
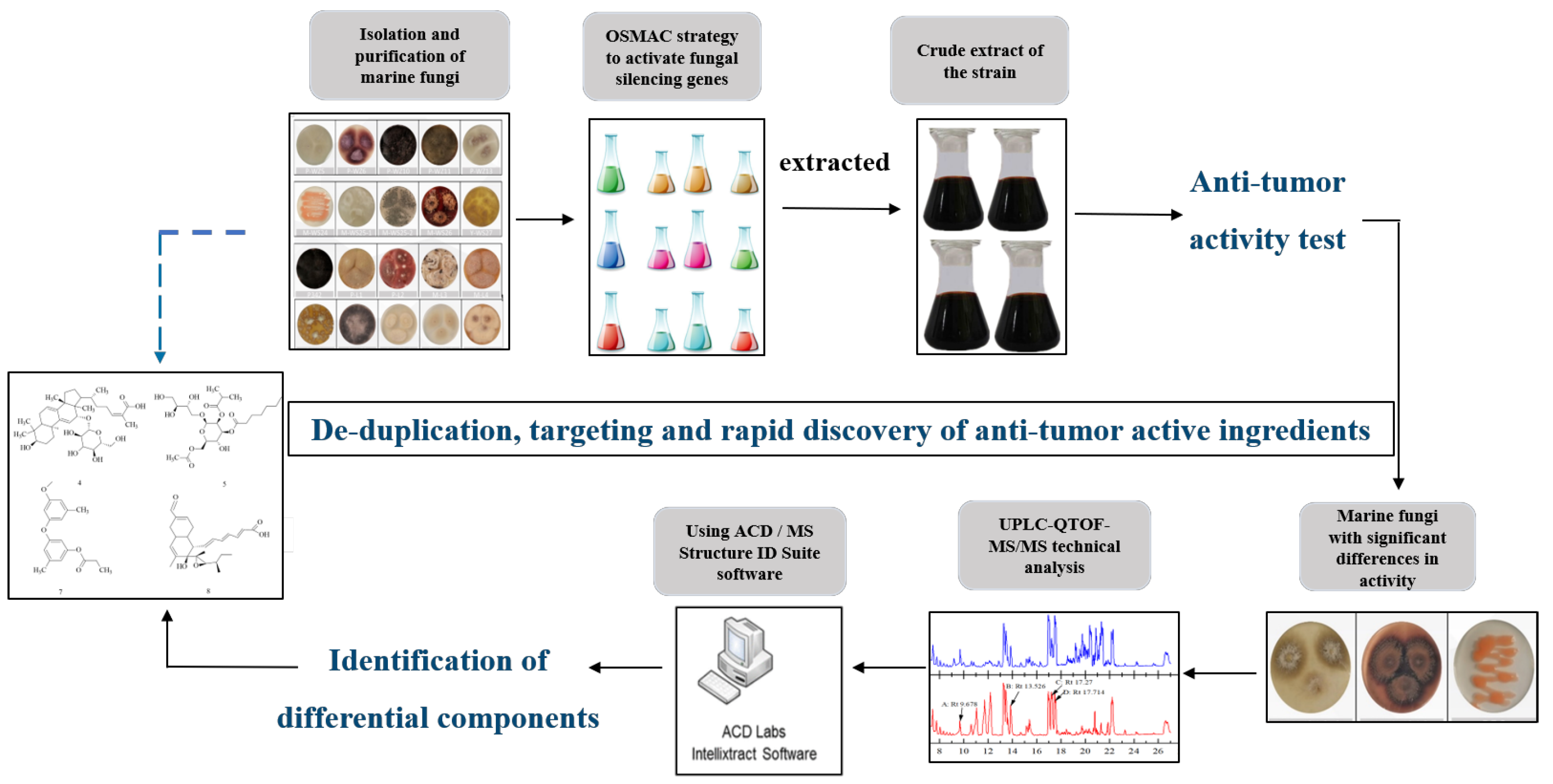
Rapid Discovery of Substances with Anticancer Potential from Marine Fungi Based on a One Strain–Many Compounds Strategy and UPLC-QTOF-MS

Abstract
:
1. Introduction
2. Results
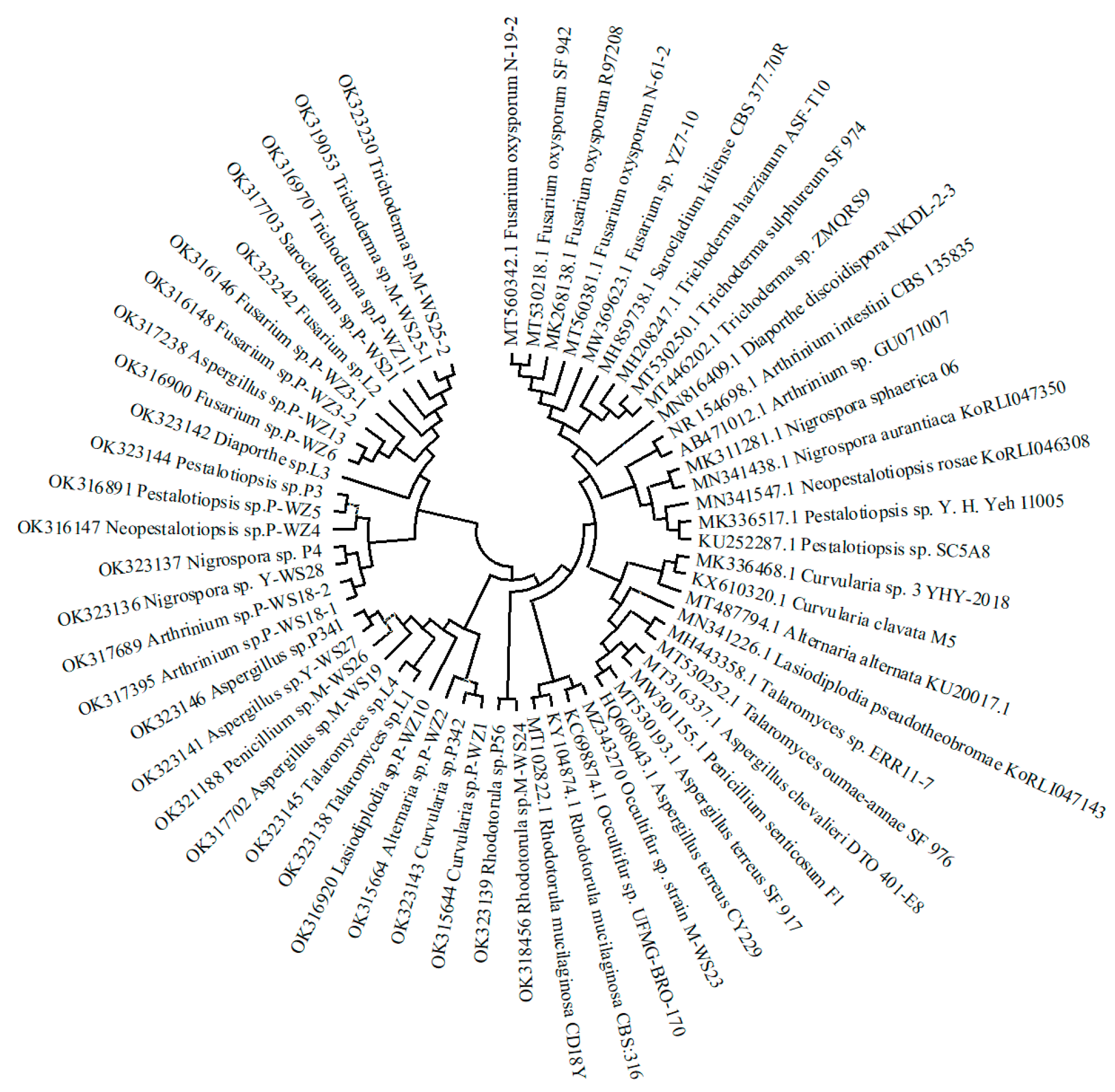
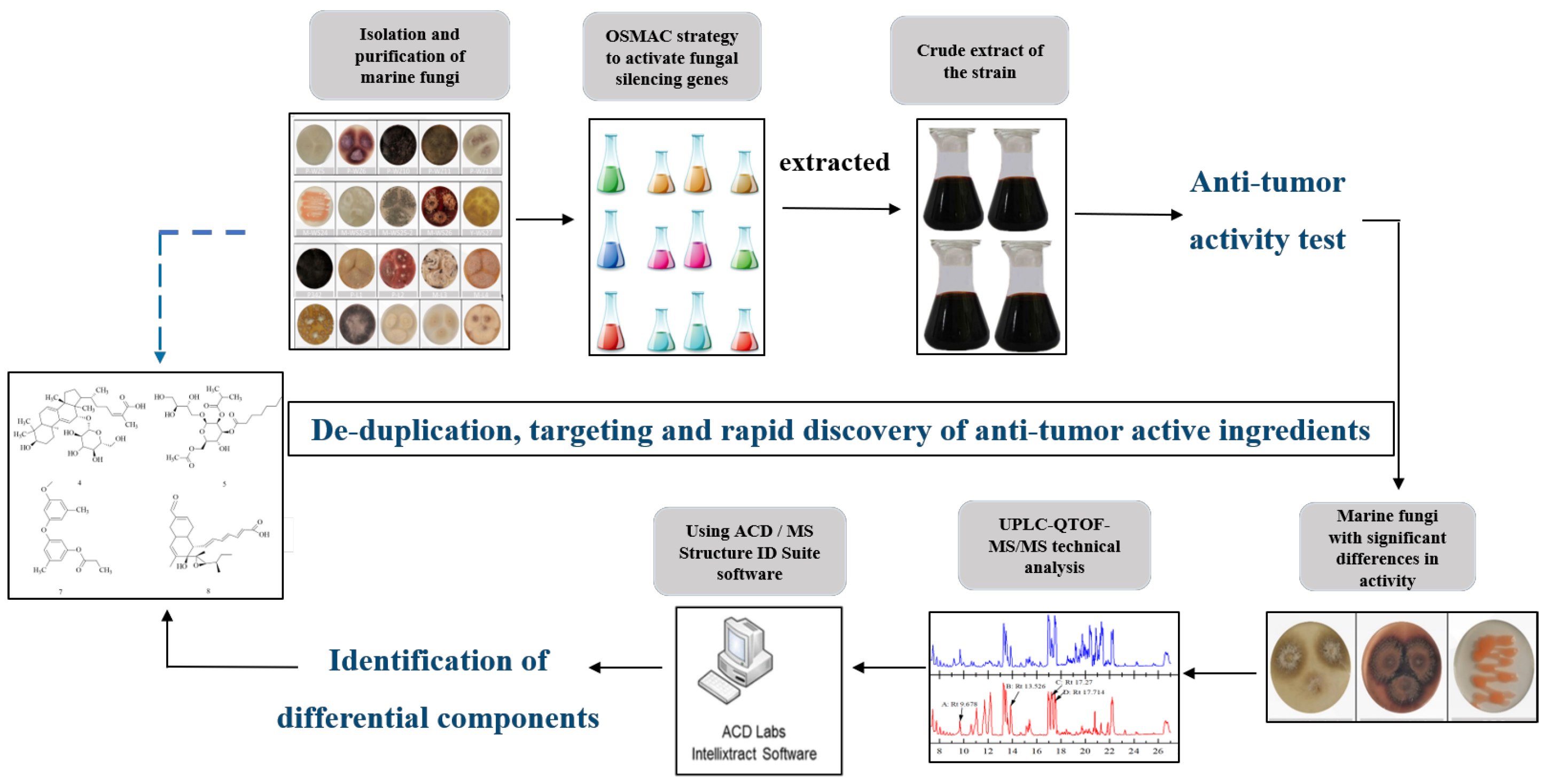
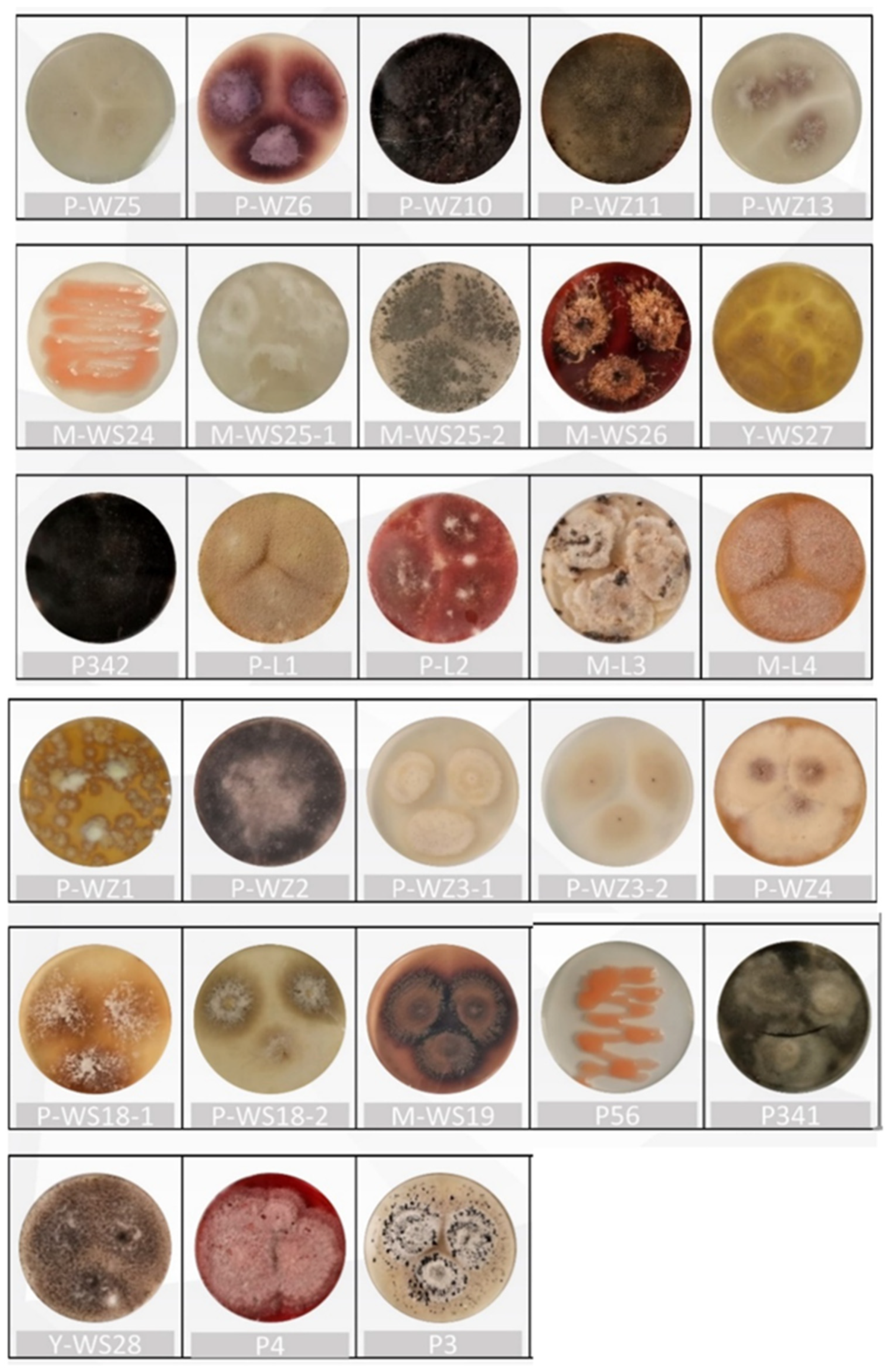
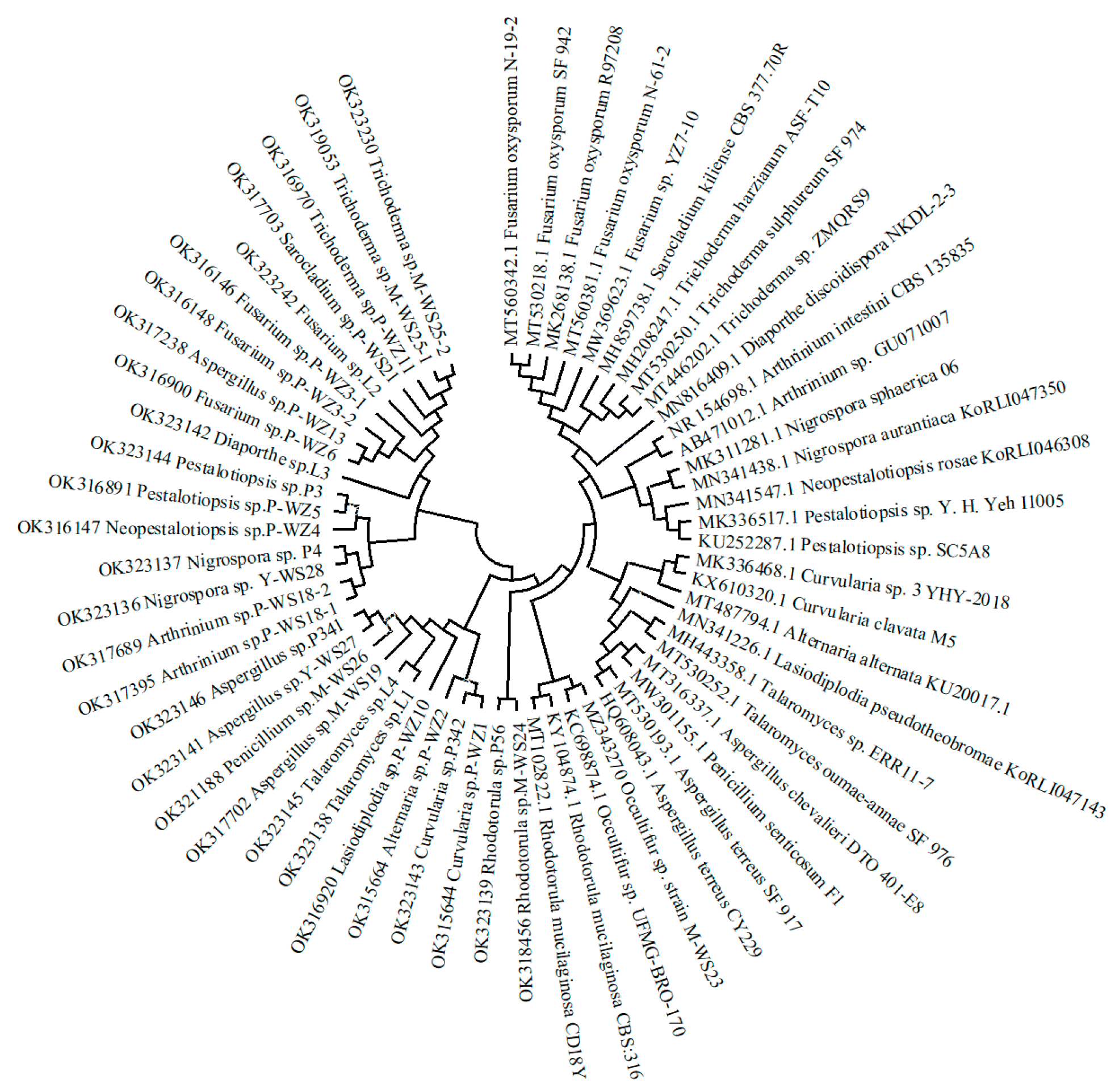
2.1. Isolation and Identification of Marine Fungi
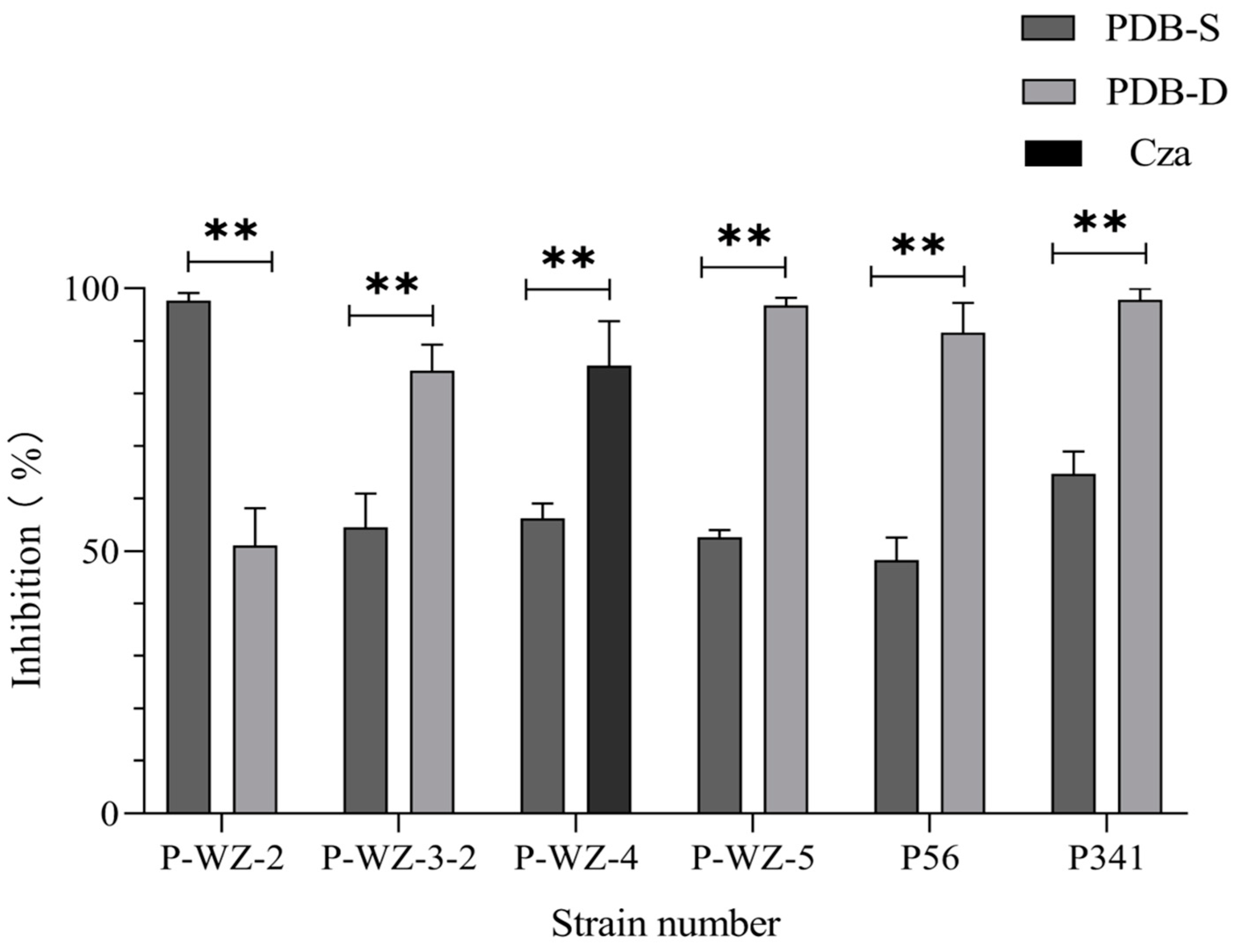
2.2. Using the OSMAC Strategy to Cultivate Marine Fungi and Their Cancer Cell Inhibition Potential
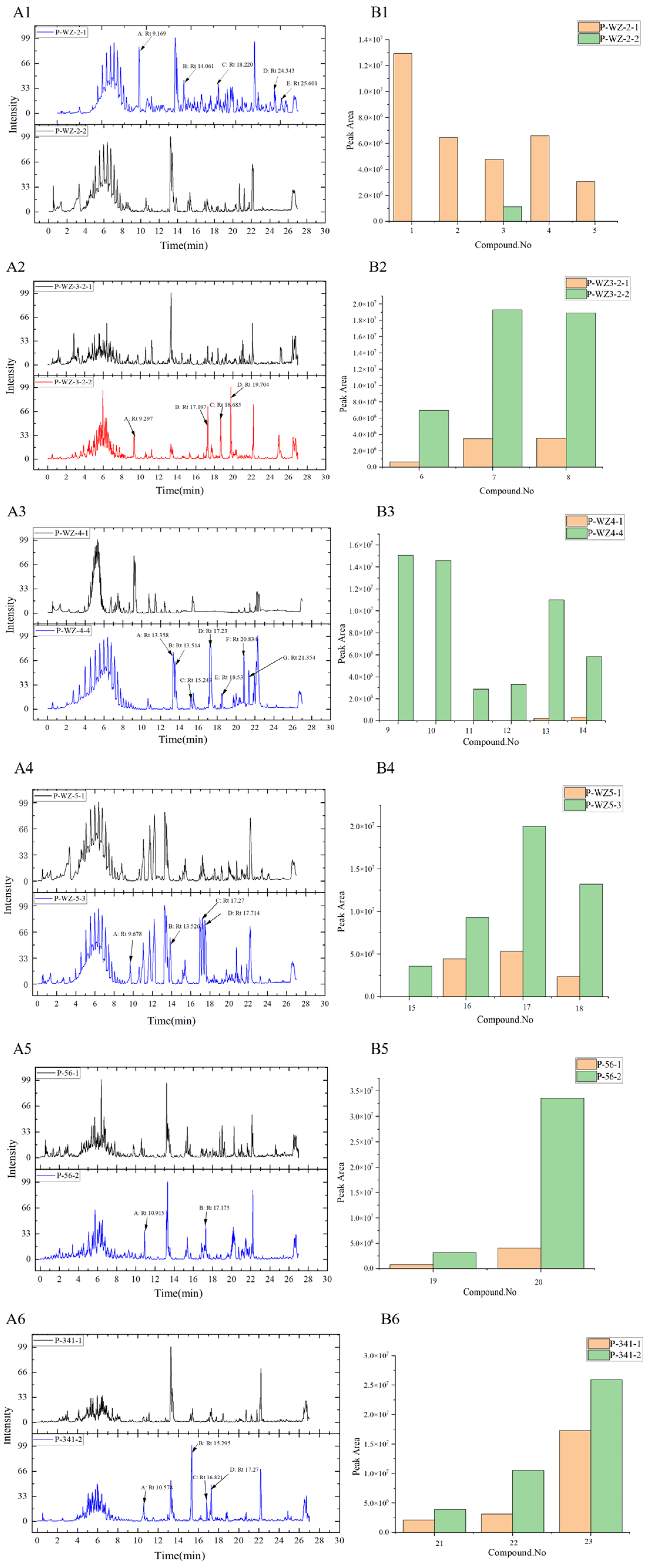
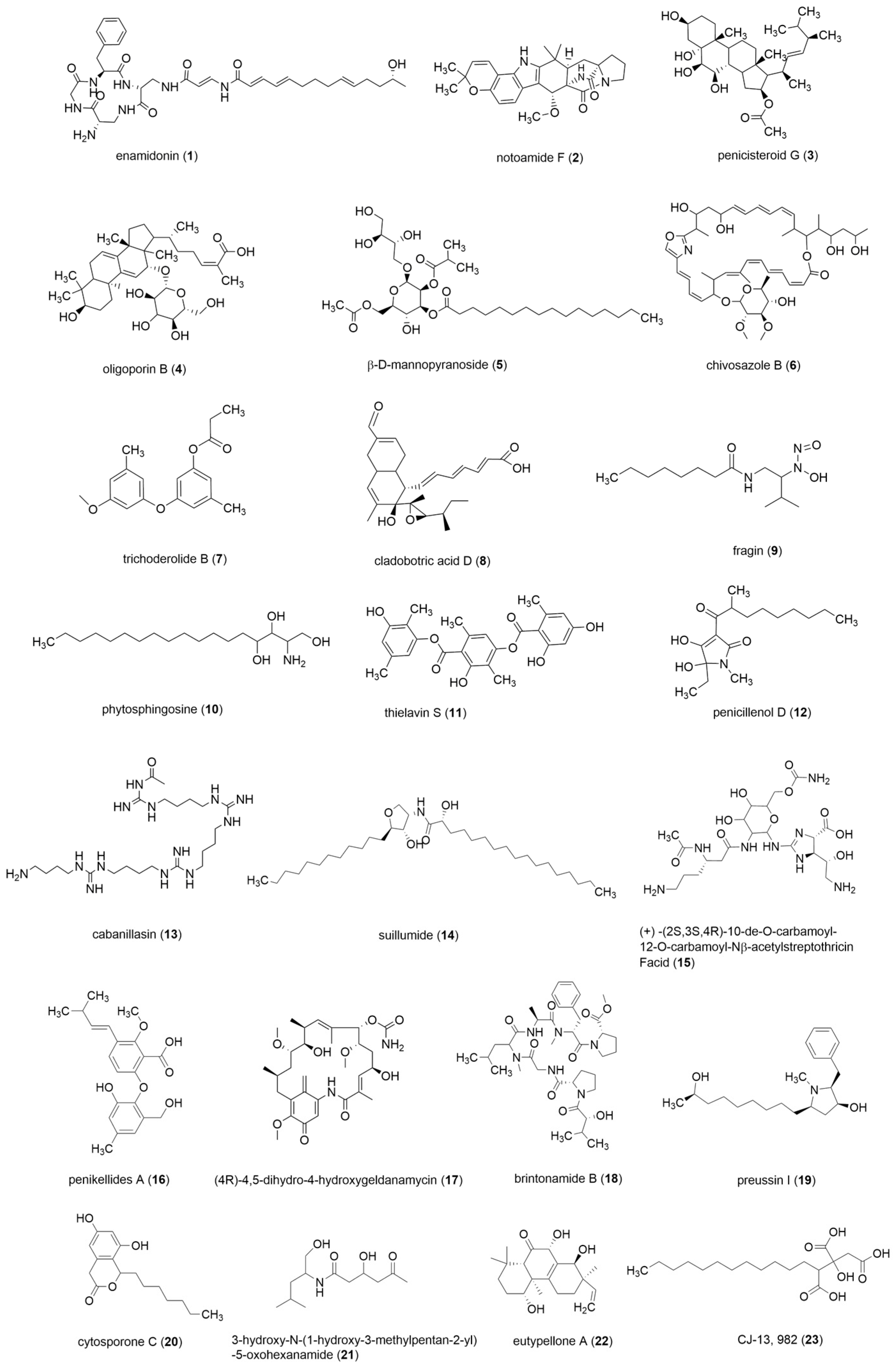
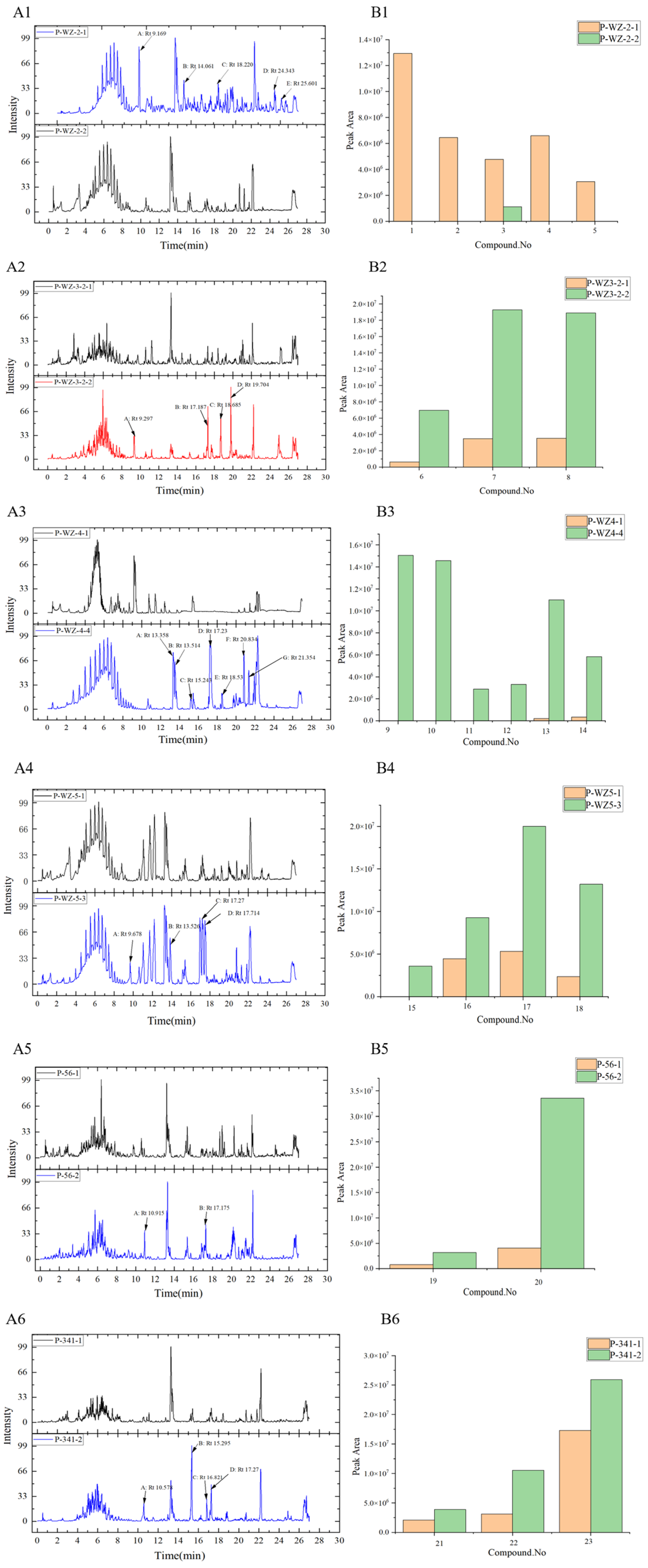
2.3. Analysis of Differential Composition of Secondary Metabolites of Strains by UPLC-QTOF-MS
3. Discussion
4. Materials and Methods
4.1. General Experimental Procedure
4.2. Marine Fungi Material
4.3. OSMAC Strategy
4.4. Cancer Cell Inhibition Rate Test
4.5. UPLC-QTOF-MS Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Florean, C.; Dicato, M.; Diederich, M. Immune-modulating and anti-inflammatory marine compounds against cancer. Semin Cancer Biol. 2022, 80, 58–72. [Google Scholar] [CrossRef] [PubMed]
- Takooree, H.; Aumeeruddy, M.Z.; Rengasamy, K.R.R. A systematic review on black pepper (Piper nigrum L.): From folk uses to pharmacological applications. Crit. Rev. Food Sci. Nutr. 2019, 59, S210–S243. [Google Scholar] [CrossRef] [PubMed]
- Alves-Silva, J.M.; Romane, A.; Efferth, T.; Salgueiro, L. North african medicinal plants traditionally used in cancer therapy. Front Pharmacol. 2017, 8, 383. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Z.Q.; Wang, J.F.; Hao, Y.Y.; Wang, Y. Recent advances in the discovery and development of marine microbial natural products. Mar. Drugs 2013, 11, 700–717. [Google Scholar] [CrossRef] [PubMed]
- Demain, A.L. Importance of microbial natural products and the need to revitalize their discovery. J. Ind. Microbiol Biotechnol. 2014, 41, 185–201. [Google Scholar] [CrossRef] [PubMed]
- Romano, S.; Jackson, S.A.; Patry, S. Extending the “one strain many compounds” (OSMAC) principle to marine microorganisms. Mar. Drugs 2018, 16, 244. [Google Scholar] [CrossRef] [PubMed]
