The Nutritional and Pharmacological Potential of New Australian Thraustochytrids Isolated from Mangrove Sediments

 , ,
, ,  ,
,  and
and 
Abstract
:1. Introduction
2. Results and Discussions
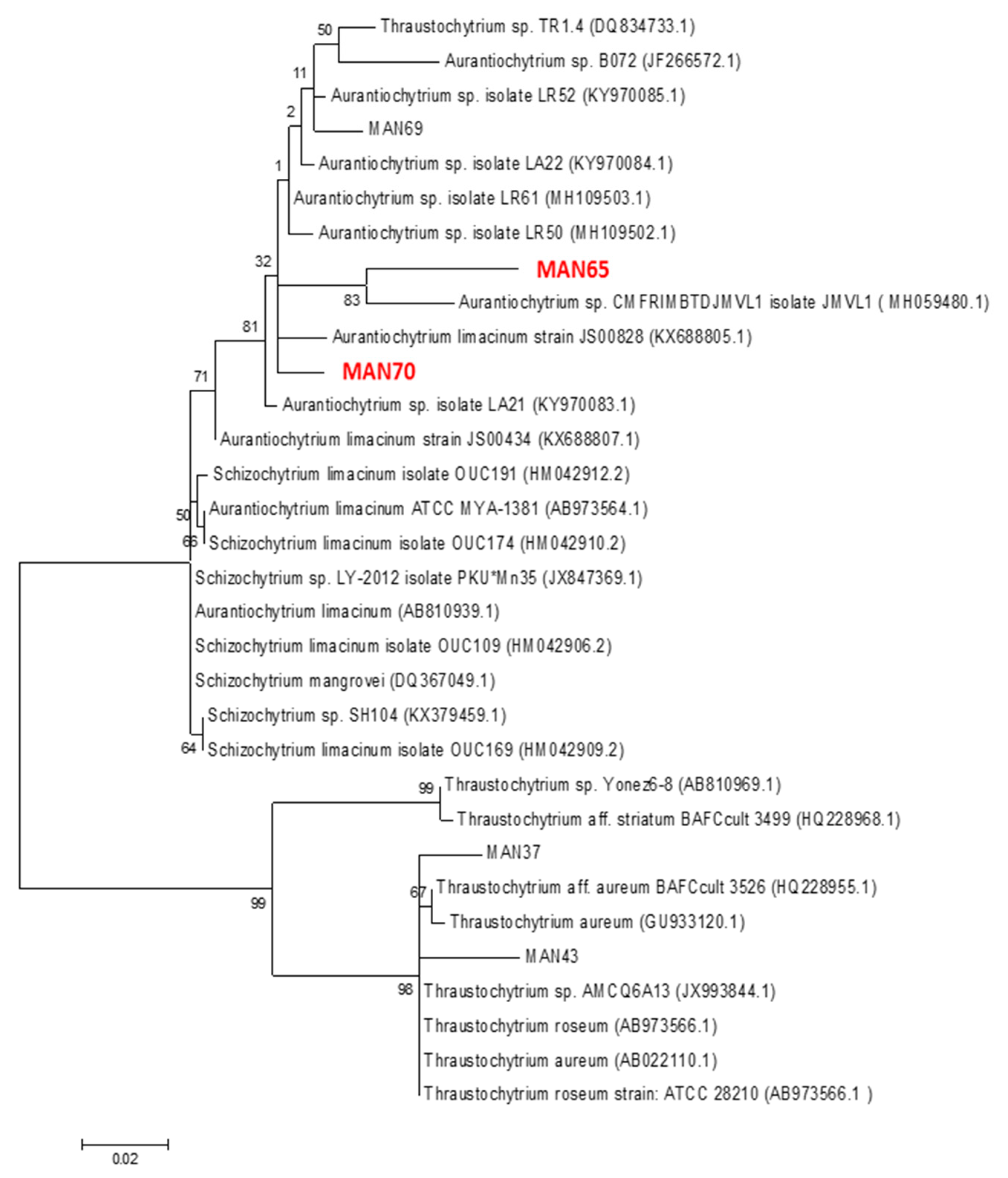
2.1. Phylogenetic Analysis of Thraustochytrids
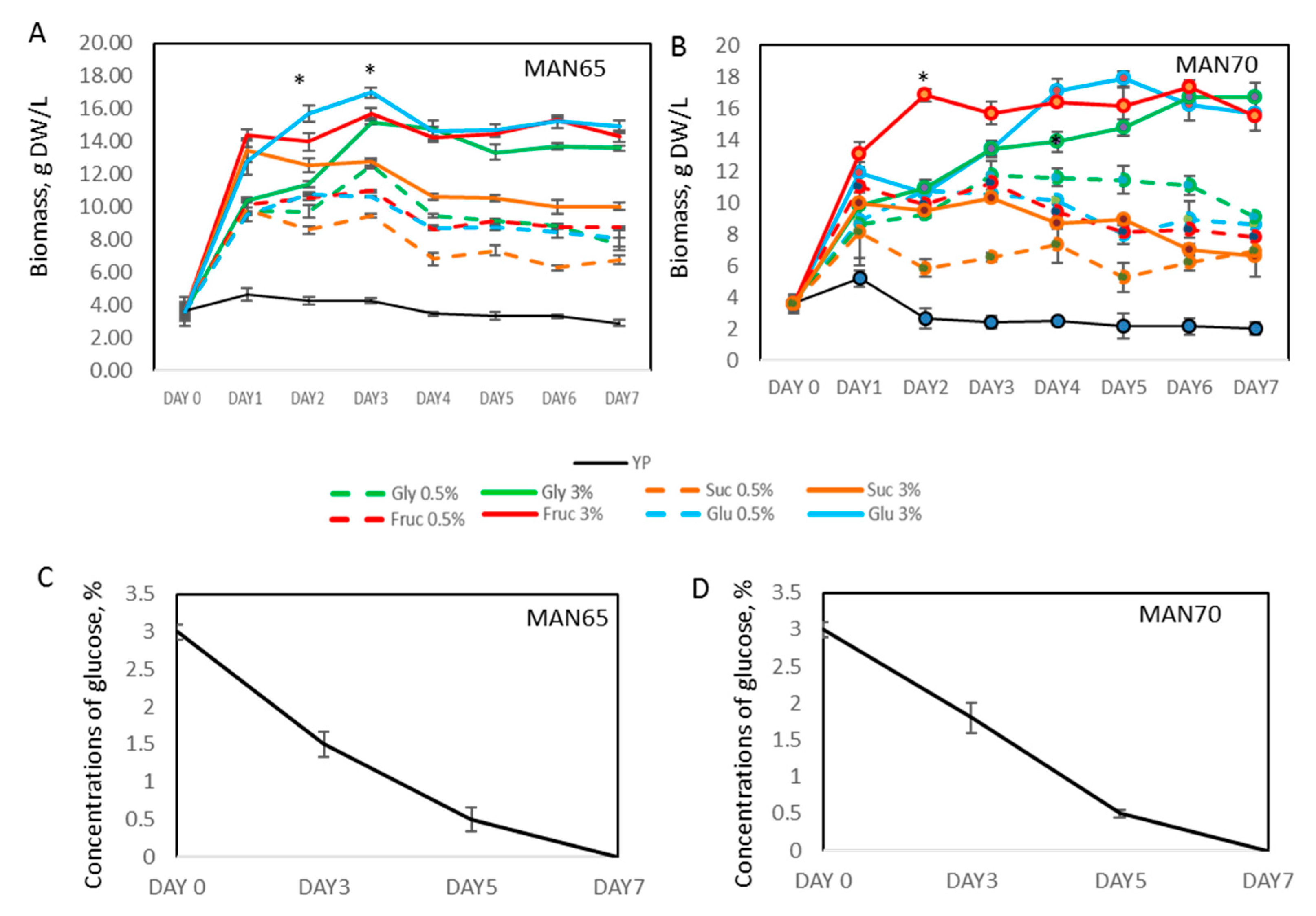
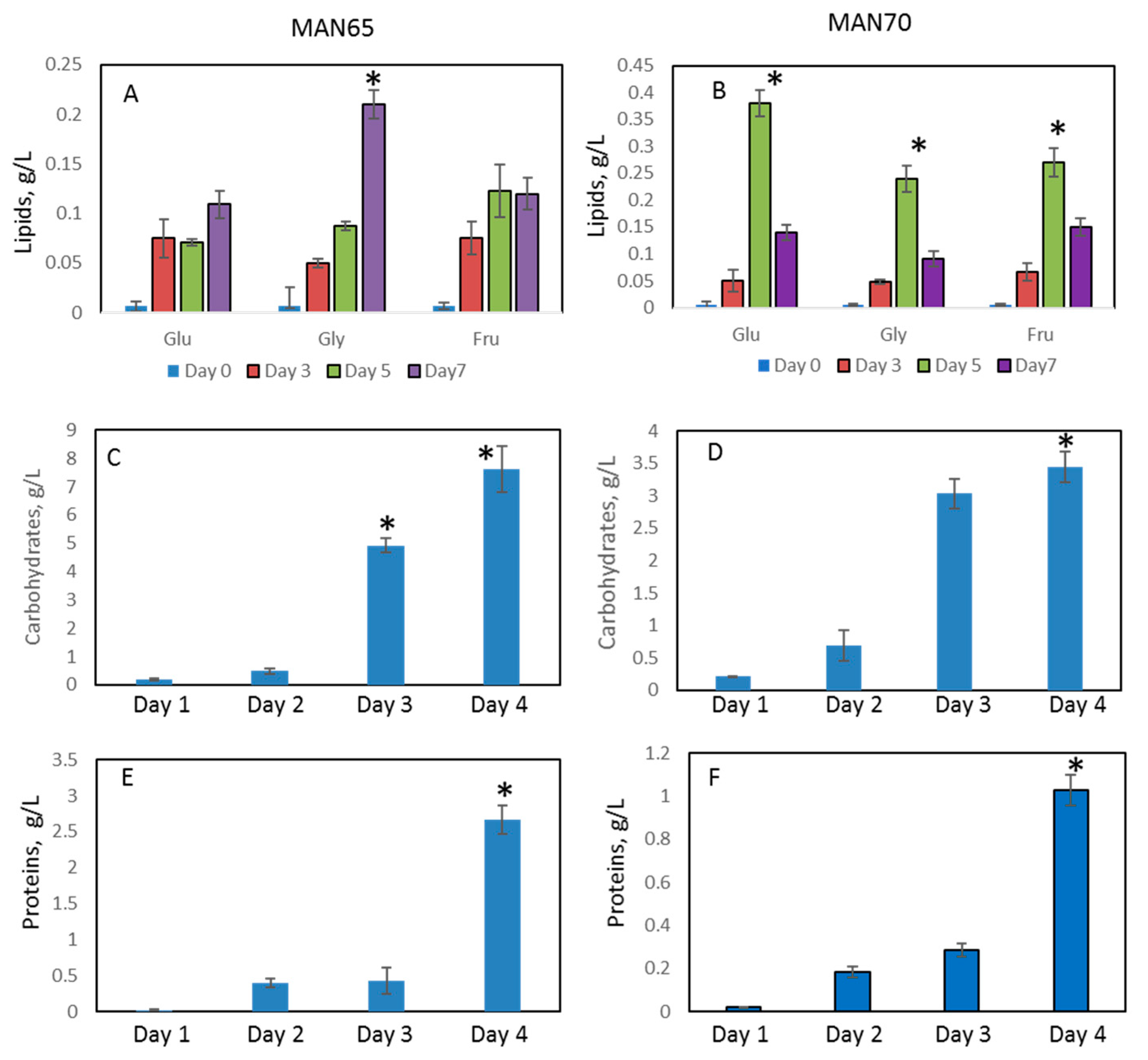
2.2. Biomass Production
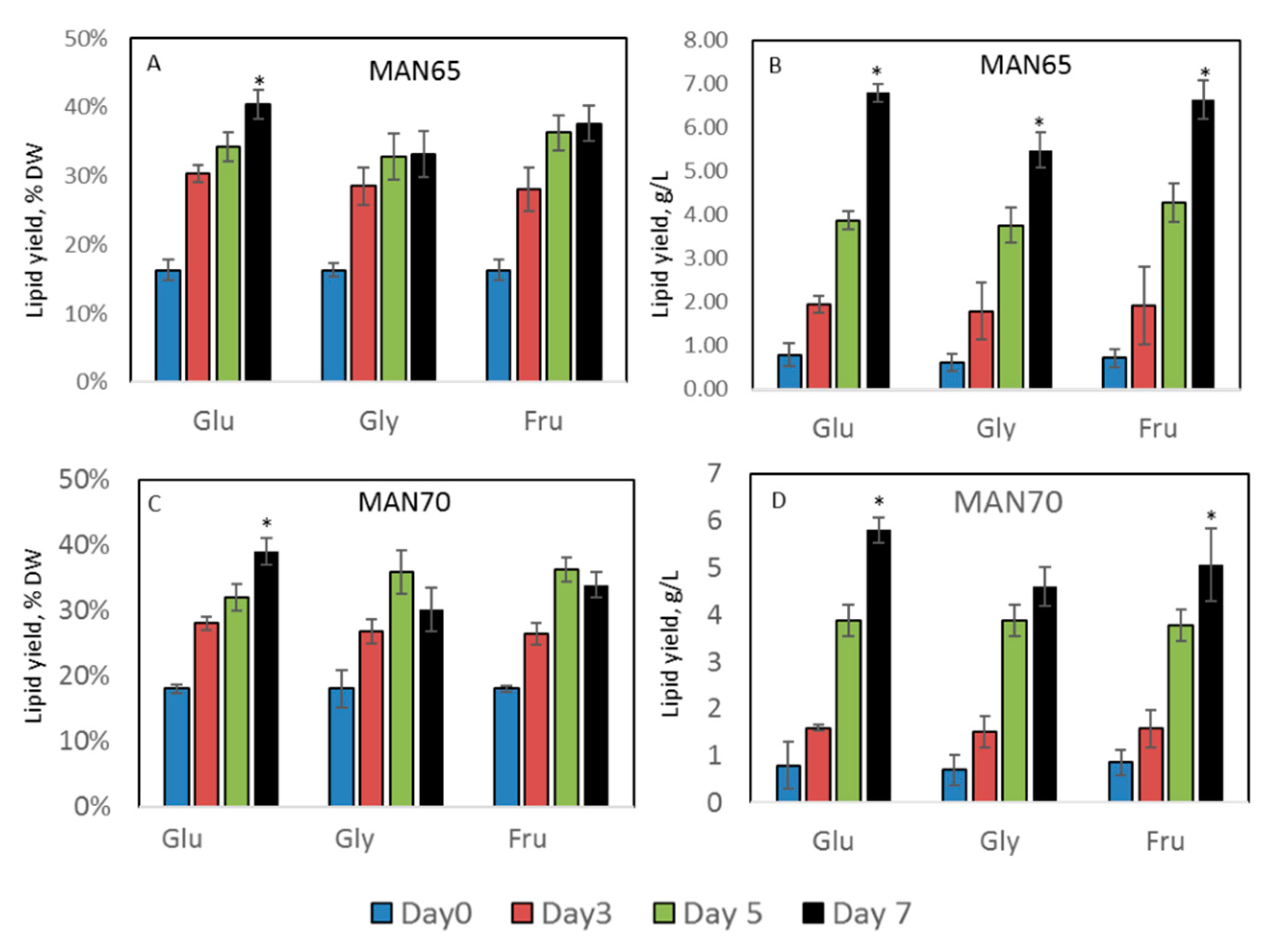
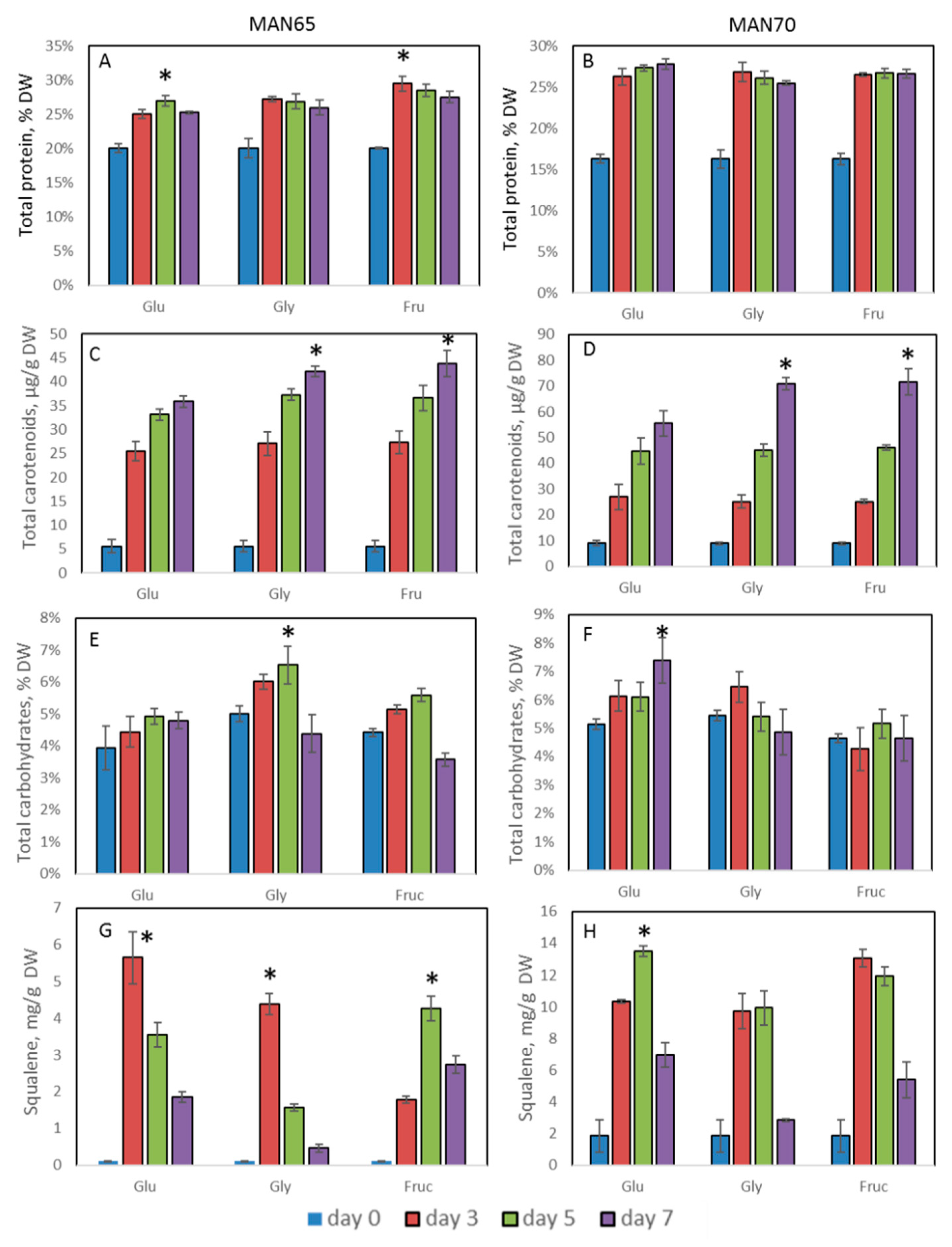
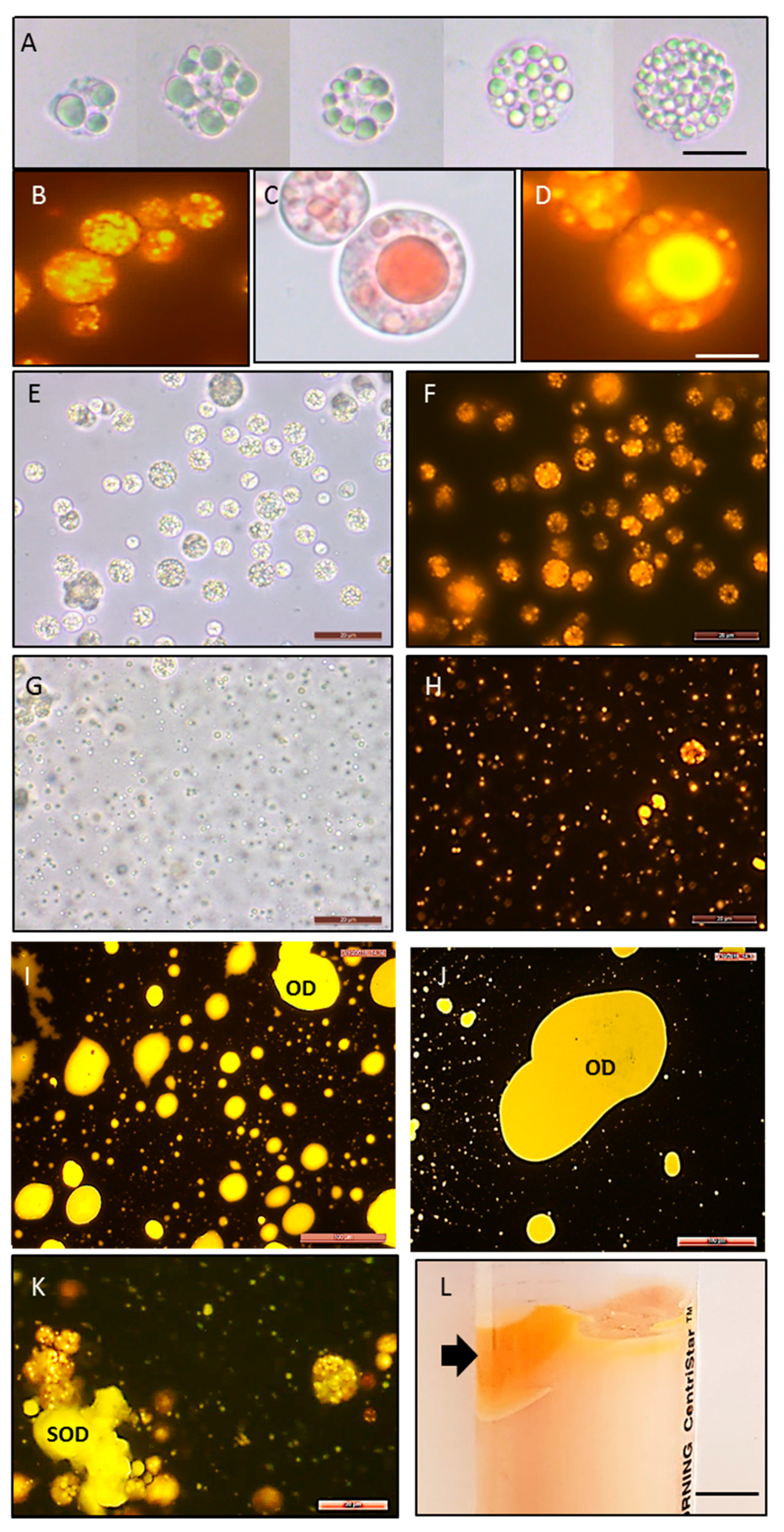
2.3. Intracellular Production of Lipids, Protein, Carbohydrates, Squalene, and Carotenoids
2.4. Extracellular Production of Protein, Lipids, Carbohydrates, EPS, and Recovered Oil Bodies
3. Materials and Methods
3.1. Strain Isolation and Phylogenetic Analysis
3.2. Growth of Thraustochytrids
3.3. Biochemical Analysis
3.3.1. Lipid Extraction and FAMEs Analysis
3.3.2. Carotenoid Extraction and Analysis
3.3.3. Squalene Extraction and Analysis
3.3.4. Total Carbohydrate Analysis
3.3.5. Total Protein Analysis
3.3.6. Microscopy and Nile Red Staining
3.4. Harvesting and Characterization of EPS
3.5. Oil Body Recovery
3.6. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Aasen, I.M.; Ertesvag, H.; Heggeset, T.M.; Liu, B.; Brautaset, T.; Vadstein, O.; Ellingsen, T.E. Thraustochytrids as production organisms for docosahexaenoic acid (DHA), squalene, and carotenoids. Appl. Microbiol. Biotechnol. 2016, 100, 4309–4321. [Google Scholar] [CrossRef] [PubMed]
- Fossier Marchan, L.; Lee Chang, K.J.; Nichols, P.D.; Mitchell, W.J.; Polglase, J.L.; Gutierrez, T. Taxonomy, ecology and biotechnological applications of thraustochytrids: A review. Biotechnol. Adv. 2018, 36, 26–46. [Google Scholar] [CrossRef] [PubMed]
- Morabito, C.; Bournaud, C.; Maes, C.; Schuler, M.; Aiese Cigliano, R.; Dellero, Y.; Marechal, E.; Amato, A.; Rebeille, F. The lipid metabolism in thraustochytrids. Prog. Lipid Res. 2019, 76, 101007. [Google Scholar] [CrossRef] [PubMed]
- Ryan, A.S.; Zeller, S.; Nelson, E.B. 15—Safety Evaluation of Single Cell Oils and the Regulatory Requirements for Use as Food Ingredients. In Single Cell Oils, 2nd ed.; Cohen, Z., Ratledge, C., Eds.; AOCS Press: Urbana, IL, USA, 2010; pp. 317–350. [Google Scholar]
