1. Introduction
The
Fucus vesiculosus Linnaeus (1753), popularly known as bladderwrack, is an edible brown macroalga (Ochrophyta, Phaeophyceae) that is commonly present on the middle shores of the western Baltic Sea, Atlantic coasts of North America, Europe, and Western Mediterranean [
1,
2]. This macroalga has been used for food and feed in Europe since the 17th century, as well as in traditional medicine in China [
3]. It is currently used as a raw material for various industries, targeting food processing, the development of new pharmaceuticals and cosmetics, along with other new products to high-end markets [
4,
5,
6,
7]. The culture of this macroalga is attracting a growing interest in Europe, mainly due to its effect on reducing the environmental impact of intensive agriculture and fish aquaculture and eutrophication [
8,
9,
10]. The production of
Fucus vesiculosus using land-based integrated multi-trophic aquaculture (IMTA) is considered a sustainable alternative to producing this macroalga [
11].
Fucus vesiculosus has already been reported to be an important source of iodine [
12], polysaccharides, e.g., fucoidans [
13,
14] and alginic acid, phlorotannins [
1], and also high-valued molecules such as lipids (1–5% per dry biomass) [
15,
16,
17,
18]. The lipid fraction of
F. vesiculosus is characterized by a high amount of essential polyunsaturated fatty acids (PUFA), such as linoleic (18:2
n-6), α-linolenic (18:3
n-3) and octadecatetraenoic (18:4
n-3) acids, as well arachidonic (20:4
n-6) and eicosapentaenoic (20:5
n-3) fatty acids, which grant this brown macroalgae high nutritional value [
15,
16,
17]. Polyunsaturated fatty acids are important in the prevention of several diseases, such as cardiovascular diseases and cancer [
19,
20], and the consumption of this brown macroalgae is often identified as a healthier choice option.
In macroalgae, fatty acids are mostly esterified to more complex lipids such as glycoglycerolipids (GLs), glycerophospholipids (PLs), and betaine lipids. These polar lipids are structural components of cell membranes and organelles, namely chloroplasts [
21], and act as sources of PUFA but also have important health benefits. Polar lipids from brown macroalgae have been reported to have anti-inflammatory and antitumoral activities [
4,
22,
23,
24,
25]. More recently, some polar lipid classes such as betaine lipids, phosphatidylethanolamine, and phosphatidylcholine phospholipids were identified in the organic solvent extracts of
F. vesiculosus, that showed an antimicrobial effect against the ESKAPE panel of bacterial, fungal and algal pathogens [
26]. These organic solvent extracts also showed a significant inhibitory effect on the pancreatic cancer cell line Panc1, with higher induction effect observed in extracts of samples collected in the winter and end of spring seasons [
26]. The knowledge on the polar lipid composition of
Fucus is an essential step for the discovery of lipidic bioactive compounds and new bioactive properties, fostering the bioprospection of lipidic extracts.
The lipid metabolism of macroalgae is modulated in response to changes in environmental factors such as temperature and light [
27,
28,
29,
30,
31,
32,
33,
34]. The variation of lipids with season was evaluated in the brown alga
Fucus serratus. It was reported that low light irradiance and a shorter light period can induce an accumulation of glycolipids monogalactosyldiacylglycerol (MGDG), digalactosyldiacylglycerol (DGDG) and sulfoquinovosyldiacylglycerol (SQDG), and phospholipids phosphatidylglycerol (PG), phosphatidylcholine (PC) and phosphatidylethanolamine (PE), both present in photosynthetic membranes [
33]. Concerning the effect of temperature, it was reported for
Fucus sp. that cold water temperature can induce an increase of degree of fatty acid unsaturation [
33,
35,
36].
So far, the lipidome from
F. vesiculosus and the seasonal effect on its profile was mainly studied using the FA composition in total lipid extracts, and after separation of lipid class by TLC [
16] but never has it been addressed at the molecular level of each molecular species. The identification of the lipid composition at the molecular level can be effectively achieved by using a liquid chromatography-mass spectrometry (LC–MS)-based lipidomic approach. This approach has been successfully used to characterize the lipidome profile of several macroalgae such as
Chondrus crispus (Florideophyceae) [
37],
Codium tomentosum (Bryopsidophyceae) [
38],
Gracilaria sp. (Florideophyceae) [
39],
Porphyra dioica (Bangiophyceae) [
40],
Ulva sp. (Ulvophyceae) [
41], fostering the valorization and the identification of potential valuable lipids in macroalgae.
In this study, we identified the lipidome of F. vesiculosus by using hydrophilic interaction chromatography (HILIC)–LC coupled with high-resolution mass spectrometry (HRMS) and evaluated the effects of seasonal variation, e.g., to winter versus spring seasons, on the lipidome. Fucus vesiculosus was collected on a land-based integrated aquaculture framework at two different sampling occasions: February (winter) and May (spring) collections. The result gathered in the present work will contribute to enhancing the knowledge on the nutritional value of this alga as food and for the bioprospection of target active ingredients for application in pharmaceutical, cosmetical and cosmeceutical industry.
3. Discussion
In this work, the profile in the polar lipid of
F. vesiculosus was identified for the first time using a modern high-resolution LC–MS platform. We have identified in the lipidome of
F. vesiculosus eighteen different classes of polar lipids such as the glycolipids MGDG, MGMG, DGDG, DGMG, SQDG, and SQMG, the phospholipids PC, LPC, PE, LPE, PG, LPG, PI, and LPI and the betaine lipids DGTA, MGTA, DGTS; and MGTS, in a total of 187 molecular species. It is well known that the lipid composition and dynamics of the algae lipidome is modulated by different environmental conditions, such as temperature and light, as usually experienced in different seasons or geographical origins [
33,
44]. Herein, we evaluated the plasticity of the lipidome of
Fucus in two seasons, February (winter) and May (spring). In that year, winter´s batch temperature and light expositions were 13.2 ± 1.17 °C and 177 µmol photons m
2 s
−1 (average irradiance), and in the spring were 17.4 ± 1.36 °C and 424 µmol photons m
2 s
−1, respectively.