- Pinedo-Rivilla, C.; Aleu, J.; Duran-Patron, R. Cryptic metabolites from marine-derived microorganisms using OSMAC and epigenetic approaches. Mar. Drugs 2022, 20, 84. [Google Scholar] [CrossRef]
- Bode, H.B.; Bethe, B.; Hofs, R.; Zeeck, A. Big effects from small changes: Possible ways to explore nature’s chemical diversity. Chembiochem 2002, 3, 619–627. [Google Scholar] [CrossRef]
- Bin, Y.; Junde, D.; Xiuping, L. New prenylated indole alkaloids from fungus Penicillium sp. derived of mangrove soil sample. Tetrahedron 2014, 70, 3859–3863. [Google Scholar]
- Amagata, T.; Tanaka, M.; Yamada, T.; Doi, M.; Minoura, K. Variation in cytostatic constituents of a sponge-derived Gymnascella dankaliensis by manipulating the carbon source. J. Nat. Prod. 2007, 70, 1731–1740. [Google Scholar] [CrossRef]
- Wang, Z.; Jia, J.; Wang, L. Anti-HSV-1 activity of aspergillipeptide D, a cyclic pentapepetide isolated from fungus Aspergillus sp. SCSIO 41501. Virol. J. 2020, 17, 41. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Li, X.M.; Meng, L.H. Bisthiodiketopiperazines and acorane sesquiterpenes produced by the marine-derived fungus Penicillium adametzioides AS-53 on different culture media. J. Nat. Prod. 2015, 78, 1294–1299. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, J.; Hubmann, G.; Rosenthal, K. Triaging of culture conditions for enhanced secondary metabolite diversity from different bacteria. Biomolecules 2021, 11, 193. [Google Scholar] [CrossRef] [PubMed]
- Farinella, V.F.; Kawafune, E.S.; Tangerina, M. OSMAC strategy integrated with molecular networking for accessing griseofulvin derivatives from endophytic fungi of Moquiniastrum polymorphum (asteraceae). Molecules 2021, 26, 7316. [Google Scholar] [CrossRef] [PubMed]
- Yao, F.H.; Liang, X.; Qi, S.H. Eight new cyclopentenone and cyclohexenone derivatives from the marine-derived fungus Aspergillus sp. SCSIO 41501 by OSMAC strategy. Nat. Prod. Res. 2021, 35, 3810–3819. [Google Scholar] [CrossRef] [PubMed]
- Schrimpe-Rutledge, A.C.; Codreanu, S.G.; Sherrod, S.D. Untargeted metabolomics strategies-challenges and emerging directions. J. Am. Soc. Mass Spectrom. 2016, 27, 1897–1905. [Google Scholar] [CrossRef] [PubMed]
- Tangerina, M.M.P.; Furtado, L.C.; Leite, V.M.B.; Bauermeister, A. Metabolomic study of marine Streptomyces sp.: Secondary metabolites and the production of potential anticancer compounds. PLoS ONE 2020, 15, e244385. [Google Scholar] [CrossRef]