- Lee Chang, K.J.; Nichols, C.M.; Blackburn, S.I.; Dunstan, G.A.; Koutoulis, A.; Nichols, P.D. Comparison of Thraustochytrids Aurantiochytrium sp., Schizochytrium sp., Thraustochytrium sp., and Ulkenia sp. for production of biodiesel, long-chain omega-3 oils, and exopolysaccharide. Mar. Biotechnol. 2014, 16, 396–411. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.; Barrow, C.J.; Puri, M. Omega-3 biotechnology: Thraustochytrids as a novel source of omega-3 oils. Biotechnol. Adv. 2012, 30, 1733–1745. [Google Scholar] [CrossRef]
- Singh, P.; Liu, Y.; Li, L.; Wang, G. Ecological dynamics and biotechnological implications of thraustochytrids from marine habitats. Appl. Microbiol. Biotechnol. 2014, 98, 5789–5805. [Google Scholar] [CrossRef]
- Quilodran, B.; Hinzpeter, I.; Hormazabal, E.; Quiroz, A.; Shene, C. Docosahexaenoic acid (C22:6n-3, DHA) and astaxanthin production by Thraustochytriidae sp AS4-A1 a native strain with high similitude to Ulkenia sp.: Evaluation of liquid residues from food industry as nutrient sources. Enzym. Microb. Technol. 2010, 47, 24–30. [Google Scholar] [CrossRef]
- Burja, A.M.; Radianingtyas, H.; Windust, A.; Barrow, C.J. Isolation and characterization of polyunsaturated fatty acid producing Thraustochytrium species: Screening of strains and optimization of omega-3 production. Appl. Microbiol. Biot. 2006, 72, 1161–1169. [Google Scholar] [CrossRef]
- Gupta, A.; Singh, D.; Barrow, C.J.; Puri, M. Exploring potential use of Australian thraustochytrids for the bioconversion of glycerol to omega-3 and carotenoids production. Biochem. Eng. J. 2013, 78, 11–17. [Google Scholar] [CrossRef]
- Xie, Y.; Sen, B.; Wang, G. Mining terpenoids production and biosynthetic pathway in thraustochytrids. Bioresour. Technol. 2017, 244 Pt 2, 1269–1280. [Google Scholar] [CrossRef]
- Eggersdorfer, M.; Wyss, A. Carotenoids in human nutrition and health. Arch. Biochem. Biophys. 2018, 652, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.R.; Lin, Y.K.; Fang, J.Y. Biological and Pharmacological Activities of Squalene and Related Compounds: Potential Uses in Cosmetic Dermatology. Molecules 2009, 14, 540–554. [Google Scholar] [CrossRef] [PubMed]
