, G.M. Production, isolation, and antimicrobial activity of hyalodendrin, a new antibiotic produced by a species of Hyalodendron. Can. J. Chem. 1974, 20, 759–764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, Y.; Dou, H.; Gong, W.; Liu, X.; Yu, Z.; Li, E.; Tan, R.; Hou, Y. Bis-N-norgliovictin, a small-molecule compound from marine fungus, inhibits LPS-induced inflammation in macrophages and improves survival in sepsis. Eur. J. Pharmacol. 2013, 705, 49–60. [Google Scholar] [CrossRef]
- Hemmi, K.; Shimazaki, N.; Shima, I.; Okamoto, M.; Keizo, Y.; Hashimoto, M. Chapter 2: PAF antagonists from microbial origin: Structure-activity relationship of diketopiperazine derivatives. In Handbook of PAF and PAF Antagonists; CRC Press: Boca Raton, FL, USA, 2019; p. 51689. [Google Scholar]
- Fitzpatrick, D.A. Horizontal gene transfer in fungi. FEMS Microbiol. Lett. 2012, 329, 1–8. [Google Scholar] [CrossRef]
- Manganiello, G.; Marra, R.; Staropoli, A.; Lombardi, N.; Vinale, F.; Nicoletti, R. The shifting mycotoxin profiles of endophytic Fusarium strains: A case study. Agriculture 2019, 9, 143. [Google Scholar] [CrossRef] [Green Version]
- Gardiner, D.; Howlett, B. Bioinformatic and expression analysis of the putative gliotoxin gene cluster of Aspergillus fumigatus. FEMS Microbiol. Lett. 2005, 248, 241–248. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Hu, P.; Pan, Y.; Zhu, Y.; Liu, X.; Che, Y.; Liu, G. Identification and characterization of the verticillin biosynthetic gene cluster in Clonostachys rogersoniana. Fungal Genet. Biol. 2017, 103, 25–33. [Google Scholar] [CrossRef]
- Cairns, T.; Meyer, V. In silico prediction and characterization of secondary metabolite biosynthetic gene clusters in the wheat pathogen Zymoseptoria tritici. BMC Genomics 2017, 18, 631. [Google Scholar] [CrossRef] [Green Version]
- Bolognesi, M.L.; Ai Tran, H.N.; Staderini, M.; Monaco, A.; López-Cobeñas, A.; Bongarzone, S.; Biarnés, X.; López-Alvarado, P.; Cabezas, N.; Caramelli, M.; et al. Discovery of a class of diketopiperazines as antiprion compounds. Chem. Med. Chem. 2010, 5, 1324–1334. [Google Scholar] [CrossRef] [PubMed]
- Fontaine, K.; Mounier, J.; Coton, E.; Hymery, N. Individual and combined effects of roquefortine C and mycophenolic acid on human monocytic and intestinal cells. World Mycot. J. 2016, 9, 51–62. [Google Scholar] [CrossRef]
- Hymery, N.; Mounier, J.; Coton, E. Effect of Penicillium roqueforti mycotoxins on Caco-2 cells: Acute and chronic exposure. Toxicol. Vitr. 2018, 48, 188–194. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Compound | Structure | Formula, Nominal Mass (U) |
|---|---|---|---|
| N-Demethyl analogues | |||
| 4 | Sch 54794; cis-dinor-bis(methylthio)silvatin |  | C18H24N2O3S2 380 |
| 5 | Sch 54796; trans-dinor-bis(methylthio)silvatin |  | C18H24N2O3S2 380 |
| 6 | Saroclazine A |  | C19H26N2O3S2 394 |
| 7 | Saroclazine B |  | C19H26N2O3S2 394 |
| Dethio analogues | |||
| 8 | Silvathione |  | C18H22N2O3S 346 |
| 9 | 3-(4-(3-Methyl-2-butenyloxy)benzyl)-3-(methylthio)-2,5-piperazinedione; dinor-methylthiosilvatin |  | C17H22N2O3S 334 |
| 10 | 6-Hydroxy-3-(4-(3-methyl-2-butenyloxy)benzyl)-3-(methylthio)piperazine-2,5-dione; dinor-hydroxy-methylthiosilvatin |  | C17H22N2O4S 350 |
| 11 | Fusaperazine B (relative stereochemistry) |  | C18H24N2O4S 364 |
| 12 | 1,4-Dimethyl-6-(4-(3-methyl-2-butenyloxy)benzyl)-6-methylsulfanyl-piperazine-2,3,5-trione; 6-oxo-methylthiosilvatin |  | C19H24N2O4S 376 |
| 13 | 6-(4-(3-Methyl-2-butenyloxy)benzyl)-6-methylsulfanyl-piperazine-2,3,5-trione; dinor-6-oxo-methylthiosilvatin |  | C17H20N2O4S 348 |
| 14 | Fusaperazine E |  | C19H24N2O3S 360 |
| 15 | Fusaperazine F |  | C19H24N2O3S 360 |
| Deprenyl analogues | |||
| 16 | cis-3-(4-Hydroxybenzyl)-1,4-dimethyl-3,6-bis(methylthio)-2,5-piperazinedione; cis-deprenyl-bis(methylthio)silvatin |  | C15H20N2O3S2 340 |