- Van der Hooft, J.J.J.; Mohimani, H.; Bauermeister, A.; Dorrestein, P.C. Linking genomics and metabolomics to chart specialized metabolic diversity. Chem. Soc. Rev. 2020, 49, 3297–3314. [Google Scholar] [CrossRef]
- Blin, K.; Shaw, S.; Steinke, K.; Villebro, R.; Ziemert, N.; Lee, S.Y.; Medema, M.H. AntiSMASH 5.0: Updates to the secondary metabolite genome mining pipeline. Nucleic Acids Res. 2019, 47, W81–W87. [Google Scholar] [CrossRef]
- Pekacar, S.; Deliorman Orhan, D. Investigation of antidiabetic effect of pistacia atlantica Leaves by activity-guided fractionation and phytochemical content analysis by LC-QTOF-MS. Front. Pharmacol. 2022, 13, 826261. [Google Scholar] [CrossRef]
- Croley, T.R.; White, K.D.; Callahan, J.H.; Musser, S.M. The chromatographic role in high resolution mass spectrometry for non-targeted analysis. J. Am. Soc. Mass Spectrom. 2012, 23, 1569–1578. [Google Scholar] [CrossRef] [PubMed]
- McEachran, A.D.; Sobus, J.R.; Williams, A.J. Identifying known unknowns using the US EPA’s CompTox chemistry dashboard. Anal. Bioanal. Chem. 2017, 409, 1729–1735. [Google Scholar] [CrossRef] [PubMed]
- Chervin, J.; Stierhof, M.; Tong, M.H.; Peace, D. Targeted dereplication of microbial natural products by high-resolution MS and predicted LC retention time. J. Nat. Prod. 2017, 80, 1370–1377. [Google Scholar] [CrossRef] [PubMed]
- Tabudravu, J.N.; Pellissier, L.; Smith, A.J.; Subko, K.; Autréau, C.; Feussner, K. LC-HRMS-Database screening metrics for rapid prioritization of samples to accelerate the discovery of structurally new natural products. J. Nat. Prod. 2019, 82, 211–220. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Xu, W.; Fan, R.Z. Penithoketone and peni-thochromones A-L, polyketides from the deep-sea-derived fungus Penicillium thomii YPGA3. J. Nat. Prod. 2020, 83, 2679–2685. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Shi, J.; Liu, R.; Liu, Z. Structure revisions of phenolic bisabolane sesquiterpenes and a ferroptosis inhibitor from the marine-derived fungus Aspergillus versicolor YPH93. J. Nat. Prod. 2023, 86, 830–841. [Google Scholar] [CrossRef]