- Reddy, L.H.; Couvreur, P. Squalene: A natural triterpene for use in disease management and therapy. Adv. Drug Deliv. Rev. 2009, 61, 1412–1426. [Google Scholar] [CrossRef] [PubMed]
- Naziri, E.; Mantzouridou, F.; Tsimidou, M.Z. Squalene resources and uses point to the potential of biotechnology. Lipid Technol. 2011, 23, 270–273. [Google Scholar] [CrossRef]
- Nakazawa, A.; Kokubun, Y.; Matsuura, H.; Yonezawa, N.; Kose, R.; Yoshida, M.; Tanabe, Y.; Kusuda, E.; Van Thang, D.; Ueda, M.; et al. TLC screening of thraustochytrid strains for squalene production. J. Appl. Phycol. 2014, 26, 29–41. [Google Scholar] [CrossRef]
- Hoang, M.H.; Ha, N.C.; Thom, L.T.; Tam, L.T.; Anh, H.T.L.; Thu, N.T.H.; Hong, D.D. Extraction of squalene as value-added product from the residual biomass of Schizochytrium mangrovei PQ6 during biodiesel producing process. J. Biosci. Bioeng. 2014, 118, 632–639. [Google Scholar] [CrossRef]
- Ren, L.J.; Ji, X.J.; Huang, H.; Qu, L.; Feng, Y.; Tong, Q.Q.; Ouyang, P.K. Development of a stepwise aeration control strategy for efficient docosahexaenoic acid production by Schizochytrium sp. Appl. Microbiol. Biotechnol. 2010, 87, 1649–1656. [Google Scholar] [CrossRef]
- Ren, L.J.; Sun, G.N.; Ji, X.J.; Hu, X.C.; Huang, H. Compositional shift in lipid fractions during lipid accumulation and turnover in Schizochytrium sp. Bioresour. Technol. 2014, 157, 107–113. [Google Scholar] [CrossRef]
- Otagiri, M.; Khalid, A.; Moriya, S.; Osada, H.; Takahashi, S. Novel squalene-producing thraustochytrids found in mangrove water. Biosci. Biotechnol. Biochem. 2017, 81, 2034–2037. [Google Scholar] [CrossRef] [Green Version]
- Marchan, L.F.; Lee Chang, K.J.; Nichols, P.D.; Polglase, J.L.; Mitchell, W.J.; Gutierrez, T. Screening of new British thraustochytrids isolates for docosahexaenoic acid (DHA) production. J. Appl. Phycol. 2017, 29, 2831–2843. [Google Scholar] [CrossRef]
- Gupta, A.; Abraham, R.E.; Barrow, C.J.; Puri, M. Omega-3 fatty acid production from enzyme saccharified hemp hydrolysate using a novel marine thraustochytrid strain. Bioresour. Technol. 2015, 184, 373–378. [Google Scholar] [CrossRef] [PubMed]