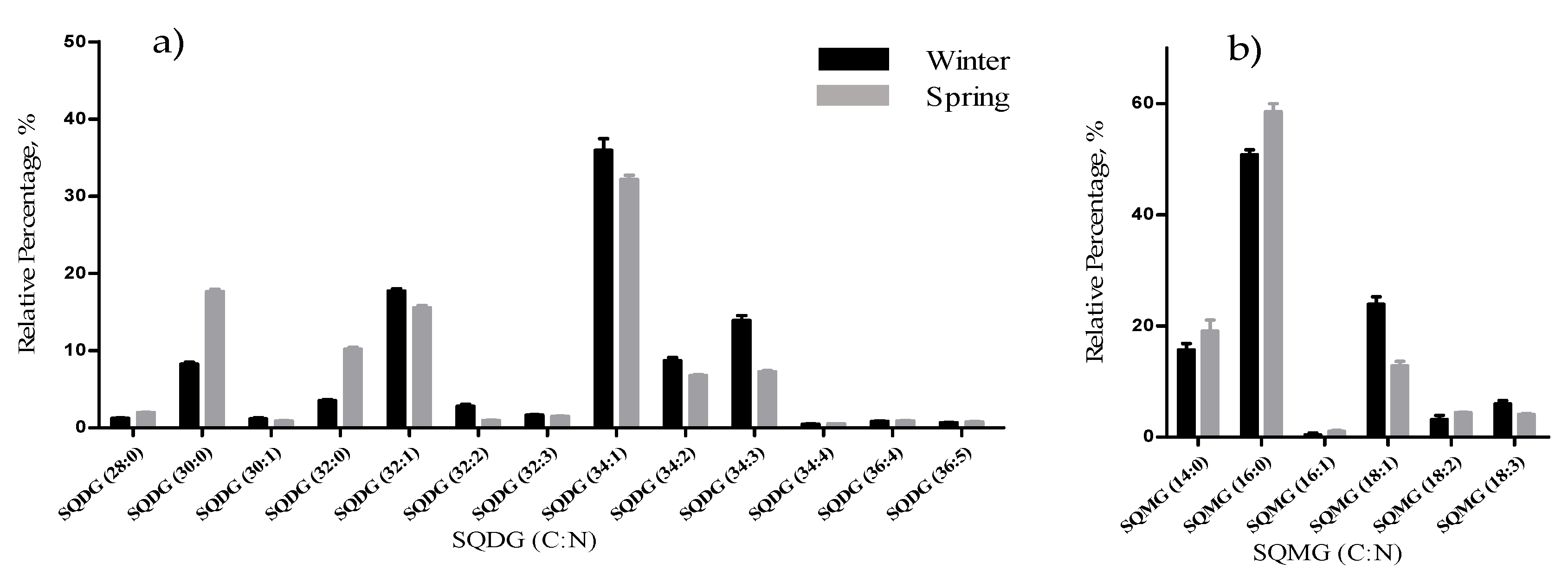
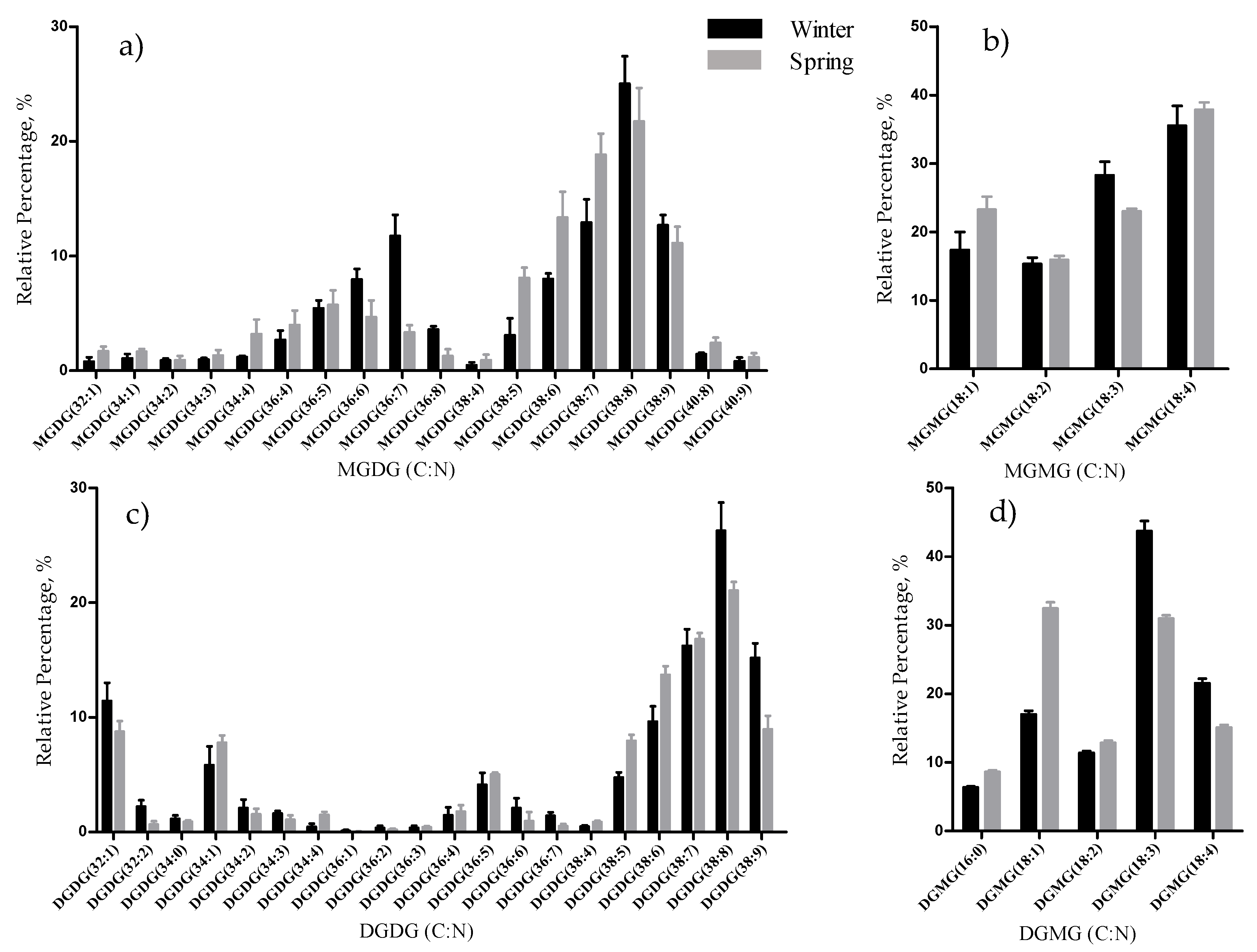
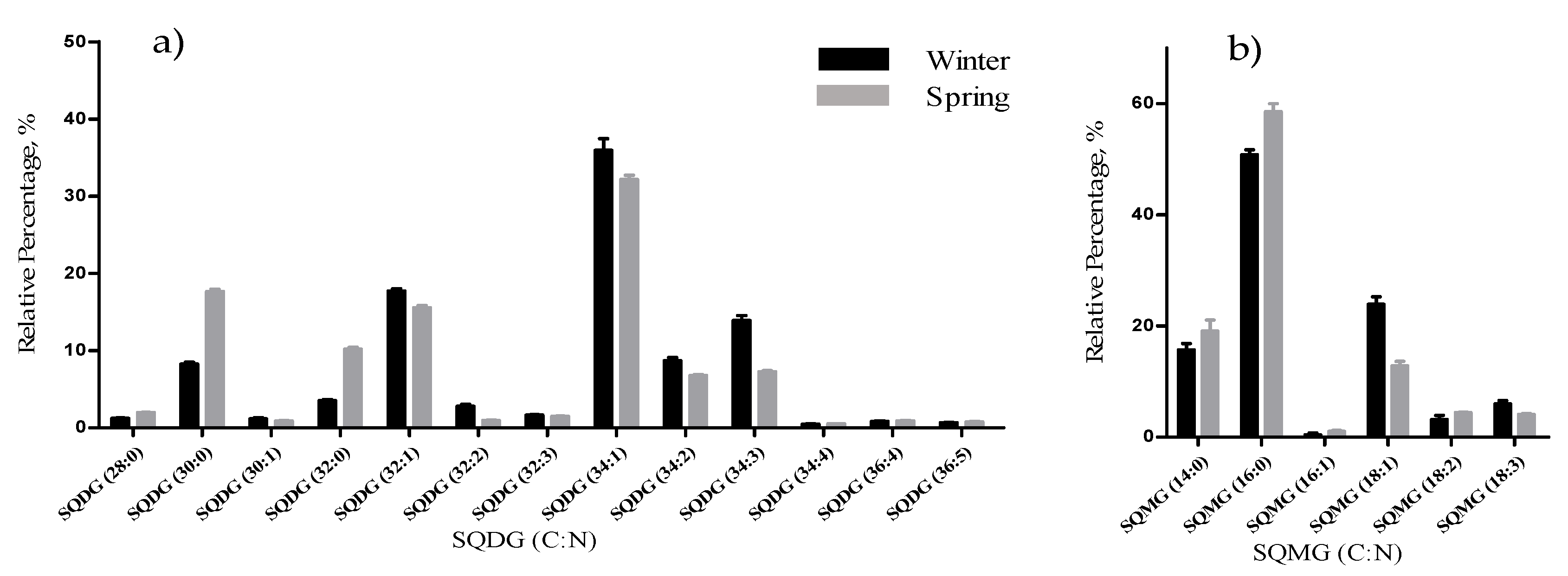
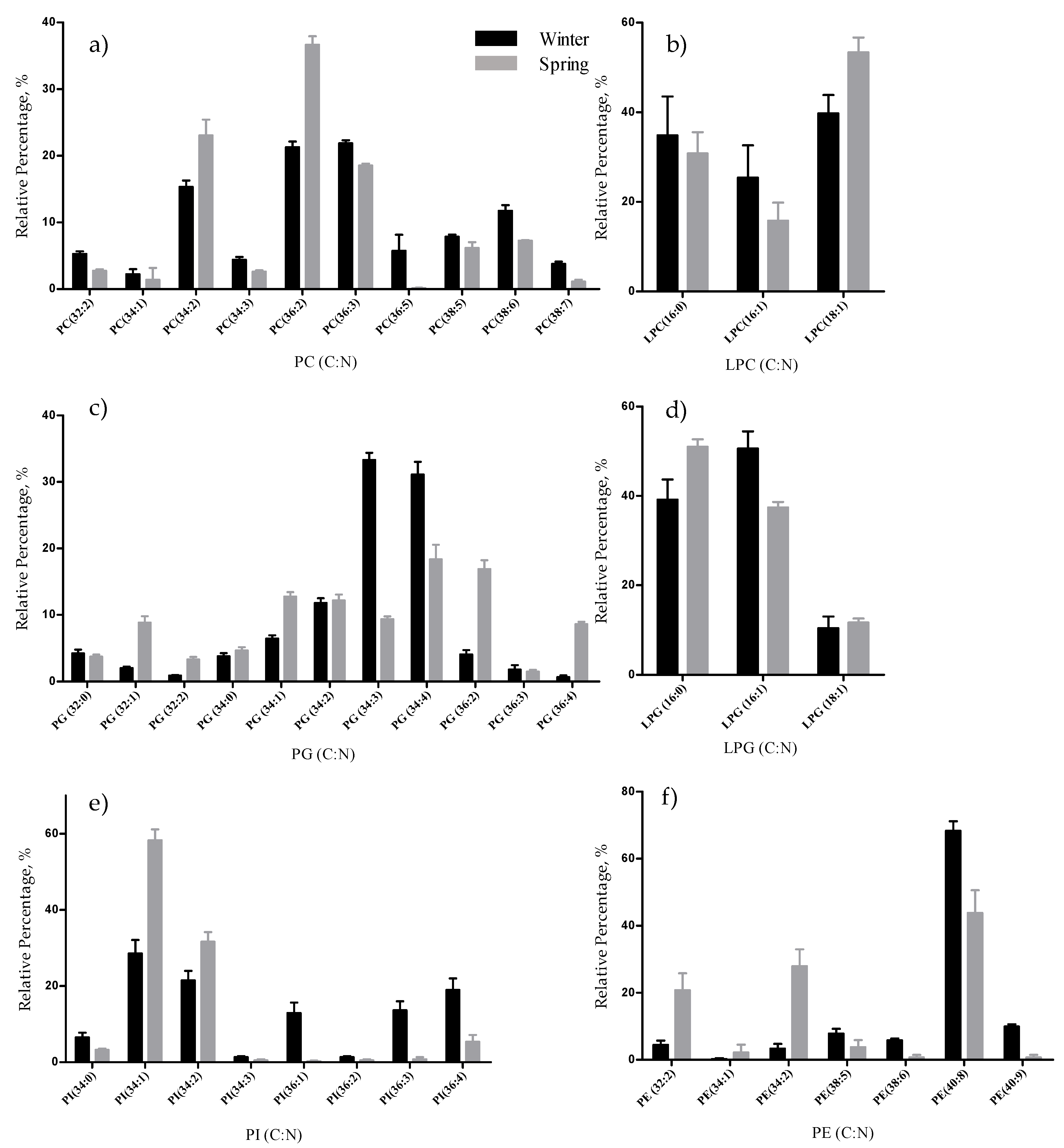
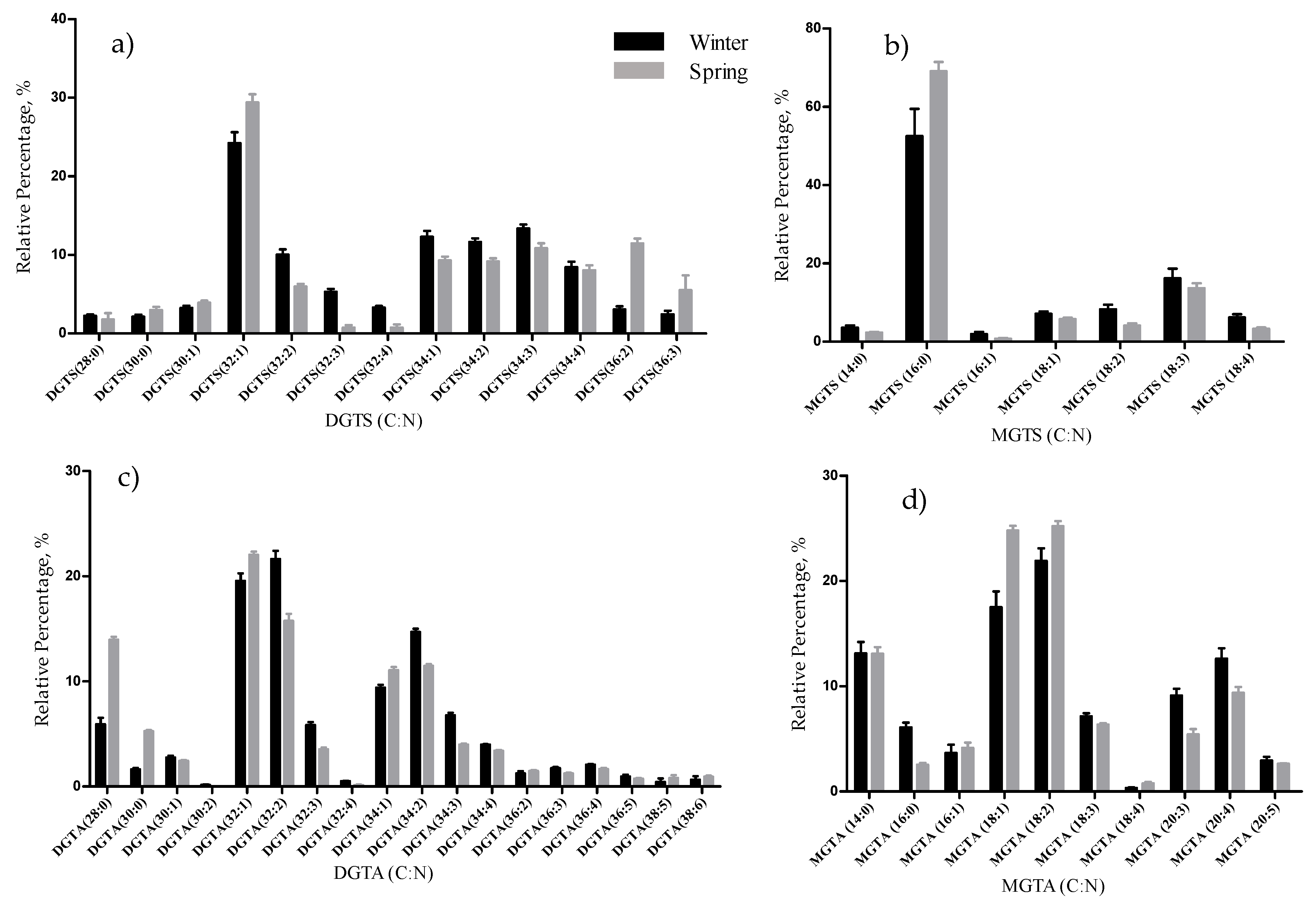
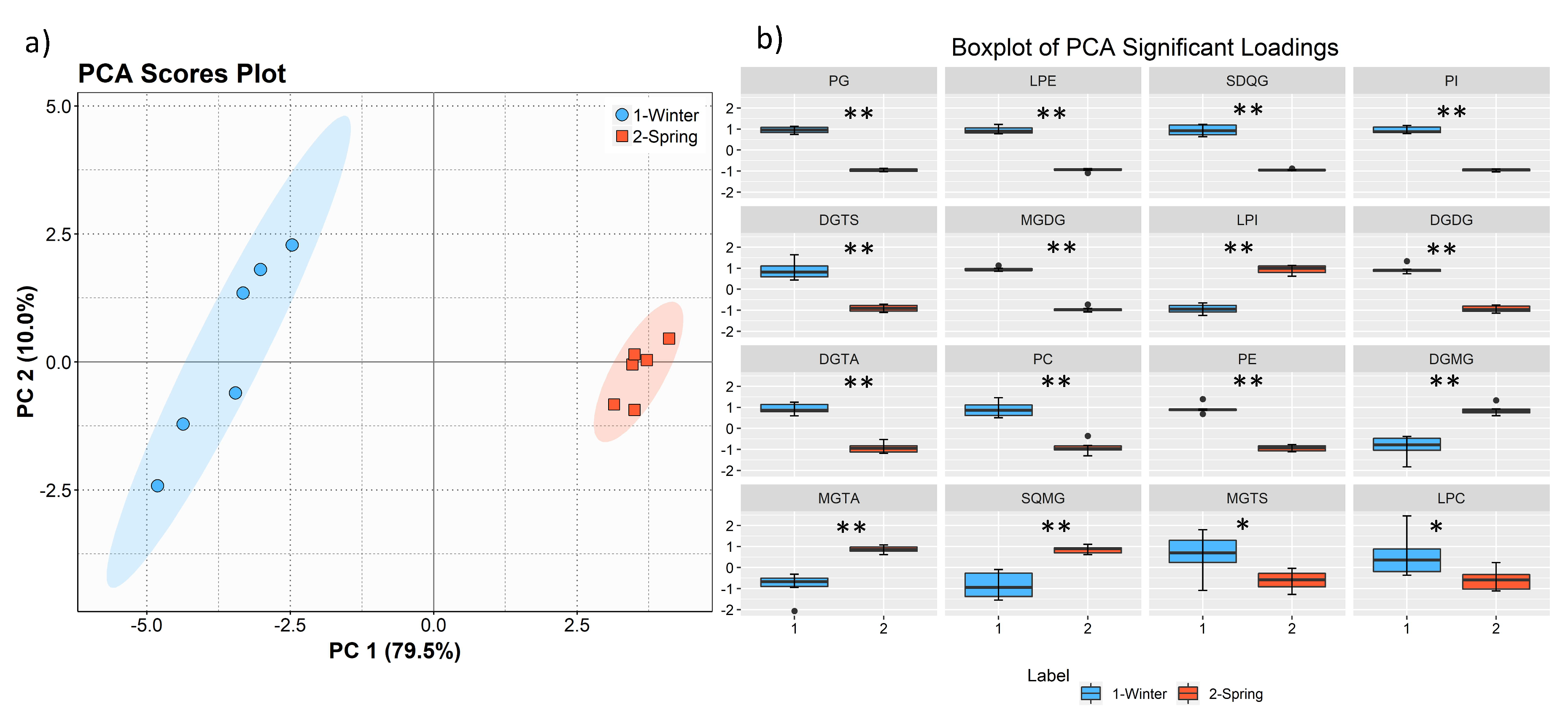
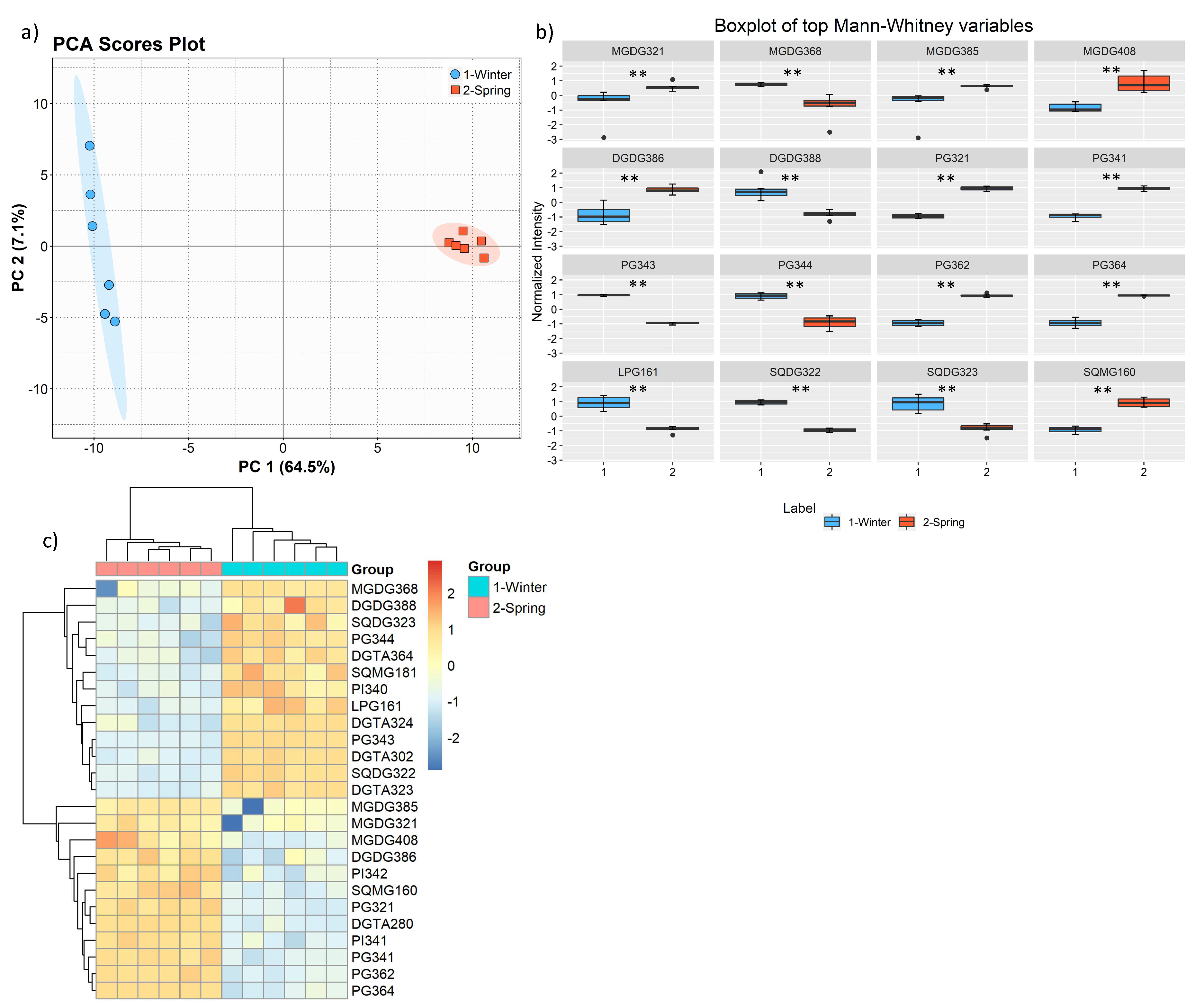
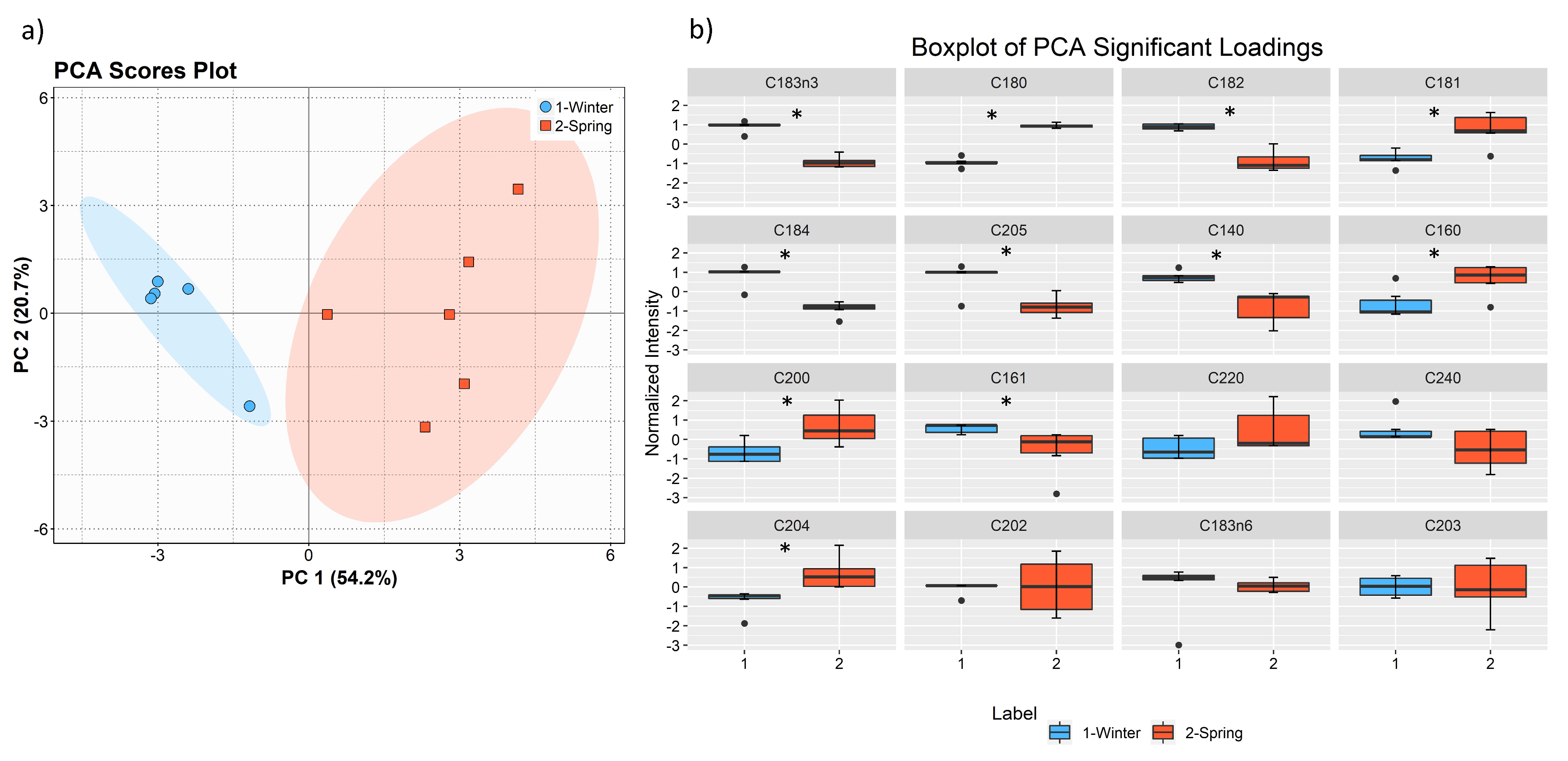
The total lipid content was significantly higher in the winter than in the spring (1.74% versus 1.47% DW), although we have identified the same polar lipids species in samples from both seasons. We observed significant