- Li, Y.H.; Li, X.M.; Yang, S.Q.; Shi, X.S. Antibacterial alkaloids and polyketide derivatives from the deep sea-derived fungus Penicillium cyclopium SD-413. Mar. Drugs 2020, 18, 553. [Google Scholar] [CrossRef]
- Son, S.; Ko, S.K.; Kim, S.M.; Kim, E.; Kim, G.S.; Lee, B. Antibacterial cyclic lipopeptide enamidonins with an enamide-linked acyl chain from a Streptomyces species. J. Nat. Prod. 2018, 81, 2462–2469. [Google Scholar] [CrossRef]
- Tsukamoto, S.; Kato, H.; Samizo, M.; Nojiri, Y. Notoamides F−K, prenylated indole alkaloids isolated from a marine-derived Aspergillus sp. J. Nat. Prod. 2008, 71, 2064–2067. [Google Scholar] [CrossRef]
- Wang, F.; Sarotti, A.M.; Jiang, G.; Huguet-Tapia, J.C.; Zheng, S.L. Waikikiamides A-C: Complex diketopiperazine dimer and diketopiperazine-polyketide hybrids from a Hawaiian marine fungal strain Aspergillus sp. FM242. Org. Lett. 2020, 22, 4408–4412. [Google Scholar] [CrossRef]
- Xie, C.L.; Zhang, D.; Xia, J.M.; Hu, C.C.; Lin, T. Steroids from the deep-sea-derived fungus Penicillium granulatum MCCC 3A00475 induced apoptosis via Retinoid X Receptor (RXR)-α pathway. Mar. Drugs 2019, 17, 178. [Google Scholar] [CrossRef] [PubMed]
- Lee, I.K.; Jang, Y.W.; Yu, S.H.; Yun, B.S. New triterpene glucosides, oligoporins A-C, from Oligoporus tephroleucus protect DNA from Fenton reaction. Bioorg. Med. Chem. Lett. 2007, 17, 4906–4909. [Google Scholar] [CrossRef] [PubMed]
- Wenzel, S.C.; Müller, R. Myxobacterial natural product assembly lines: Fascinating examples of curious biochemistry. Nat. Prod. Rep. 2007, 24, 1211–1224. [Google Scholar] [CrossRef] [PubMed]
- Perlova, O.; Gerth, K.; Kaiser, O.; Hans, A.; Müller, R. Identification and analysis of the chivosazol biosynthetic gene cluster from the myxobacterial model strain Sorangium cellulosum So ce56. J. Biotechnol. 2006, 121, 174–191. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.W.; Gong, B.Q.; Yuan, J.; Li, H.J. l-Phenylalanine alters the privileged secondary metabolite production in the marine-derived fungus Trichoderma erinaceum F1-1. J. Nat. Prod. 2020, 83, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Mitova, M.I.; Lang, G.; Blunt, J.W.; Cummings, N.J. Cladobotric acids A-F: New cytotoxic polyketides from a new zealand Cladobotryum sp. J. Org. Chem. 2006, 71, 492–497. [Google Scholar] [CrossRef] [PubMed]