| 17 | trans-6-(4-Hydroxybenzyl)-1,4-dimethyl-3,6-bis(methylthio)piperazine-2,5-dione; trans-deprenyl-bis(methylthio)silvatin |  | C15H20N2O3S2 340 |
| 18 | Fusaperazine A |  | C13H16N2O3S2 312 |
| 19 | Citriperazine A |  | C13H16N2O2S2 296 |
| 20 | Citriperazine B |  | C13H16N2O2S2 296 |
| Prenyl chain modified analogues | |||
| 21 | cis-3-(4-(4-Hydroxy-3-methyl-2-butenyl)oxy)benzyl)-1,4-dimethyl-3,6-bis(methylthio)piperazine-2,5-dione; bis-(methylthio)silvatinol |  | C20H28N2O4S2 424 |
| 22 | Bilain A |  | C20H26N2O5S2 438 |
| 23 | Bilain B |  | C20H30N2O5S2 442 |
| 24 | Bilain C |  | C23H31N3O6S2 509 |
| 25 | Bilain D |  | C20H30N2O5S2 442 |
| 26 | Bilain E |  | C21H32N2O5S2 456 |
| 27 | Bilain F |  | C21H32N2O5S2 456 |
| Species (Strain) | Source | Geographic Origin | Compound Code | Ref. |
|---|---|---|---|---|
| Cordyceps javanicus1 (961331) | Jaspis cf. coriacea (sponge) | Fiji | 1, 3 | [17] |
| Fusarium chlamydosporum (OUPS-N124) | Carpopeltis affinis (red alga) | Japan | 1, 4, 5, 11, 16, 18 | [8] |
| Nigrospora sp. (PSU-F12) | Annella sp. (gorgonian) | Similan Islands (Thailand) | 5 | [18] |
| Penicillium bilaiae (MST-MF667) | Boat ramp | Huon estuary, Tasmania (Australia) | 1, 22, 23, 24 | [19] |
| Penicillium commune (518) | Muricella abnormalis (gorgonian) | Danzhou, Hainan (China) | 1 | [20] |
| Penicillium crustosum (HDN153086) | Sediment | Prydz Bay (Antarctica) | 1, 3, 15 | [21] |
| Penicillium sp. (KMM 4672) | Padina sp. (brown alga) | Vietnam | 19, 20 | [22] |
| Penicillium sp. (2556) | Mangrove plant | China | 4, 5 | [23] |
| Penicillium waksmanii (OUPS-N133) | Sargassum ringgoldianum (brown alga) | Japan | 1, 16, 21 | [9] |
| Sarocladium kiliense (HDN11-84) | Rhizosphere soil of Thespesia populnea (mangrove) | Guangxi (China) | 1, 6, 7, 12 | [24] |
| Trichoderma virens (Y13-3) | Gracilaria vermiculophylla (red alga) | Yangma Island (China) | 16, 17 | [6] |
| Species (Strain) | Source | Geographic Origin | Compound Code | Ref. |
|---|---|---|---|---|
| Aspergillus silvaticus (IFO8173) | Soil | Tafo (Ghana) | 8, 2 | [4] |
| Coriolus (=Irpex) consors (ATCC11574) | ATCC collection | 1, 3 | [25] | |
| Penicillium amphipolaria (DAOM695760) | Soil | Quartermain Mountains (Antarctica) | 14 | [26] |
| Penicillium brevicompactum | Contaminant in culture of Ceratocystis ulmi (plant pathogenic fungus) | Edmonton (Canada) | 1, 9, 10, 16 | [10] |
| Penicillium crustosum (VR4) | Viguiera robusta (plant) | Brazil | 1, 3, 14 | [27] |
| Penicillium crustosum (MK285663) | Fruiting body of Isaria cicadae (entomopathogenic fungus) | Sichuan province (China) | 1, 13, 25, 26, 27 | [7] |
| Penicillium crustosum (YN-HT-15) | Red soil | Yunnan (China) | 12, 13 | [28] |
| Penicillium roqueforti (ATCC10110) | Blue cheese | USA | 1 | [29] |
| Penicillium sp. | Endophytic in Pinellia ternata (plant) | Nanjing (China) | 1 | [30] |
| Tolypocladium sp. | Quercus virginiana (plant) | Tamalupas (Mexico) | 1, 4, 5, 9 | [5] |
| Trichoderma virens1 (CMI101525) | Soil | California, USA | 1, 16 | [3,11] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salvatore, M.M.; Nicoletti, R.; DellaGreca, M.; Andolfi, A. Occurrence and Properties of Thiosilvatins. Mar. Drugs 2019, 17, 664. https://doi.org/10.3390/md17120664
Salvatore MM, Nicoletti R, DellaGreca M, Andolfi A. Occurrence and Properties of Thiosilvatins. Marine Drugs. 2019; 17(12):664. https://doi.org/10.3390/md17120664
Chicago/Turabian StyleSalvatore, Maria Michela, Rosario Nicoletti, Marina DellaGreca, and Anna Andolfi. 2019. "Occurrence and Properties of Thiosilvatins" Marine Drugs 17, no. 12: 664. https://doi.org/10.3390/md17120664
APA StyleSalvatore, M. M., Nicoletti, R., DellaGreca, M., & Andolfi, A. (2019). Occurrence and Properties of Thiosilvatins. Marine Drugs, 17(12), 664. https://doi.org/10.3390/md17120664