differences in the relative abundances of several polar lipids classes and molecular species (
Figure 1,
Figure 2,
Figure 3,
Figure 4,
Figure 5,
Figure 6 and
Figure 7,
Table 5), but also in several fatty acids from the total lipid extracts (
Figure 8). There was an increase of the relative abundance (RA) of lyso-glycolipids and lyso-betaine lipids in May, while diacyl polar lipids had higher RA in the winter samples. Also, samples from the winter season were more abundant in glycolipids and phospholipids containing highly unsaturated FA and in betaine lipids combining short chain acid saturated FA and PUFA. Similarly, the glycolipids of samples from winter had higher RA of species containing long-chain FA with degrees of unsaturation from 3 to 5 double bonds, when compared with samples from spring. This tendency was previously reported for
F. serratus [
33], that in the winter had increased total percentage of PUFA and polar lipids rich in PUFA (MGDG, PG, and DGDG). Such seasonal changes in the polar lipid composition were associated by the authors with temporal changes in winter (low light intensity, low irradiance, and shorter day length). Under low light intensity, the RA of the glycolipids from chloroplast membranes (MGDG, DGDG, SQDG) increased most probably to adaptation of the thylakoids membranes and the maintenance of photosynthetic processes [
45,
46,
47,
48]. The ability of algae to adjust the ratio MGDG/DGDG composition has been suggested to be a strategy in coping with the low temperature [
33,
49], and the combined effect of the increased level of unsaturations of glycolipids is considered crucial for the stabilization of the photosynthetic apparatus and of proteins in the membrane. Also, the increase of the RA of the SQDG class could be related to its protective effect on the photosynthetic protein complexes [
33]. The plasticity of phospholipids to weather seasons also followed this trend in the acclimatization to the winter season, characterized by higher RA of unsaturated FA, mainly due to the contribution of PG. PG has an important role associated with the photosynthetic membrane preservation, which can be comparable to SQDG [
33].