- Akira, M.; Kosei, H.; Saburo, T. Fragin, a New Biologically Active Metabolite of a Pseudomonas. Agric. Biol. Chem. 1969, 33, 1599–1605. [Google Scholar]
- Tamura, S.; Murayama, A.; Hata, K. Isolation and structural elucidation of fragin, a new plant growth inhibitor produced by a Pseudomonas. Agric. Biol. Chem. 1967, 31, 758–759. [Google Scholar] [CrossRef]
- Nagahara, Y.; Shinomiya, T.; Kuroda, S.; Kaneko, N. Phytosphingosine induced mitochondria-involved apoptosis. Cancer Sci. 2005, 96, 83–92. [Google Scholar] [CrossRef]
- Glenz, R.; Kaiping, A.; Göpfert, D.; Weber, H. The major plant sphingolipid long chain base phytosphingosine inhibits growth of bacterial and fungal plant pathogens. Sci. Rep. 2022, 12, 1081. [Google Scholar] [CrossRef]
- Weber, S.S.; de Souza, A.C.S.; Soares, D.C.L. Chemical profile, antimicrobial potential, and antiaggregant activity of supercritical fluid extract from Agaricus bisporus. Chem. Pap. 2022, 76, 6205–6214. [Google Scholar] [CrossRef]
- De Medeiros, L.S.; Abreu, L.M.; Nielsen, A.; Ingmer, H. Dereplication-guided isolation of depsides thielavins S-T and lecanorins D-F from the endophytic fungus Setophoma sp. Phytochemistry 2015, 11, 1154–1162. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Zhang, Q.Q.; Hu, X. Novel cytotoxic metabolites from the marine-derived fungus Trichoderma citrinoviride. Heterocycles 2014, 89, 189. [Google Scholar] [CrossRef]
- Houard, J.; Aumelas, A.; Noël, T.; Pages, S.; Givaudan, A. Cabanillasin, a new antifungal metabolite, produced by entomopathogenic Xenorhabdus cabanillasii JM26. J. Antibiot. 2013, 66, 617–620. [Google Scholar] [CrossRef] [PubMed]
- León, F.; Brouard, I.; Torres, F.; Quintana, J. A new ceramide from suillus luteus and its cytotoxic activity against human melanoma cells. Chem. Biodivers. 2008, 5, 120–125. [Google Scholar] [CrossRef] [PubMed]
- Gan, M.; Zheng, X.; Gan, L.; Guan, Y.; Hao, X. Streptothricin derivatives from Streptomyces sp. I08A 1776. J. Nat. Prod. 2011, 74, 1142–1147. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Li, X.M.; Li, C.S. Diphenyl ether and benzophenone derivatives from the marine mangrove-derived fungus Penicillium sp. MA-37. Phytochemistry Lett. 2014, 9, 22–25. [Google Scholar] [CrossRef]