- Lee Chang, K.J.; Dumsday, G.; Nichols, P.D.; Dunstan, G.A.; Blackburn, S.I.; Koutoulis, A. High cell density cultivation of a novel Aurantiochytrium sp. strain TC 20 in a fed-batch system using glycerol to produce feedstock for biodiesel and omega-3 oils. Appl. Microbiol. Biotechnol. 2013, 97, 6907–6918. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.; Kim, E.J.; Ryu, B.G.; Park, S.; Choi, Y.E.; Yang, J.W. A novel fed-batch process based on the biology of Aurantiochytrium sp KRS101 for the production of biodiesel and docosahexaenoic acid. Bioresour. Technol. 2013, 135, 269–274. [Google Scholar] [CrossRef] [PubMed]
- Hinzpeter, I.; Quilodran, B.; Stead, R.; Trujillo, L.; Vidal, J.; Shene, C. Isolation of thraustochytrid strains in the coastal zone of Puerto Montt, Chile and evaluation of Docosahexaenoic acid (22:6n-3, DHA) production. Afinidad 2009, 66, 482–487. [Google Scholar]
- Khan, Z.; Bhadouria, P.; Bisen, P.S. Nutritional and therapeutic potential of Spirulina. Curr. Pharm. Biotechnol. 2005, 6, 373–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miranda, A.F.; Liu, Z.; Rochfort, S.; Mouradov, A. Lipid production in aquatic plant Azolla at vegetative and reproductive stages and in response to abiotic stress. Plant Physiol. Biochem. 2018, 124, 117–125. [Google Scholar] [CrossRef]
- Cagnac, O.; Rols, C.; Pagliardini, J.; Calleja, P.; Gady, C.; Vandeplas, S. Protein-Rich Biomass of Thraustochytrids, Culturing Method, and Uses. Patent number WO/2017/012931, 7 July 2015. [Google Scholar]
- Garaiová, M.; Zambojová, V.; Šimová, Z.; Griač, P.; Hapala, I. Squalene epoxidase as a target for manipulation of squalene levels in the yeast Saccharomyces cerevisiae. FEMS Yeast Res. 2014, 14, 310–323. [Google Scholar] [CrossRef] [Green Version]
- Maurya, R.; Paliwal, C.; Ghosh, T.; Pancha, I.; Chokshi, K.; Mitra, M.; Ghosh, A.; Mishra, S. Applications of de-oiled microalgal biomass towards development of sustainable biorefinery. Bioresour. Technol. 2016, 214, 787–796. [Google Scholar] [CrossRef]