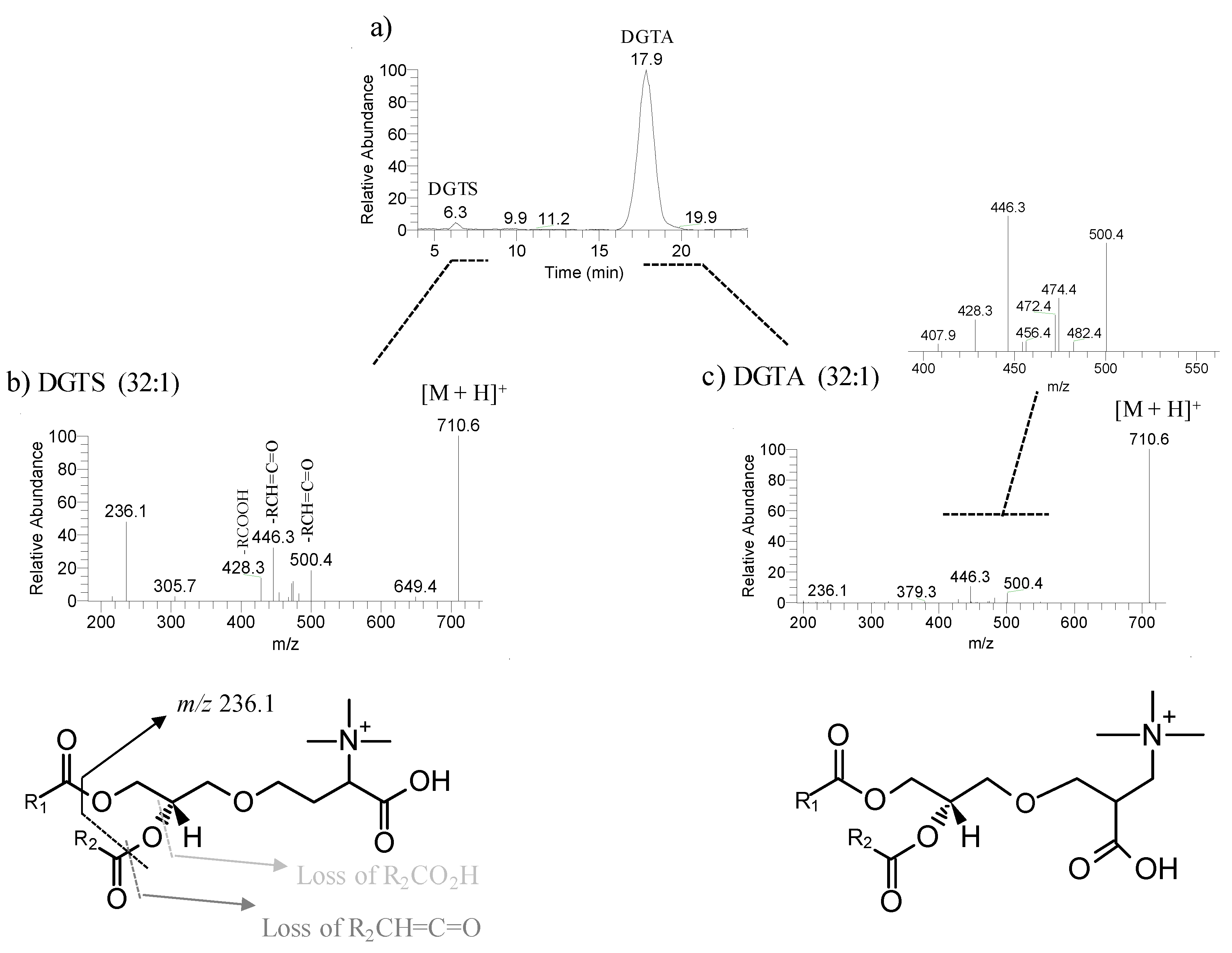
In this study, DGTA were the most abundant betaine lipids, as in previous reports that considered this betaine class a taxonomic marker of brown macroalgae [
50,
51]. DGTA are found exclusively in the brown macroalgae while DGTS are typically present in the green and red macroalgae. The plasticity and adaptation to environmental change of betaine lipids were evidenced by the increase of molecular species with a longer chain length and unsaturated FA in the winter samples. The role of betaine lipids in response to seasonal changes has not been reported yet. Nevertheless, in the summer, DGTA was found to be overexpressed in
F. vesiculosus, with higher RA of saturated molecular species such as DGTA 28:0 and 30:0 [
26]. However, the authors did not find a relation between unsaturation and the seasonality pattern, as we describe it in this study.