- Zhang, F.; Gang, D.; Li, L. Isolation, antimicrobial activity, and absolute configuration of the furylidene tetronic acid core of pestalotic acids A-G. Org. Biomol. Chem. 2012, 10, 5307. [Google Scholar] [CrossRef]
- Li, T.; Ni, S.; Jia, C.; Wang, H.; Sun, G.; Wu, L.; Gan, M. Identification of 4,5-dihydro-4-hydroxygeldanamycins as shunt products of geldanamycin biosynthesis. J. Nat. Prod. 2012, 75, 1480–1484. [Google Scholar] [CrossRef]
- Al-Awadhi, F.H.; Gao, B.; Rezaei, M.A.; Kwan, J.C.; Li, C. Discovery, synthesis, pharmacological profiling, and biological characterization of brintonamides A-E, novel dual protease and GPCR modulators from a marine Cyanobacterium. J. Med. Chem. 2018, 61, 6364–6378. [Google Scholar] [CrossRef]
- Gu, B.B.; Jiao, F.R.; Wu, W.; Jiao, W.H.; Li, L.; Sun, F. Preussins with Inhibition of IL-6 expression from Aspergillus flocculosus 16D-1, a fungus isolated from the marine sponge phakellia fusca. J. Nat. Prod. 2018, 81, 2275–2281. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.X.; Li, H.H.; Chen, Y.C. Cytotoxic secondary metabolites from an endophytic fungal strain of phomopsis phyllanthicola. Chin. J. Org. Chem. 2019, 39, 1475. [Google Scholar] [CrossRef]
- Lena, H.; Debbab, A.; Ngoc, T.D. Polyketides from the mangrove-derived endophytic fungus Acremonium strictum. Tetrahedron Lett. 2014, 55, 3463–3468. [Google Scholar]
- Xiao, J.; Zhang, Q.; Gao, Y.Q. Secondary metabolites from the endophytic Botryosphaeria dothidea of melia azedarach and their antifungal, antibacterial, antioxidant, and cytotoxic activities. J. Agric. Food Chem. 2014, 62, 3584–3590. [Google Scholar] [CrossRef]
- Isaka, M.; Palasarn, S.; Prathumpai, W.; Laksanacharoen, P. Pimarane diterpenes from the endophytic fungus Eutypella sp. BCC 13199. Chem. Pharm. Bull. 2011, 59, 1157–1159. [Google Scholar] [CrossRef]
- Watanabe, S.; Hirai, H.; Kambara, T.; Kojima, Y.; Nishida, H.; Sugiura, A. CJ-13,981 and CJ-13,982, new squalene synthase inhibitors. J. Antibiot. 2001, 54, 1025–1030. [Google Scholar] [CrossRef]
- Xie, F.; Sun, Y.; Zi, Z.F.; Wang, W.J.; Wan, D.Y.; Zhou, H.; Ding, Z.T. Discovery of pyranonaphthoquinones and an eighteen-membered ring macrolide from the rhizospheric soil-derived fungus Phialocephala sp. YUD18001 by OSMAC strategy. Fitoterapia 2023, 25, 105690. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Crude Extracts | Culture Conditions | Growth Inhibition (%) | Crude Extracts | Culture Conditions | Growth Inhibition (%) |