- Rashid, N.; Rehman, M.S.U.; Han, J.I. Recycling and reuse of spent microalgal biomass for sustainable biofuels. Biochem. Eng. J. 2013, 75, 101–107. [Google Scholar] [CrossRef]
- Gatrell, S.; Lum, K.; Kim, J.; Lei, X.G. Potential of defatted microalgae from the biofuel industry as an ingredient to replace corn and soybean meal in swine and poultry diets. J. Anim. Sci. 2014, 92, 1306–1314. [Google Scholar] [CrossRef]
- Rothlisberger-Lewis, K.L.; Foster, J.L.; Hons, F.M. Soil carbon and nitrogen dynamics as affected by lipid-extracted algae application. Geoderma 2016, 262, 140–146. [Google Scholar] [CrossRef]
- Maurya, R.; Ghosh, T.; Paliwal, C.; Shrivastav, A.; Chokshi, K.; Pancha, I.; Ghosh, A.; Mishra, S. Biosorption of Methylene Blue by De-Oiled Algal Biomass: Equilibrium, Kinetics and Artificial Neural Network Modelling. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jain, R.; Raghukumar, S.; Tharanathan, R.; Bhosle, N.B. Extracellular polysaccharide production by thraustochytrid protists. Mar. Biotechnol. 2005, 7, 184–192. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Singh, P.; Liang, Y.; Li, J.; Xie, N.; Song, Z.; Daroch, M.; Leng, K.; Johnson, Z.I.; Wang, G. Abundance and Molecular Diversity of Thraustochytrids in Coastal Waters of Southern China. FEMS Microbiol. Ecol. 2017, 93. [Google Scholar] [CrossRef]
- De Philippis, R.; Margheri, M.C.; Materassi, R.; Vincenzini, M. Potential of unicellular cyanobacteria from saline environments as exopolysaccharide producers. Appl. Environ. Microbiol. 1998, 64, 1130–1132. [Google Scholar] [CrossRef] [Green Version]
- De Philippis, R.; Vincenzini, M. Exocellular polysaccharides from cyanobacteria and their possible applications. FEMS Microbiol. Rev. 1998, 22, 151–175. [Google Scholar] [CrossRef]
- Gutiérrez, A.; Prieto, A.; Martínez, A.T. Structural characterization of extracellular polysaccharides produced by fungi from the genus Pleurotus. Carbohydr. Res. 1996, 281, 143–154. [Google Scholar] [CrossRef]
- Decho, A.; Decho, A.W. Microbial exopolymer secretions in ocean environments: Their role(s) in food webs and marine processes. Oceanogr. Mar. Biol. Annu. Rev. 1990, 28, 73–154. [Google Scholar]