The trend we have observed for the increase of FA unsaturation in the winter season was corroborated either by the polar lipidomics data and the total FA data and further evidenced by using the Double bond index (DBI) analysis. The adaptation of the lipid metabolism in brown macroalgae to lower temperatures during winter, by modulating the biosynthesis of PUFA and increasing lipid desaturation, was already reported in studies that evaluated the fatty acid profile of
F. vesiculosus [
33,
52,
53]. A similar effect was observed in the brown macroalgae
Undaria pinnafida [
29],
Sargassum pallidium [
28],
Egregia menziesii [
32], and
Costaria costata [
54] that showed higher content of PUFA and higher
n-3/
n-6 ratio in response to cold temperatures. The proportion of saturated and unsaturated acyl chains in membrane lipids is a critical factor that affects lipid packing, membrane viscosity, and water permeability. In Poikilothermic organisms, such as algae, that do not control their body temperature, the increase the proportion of unsaturated acyl chains in membrane lipids is essential to maintain fluidity at lower temperatures and to hold photosynthesis [
49].
Herein, we have also profiled the total fatty acids, including those esterified to polar lipid in
F. vesiculosus, and find that included saturated, monounsaturated and polyunsaturated FA, with a predominance of omega 3 PUFA 18:3, 18:4 and 20:5. As observed in the polar lipid composition determined by LC–MS/MS, omega 6 PUFA, namely 18:2 and 20:4, were also abundant. The calculated ratio of
n-6/
n-3 FA was 1.20 in winter and 1.95 in spring, with an average of 1.57, which is a lower ratio than the maximum recommended by the World Health Organization (lower than 10) [
55,
56]. As such,
F. vesiculosus can be considered a sustainable source of PUFA [
28,
29,
51,
57], either available by the consumption as edible seaweed, or used as a raw material in food industries and biotechnological applications [
4]. Due to its content in
n-6 and
n-3 PUFA, the consumption of this seaweed could have benefits for preventing cardiovascular disorders, depression and other mental disorders [
56]. Despite the observed variations in the RA of polar lipids and total FA, the molecular species bearing PUFA were still the most abundant ones in both seasons, and thus both batches have a remarkable composition to be used for food or feed diet as a source of lipids with nutritional value.
Phospholipids can also be explored as add value ingredients. They are considered appealing food ingredients, for fortification of the content of foods in
n-3 FAs such as 20:5 (
n-3) and to enhance its nutritional values [
58]. There are also recognized health benefits in the consumption of PL bearing to
n-6 and
n-3 PUFA, contributing to maintain normal physiological functions and to prevent inflammatory diseases, among others. This foster the growing interests in the use of PLs from macroalgae in the nutraceutical industries [
58,
59,
60]. Polar lipids are also claimed for cosmetics and pharmaceutical applications, namely phospholipids used as carriers liposomes, lipid emulsions, micelles, drug-phospholipids complexes and cochleates [
61,
62].
The polar lipidome of
F. vesiculosus also included some species that were reported as bioactive phytochemicals. The glycolipids MGDG (20:5/18:3) and MGDG (20:5/18:4), that in this study were abundant species in both seasons, were previously isolated from
Fucus spiralis [
24]. These species showed anti-inflammatory activity, as they reduced the NO release by activated macrophages in a dose-dependent manner [
24]. Furthermore, MGDG (18:4/16:0), DGDG(18:4/16:0), DGDG (20:4/16:0), DGDG (20:5/16:0) and SQDG (20:5/14:0), also identified in this study, were associated with anti-inflammatory activity through down-regulation of iNOS [
63]. Also, the MGDG (20:5/18:4) was found to inhibit the growth of human melanoma cells [
25]. Antimicrobial activity was associated with MGDG (20:5/18:4) and MGDG (18:3/18:4) molecular species [
64] and also SQDG molecular species [
22]. Sulfolipids, such as SQDG or SQMG, which in this study increased in the spring samples, were associated with anti-HSV1 and anti-HSV2 activities [
25] while SQMG isolated from the extract of the brown macroalga was active against
Xanthomonas oryzae pv. Oryzae and associated with antibacterial activity [
65]. Glycolipids can also be ingredients in cosmetic formulations and as biosurfactants, highly considered to replace synthetic surfactants [
66].
The composition in lipid herein assigned to F. vesiculosus supports its use as a functional food or as raw biomass, source of polar lipids, to be used as food ingredients, or even as a source of bioactive lipids. Our results on the lipidomics profile of F. vesiculosus in two different seasons contribute to a better understanding of the lipidome plasticity and adaptation to environmental changes and reinforces the importance on the knowledge of the lipidome at a molecular level for bioprospection of the potential of brown macroalgae as a source of natural products for different biotechnological applications.
5. Conclusions
This work showed new insights into the identification of the lipidome of F. vesiculosus at a molecular level. A total of 187 molecular species distributed by glycolipids, phospholipids, and betaine lipids were identified. The lyso-betaine MGTA is reported for the first time in macroalgae. The lipidome plasticity and adaptation to environmental changes with the season (winter and spring) of F. vesiculosus was evidenced by the variation in the content of lipids bearing MUFA and PUFA. Multivariate statistical analysis, PCA and cluster analysis, showed a clear clustering according to lipid diversity and seasonal groups. In winter (February), the effects of lower temperature and lower light exposition period lead to the increase of polyunsaturated esterified FA majorly of glycolipids, some phospholipid and DGTA betaine lipid classes.
F. vesiculosus lipidome contained eighteen classes of polar lipids that were the main source of the 18:3(n-3), 18:4(n-3), 20:4(n-6), and 20:5(n-3) PUFA, with high nutritional value. The low n-6/n-3 ratio renders F. vesiculosus suitable for human consumption and feed, independent of the seasonal variation (February and May collections). Also, PUFA bearing polar lipid species that were reported with important bioactive properties were identified in the lipdome of Fucus, with relevance for the anti-inflammatory, antitumoral and antimicrobial properties of some lipid species, fostering the use of F. vesiculosus as a suitable raw material of active ingredients. Noteworthy, the glycolipids species more abundant, mainly in the winter lipidome, were reported to have anti-inflammatory and antitumoral properties. Whereas, sulfolipids SQDG and SQMG species, more abundant in the spring, were associated with antimicrobial activity.

 ,
, 









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}