|---|---|---|---|---|---|
| P3-1 | PDB-S | 78.07 | P-WZ-4-1 | PDB-S | 57.23 |
| P3-2 | PDB-W | 84.38 | P-WZ-4-4 | Cza | 87.31 |
| P3-3 | PDB-D | 81.71 | P-WZ-4-5 | Eugon | 77.07 |
| P3-5 | Eugon | 82.32 | P-WZ-4-6 | ME | 62.81 |
| P3-6 | ME | 75.25 | P-WZ-4-7 | Ts | 63.71 |
| P3-7 | Ts | 78.16 | P-WZ-4-8 | DM | 75.47 |
| P3-8 | DM | 91.48 | P-WZ-5-1 | PDB-S | 53.62 |
| P-WZ-1-1 | PDB-S | 96.03 | P-WZ-5-2 | PDB-W | 97.24 |
| P-WZ-1-2 | PDB-W | 87.66 | P-WZ-5-3 | PDB-D | 96.78 |
| P-WZ-1-3 | PDB-D | 78.25 | P-WZ-5-4 | Cza | 81.30 |
| P-WZ-1-4 | Cza | 89.33 | P-WZ-5-5 | Eugon | 66.17 |
| P-WZ-1-5 | Eugon | 93.03 | P-WZ-5-6 | ME | 64.15 |
| P-WZ-1-6 | ME | 73.06 | P-WZ-5-7 | Ts | 85.12 |
| P-WZ-1-7 | Ts | 69.93 | P-WZ-5-8 | DM | 79.09 |
| P-WZ-1-8 | DM | 77.62 | P-WZ-6-1 | PDB-S | 82.76 |
| P4-1 | PDB-S | 99.11 | P-WZ-6-2 | PDB-D | 56.75 |
| P4-2 | PDB-D | 99.16 | P-WZ-3-1-1 | PDB-S | 98.00 |
| P56-1 | PDB-S | 48.30 | P-WZ-3-1-2 | PDB-D | 98.20 |
| P56-2 | PDB-D | 91.62 | P-WZ-3-2-1 | PDB-S | 55.06 |
| P341-1 | PDB-S | 63.71 | P-WZ-3-2-2 | PDB-D | 85.86 |
| P341-2 | PDB-D | 99.37 | P-WZ-2-1 | PDB-S | 97.73 |
| P342-1 | PDB-S | 69.44 | P-WZ-2-2 | PDB-D | 51.06 |
| P342-2 | PDB-W | 64.13 | P-WZ-10-1 | PDB-S | 92.32 |
| P-WZ-13-1 | PDB-S | 91.47 | P-WZ-10-2 | PDB-D | 100.00 |
| P-WZ-13-2 | PDB-D | 93.84 | P-WZ-11-1 | PDB-S | 80.83 |
| P-WS-18-1-1 | PDB-S | 94.31 | P-WZ-11-2 | PDB-D | 86.94 |
| P-WS-18-1-2 | PDB-D | 70.57 | M-WS-26-1 | PDB-S | 91.16 |
| P-WS-18-2-1 | PDB-S | 88.96 | M-WS-26-2 | PDB-D | 74.49 |
| P-WS-18-2-2 | PDB-D | 97.50 | M-WS-24-1 | PDB-S | 81.27 |
| P-L1-1 | PDB-S | 93.51 | M-WS-24-2 | PDB-D | 100.00 |
| P-L1-2 | PDB-D | 93.59 | M-WS-25-1-1 | PDB-S | 90.03 |
| P-L2-1 | PDB-S | 98.62 | M-WS-25-1-2 | PDB-D | 100.00 |
| P-L2-2 | PDB-D | 99.69 | M-WS-25-2-1 | PDB-S | 91.81 |
| P-L3-1 | PDB-S | 67.16 | M-WS-25-2-2 | PDB-D | 68.79 |
| P-L3-2 | PDB-D | 78.49 | M-WS-19-1 | PDB-S | 96.51 |
| P-L4-1 | PDB-S | 86.27 | M-WS-19-2 | PDB-D | 95.37 |
| P-L4-2 | PDB-D | 92.01 | Y-WS-28-1 | PDB-S | 87.09 |
| Y-WS-27-1 | PDB-S | 97.61 | Y-WS-28-2 | PDB-D | 55.33 |