- Colwell, R.R.; Pariser, E.R.; Sindkey, A.J. Biotechnology of Marine Polysaccharides; Hemisphere Publishing Corporation: Washington, DC, USA, 1986. [Google Scholar]
- Sogawa, K.; Kodama, E.; Matsuda, M.; Shigeta, S.; Okutani, K. Marine microalgal polysaccharide induces apoptosis in human lymphoid cells. J. Mar. Biotechnol. 1998, 6, 35–38. [Google Scholar]
- Sutherland, I. Biosynthesis and composition of gram-negative bacterial extracellular and wall polysaccharides. Annu. Rev. Microbiol. 1985, 39, 243–270. [Google Scholar] [CrossRef]
- Iwanaga, D.; Gray, D.A.; Fisk, I.D.; Decker, E.A.; Weiss, J.; McClements, D.J. Extraction and Characterization of Oil Bodies from Soy Beans: A Natural Source of Pre-Emulsified Soybean Oil. J. Agric. Food Chem. 2007, 55, 8711–8716. [Google Scholar] [CrossRef] [PubMed]
- Nikiforidis, C.V.; Matsakidou, A.; Kiosseoglou, V. Composition, properties and potential food applications of natural emulsions and cream materials based on oil bodies. RSC Adv. 2014, 4, 25067–25078. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, T.; Sakiyama, R.; Iimi, Y.; Sekine, S.; Abe, E.; Nomura, K.H.; Nomura, K.; Ishibashi, Y.; Okino, N.; Hayashi, M.; et al. Regulation of TG accumulation and lipid droplet morphology by the novel TLDP1 in Aurantiochytrium limacinum F26-b. J. Lipid Res. 2017, 58, 2334–2347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tzen, J.; Cao, Y.; Laurent, P.; Ratnayake, C.; Huang, A. Lipids, Proteins, and Structure of Seed Oil Bodies from Diverse Species. Plant Physiol. 1993, 101, 267–276. [Google Scholar] [CrossRef] [Green Version]
- Gupta, A.; Wilkens, S.; Adcock, J.L.; Puri, M.; Barrow, C.J. Pollen baiting facilitates the isolation of marine thraustochytrids with potential in omega-3 and biodiesel production. J. Ind. Microbiol. Biot. 2013, 40, 1231–1240. [Google Scholar] [CrossRef] [PubMed]
- Chandran, R.; Nivedhini, V.; Parimelazhagan, T. Nutritional composition and antioxidant properties of Cucumis dipsaceus Ehrenb. ex Spach leaf. Sci. World J. 2013, 2013, 890451. [Google Scholar] [CrossRef] [Green Version]
- Chang, K. New Australian thraustochytrids: A Renewable Source of Biofuels, Omega–3 Oils and other Bioproducts. Ph.D. Thesis, University of Tasmania, Tasmania, Australia, 2013. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains/Carbon sources # | FAME | Concentrations of FAMEs, mg/g DW | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| SAFA % | MUFA% | PUFA % | C15:0 | C16:0 | C17:0 | C18:0 | C18:1 | C20:4 | C20:5, EPA | C22:5, DPA | C22:6, DHA | |
| MAN65_GLY | 51.6 ± 7.5 | 2.8 ± 0.4 | 46.7 ± 7.2 | 26.6 ± 5.8 | 54.3 ± 6.7 * | 9.1 ± 1.3 | 9.3 ± 2.3 | 5.5 ± 1.1 | 3.4 ± 1.1 | 5.5 ± 1.6 | 14.6 ± 3.3 | 66.2 ± 11.1 * |
| MAN65_GLU | 43.7 ± 6.2 | 1.8 ± 0.5 | 54.5 ± 10.2 | 16.6 ± 9.8 | 119.4 ± 23.4 * | 9.1 ± 1.8 | 9.6 ± 1.5 | 5.8 ± 1.2 | 3.0 ± 1.6 | 0.8 ± 1.1 | 31.6 ± 4.2 | 147 ± 18.7 * |
| MAN65_FRU | 55.6 ± 4.7 | 3.7 ± 0.3 | 40.5 ± 4 | 8.9 ± 1.7 | 34.5 ± 10.0 * | 3.4 ± 1.0 | 6.7 ± 0.6 | 3.6 ± 0.3 | 5.4 ± 0.9 | 5.8 ± 1.8 | 4.6 ± 0.10 | 33.6 ± 6.8 * |
| MAN70_GLY | 18.5 ± 4.2 | 0.6 ± 0.2 | 80 ± 18.2 | 8.5 ± 1.5 | 8.2 ± 1.7 * | 5.8 ± 1.3 | 9.0 ± 1.6 | 1.2 ± 0.2 | 0.8 ± 0.1 | 3.2 ± 0.6 | 29.7 ± 5.4 | 104.0 ± 22 * |
| MAN70_GLU | 22.5 ± 3.3 | 0.65 ± 0.3 | 76 ± 17.3 | 19.2 ± 3.7 | 14.7 ± 2.4 * | 9.8 ± 1.4 | 8.9 ± 2.7 | 1.2 ± 0.3 | 0.8 ± 0.2 | 1.0 ± 0.6 | 25.7 ± 5.0 | 157.5 ± 21.1 * |
| MAN70_FRU | 21.6 ± 3.0 | 0.4 ± 0.3 | 78 ± 10.6 | 10.5 ± 7.7 | 14.7 ± 4.4 * | 8.5 ± 1.5 | 6.0 ± 1.1 | 0.8 ± 0.3 | 0.5 ±0.3 | 1.8 ± 0.4 | 22.7 ± 4.4 | 123.0 ± 28.1 * |
| Strains | Total # | Carotenoids, % | Cells Color | |||
|---|---|---|---|---|---|---|
| Ast, % | Canth, % | Echi, % | β-Car, % | |||
| MAN70 # | 35.8 ± 2.1 | 7.5 ± 0.8 | 35.6 ± 10.1 | 33.0 ± 7.2 | 21.3 ± 8.2 | O |
| MAN65 # | 55.2 ± 11.2 | 1.1 ± 0.5 | 36.6 ± 8.42 | 32.3 ± 10.2 | 30.7 ± 9.8 | Y |
| Biomass | Lipid | Proteins | Carbohydrates | |||
|---|---|---|---|---|---|---|
| % DW | mg/g DW | % DW | mg/g DW | % DW | mg/g DW | |
| MAN65 cells | 29 ± 2.1 | 300 ± 27.0 | 22 ± 7.0 | 220 ± 17.5 | 9 ± 1.0 | 90 ± 8.2 |
| MAN65 cake | 2.1 ± 0.2 | 1.8 ± 0.1 | 28 ± 3.1 | 280 ± 18.2 | 16 ± 2.1 | 160 ± 17.0 |
| MAN70 cells | 28 ± 3.7 | 295 ± 32.1 | 16 ± 2.6 | 160 ± 12.4 | 17 ± 2.5 | 170 ± 12.2 |
| MAN70 cake | 3.2 ± 0.2 | 2.1 ± 0.7 | 21 ± 3.7 | 210 ± 11.5 | 22 ± 4.5 | 220 ± 13.3 |