| Y-WS-27-2 | PDB-D | 98.50 |
| Strain | Crude Extracts | IC50 (µM) | |||
|---|---|---|---|---|---|
| Hela | MCF-7 | A549 | HK-2 | ||
| P-WZ-2 | P-WZ-2-1 | 17.01 ± 0.44 | 20.62 ± 0.16 | 14.17 ± 0.18 | 21.6 ± 0.54 |
| P-WZ-2-2 | 63.85 ± 0.68 | - | - | 76.45 ± 0.47 | |
| P-WZ-3-2 | P-WZ-3-2-1 | - | 25.98 ± 0.77 | - | 27.2 ± 0.28 |
| P-WZ-3-2-2 | - | 23.23 ± 0.22 | 38.28 ± 0.80 | 34.28 ± 0.91 | |
| P-WZ-4 | P-WZ-4-1 | 76.12 ± 0.72 | - | - | 76.9 ± 0.45 |
| P-WZ-4-4 | 23.88 ± 0.53 | 14.91 ± 0.98 | 35.88 ± 0.36 | 9.569 ± 0.30 | |
| P-WZ-5 | P-WZ-5-1 | 79.42 ± 0.96 | - | 72.19 ± 0.26 | - |
| P-WZ-5-3 | 25.29 ± 0.80 | 15.9 ± 0.26 | 26.31 ± 0.69 | 19.01 ± 0.91 | |
| P56 | P56-1 | 85.6 ± 0.29 | - | 94.31 ± 0.71 | - |
| P56-2 | 25.91 ± 0.73 | - | 19.64 ± 0.59 | 49.22 ± 0.55 | |
| P341 | P341-1 | 64.88 ± 0.56 | 53.22 ± 0.39 | 71.94 ± 0.26 | 43.31 ± 0.25 |
| P341-2 | 42.14 ± 0.20 | 34.5 ± 0.72 | 23.48 ± 0.23 | 36.66 ± 0.81 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, Y.-T.; Zhao, X.-N.; Zhang, P.-X.; Wang, C.-F.; Li, J.; Wei, X.-Y.; Shi, J.-Q.; Dai, W.; Zhang, Q.; Liu, J.-Q. Rapid Discovery of Substances with Anticancer Potential from Marine Fungi Based on a One Strain–Many Compounds Strategy and UPLC-QTOF-MS. Mar. Drugs 2023, 21, 646. https://doi.org/10.3390/md21120646
Wu Y-T, Zhao X-N, Zhang P-X, Wang C-F, Li J, Wei X-Y, Shi J-Q, Dai W, Zhang Q, Liu J-Q. Rapid Discovery of Substances with Anticancer Potential from Marine Fungi Based on a One Strain–Many Compounds Strategy and UPLC-QTOF-MS. Marine Drugs. 2023; 21(12):646. https://doi.org/10.3390/md21120646
Chicago/Turabian StyleWu, Yu-Ting, Xiao-Na Zhao, Pei-Xi Zhang, Cui-Fang Wang, Jing Li, Xiao-Yue Wei, Jia-Qi Shi, Wang Dai, Qi Zhang, and Jie-Qing Liu. 2023. "Rapid Discovery of Substances with Anticancer Potential from Marine Fungi Based on a One Strain–Many Compounds Strategy and UPLC-QTOF-MS" Marine Drugs 21, no. 12: 646. https://doi.org/10.3390/md21120646
APA StyleWu, Y. -T., Zhao, X. -N., Zhang, P. -X., Wang, C. -F., Li, J., Wei, X. -Y., Shi, J. -Q., Dai, W., Zhang, Q., & Liu, J. -Q. (2023). Rapid Discovery of Substances with Anticancer Potential from Marine Fungi Based on a One Strain–Many Compounds Strategy and UPLC-QTOF-MS. Marine Drugs, 21(12), 646. https://doi.org/10.3390/md21120646