| Thraustochytrids/Time/Carbone Source | EPS (g/L) | Carbohydrates, % | Proteins, % | Lipids, % |
|---|---|---|---|---|
| MAN65/ Day 0/ Glu | 0.11 ± 0.1 | 14 ± 1.5 | 7 ± 0.5 | 28 ± 4.5 |
| MAN65/ Day 7/ GLY | 6.2 ± 0.5 | 6 ± 0.6 | 3 ± 0.6 | 18 ± 3.8 |
| MAN65/ Day 7/ FRU | 5.5 ± 0.3 | 7 ± 0.7 | 15 ± 1.4 | 31 ± 11.2 |
| MAN65/ Day 7/ GLU | 6.3 ± 1.2 | 6 ± 0.5 | 8 ± 1.7 | 13 ± 3.3 |
| MAN70/ Day 0/Glu | 0.1 ± 0.1 | 17 ± 2.4 | 11 ± 2.2 | 41 ± 6.5 |
| MAN70/ Day 7/ GLY | 4.0 ± 0.5 | 28 ± 5.5 | 14 ± 3.6 | 17 ± 4.4 |
| MAN70/ Day/ 7 FRU | 4.9 ± 0.3 | 14 ± 3.8 | 10 ± 0.5 | 21 ± 4.57 |
| MAN70/ Day 7/ GLU | 4.0 ± 0.5 | 12 ± 0.5 | 10 ± 0.5 | 32 ± 5.6 |
| pH 12 Treatment | Oil Body Yield, % DW | Lipid Yield, % Oil Body | Protein Yield, % Oil Body |
|---|---|---|---|
| 15 min | 28.1 ± 2.8 | 59.6 ± 5.5 | 0.22 ± 0.1 |
| 3 h | 35.0 ± 2.2 | 67.4 ± 10.8 | 0.15 ± 0.1 |
| 6 h | 20.0 ± 2.8 | 81.0 ± 12.5 | 0.26 ± 0.1 |
| 12 h | 34.0 ± 2.2 | 82.1 ± 11.8 | 0.49 ± 0.1 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nham Tran, T.L.; Miranda, A.F.; Gupta, A.; Puri, M.; Ball, A.S.; Adhikari, B.; Mouradov, A. The Nutritional and Pharmacological Potential of New Australian Thraustochytrids Isolated from Mangrove Sediments. Mar. Drugs 2020, 18, 151. https://doi.org/10.3390/md18030151
Nham Tran TL, Miranda AF, Gupta A, Puri M, Ball AS, Adhikari B, Mouradov A. The Nutritional and Pharmacological Potential of New Australian Thraustochytrids Isolated from Mangrove Sediments. Marine Drugs. 2020; 18(3):151. https://doi.org/10.3390/md18030151
Chicago/Turabian StyleNham Tran, Thi Linh, Ana F. Miranda, Adarsha Gupta, Munish Puri, Andrew S. Ball, Benu Adhikari, and Aidyn Mouradov. 2020. "The Nutritional and Pharmacological Potential of New Australian Thraustochytrids Isolated from Mangrove Sediments" Marine Drugs 18, no. 3: 151. https://doi.org/10.3390/md18030151
APA StyleNham Tran, T. L., Miranda, A. F., Gupta, A., Puri, M., Ball, A. S., Adhikari, B., & Mouradov, A. (2020). The Nutritional and Pharmacological Potential of New Australian Thraustochytrids Isolated from Mangrove Sediments. Marine Drugs, 18(3), 151. https://doi.org/10.3390/md18030151