
Diversity and Activity of Bacteria Cultured from a Cup—The Sponge Calyx nicaeensis



Abstract
:1. Introduction
2. Results
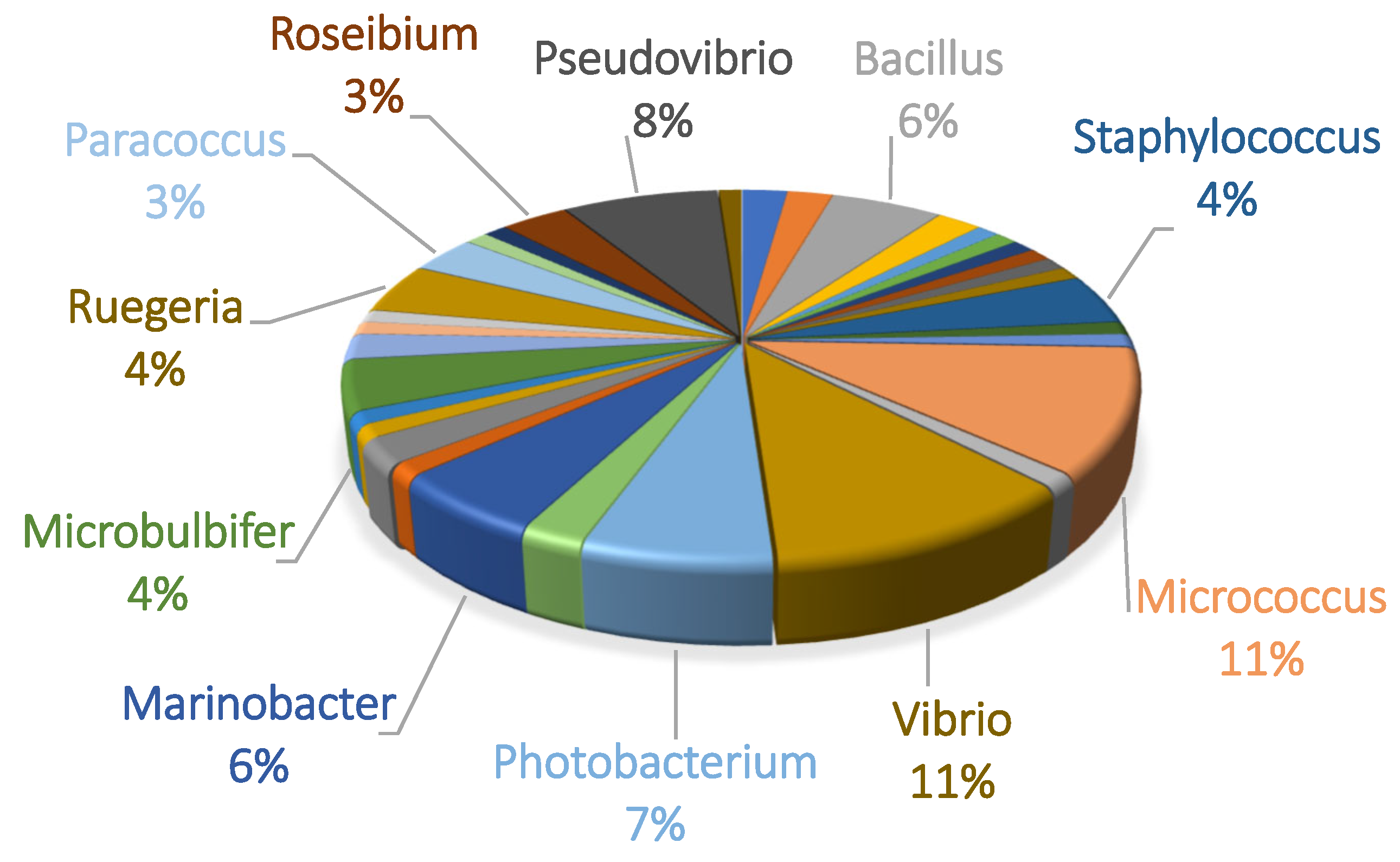
2.1. Microbial Diversity of C. nicaeensis—Associated Cultivable Bacteria
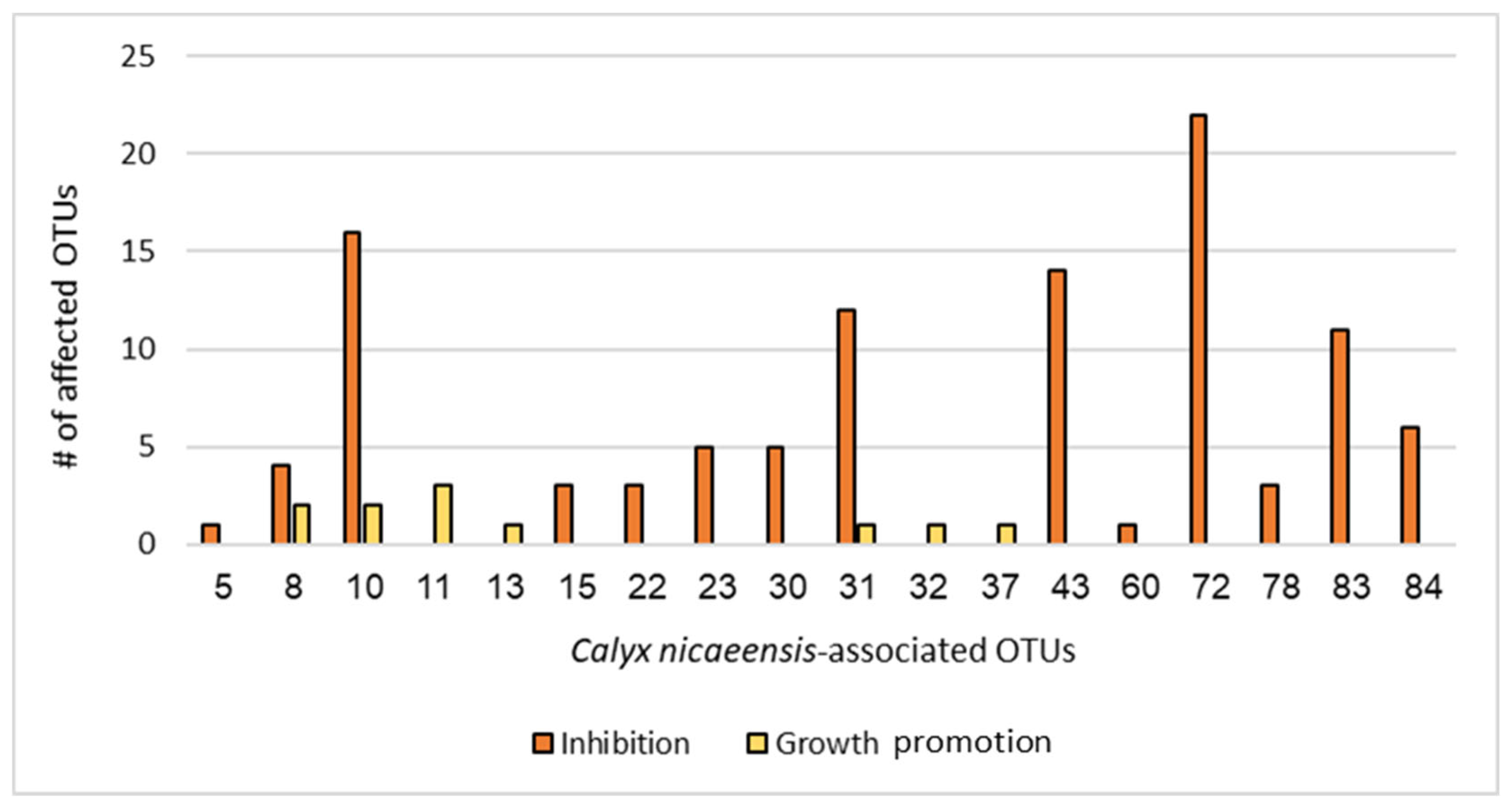
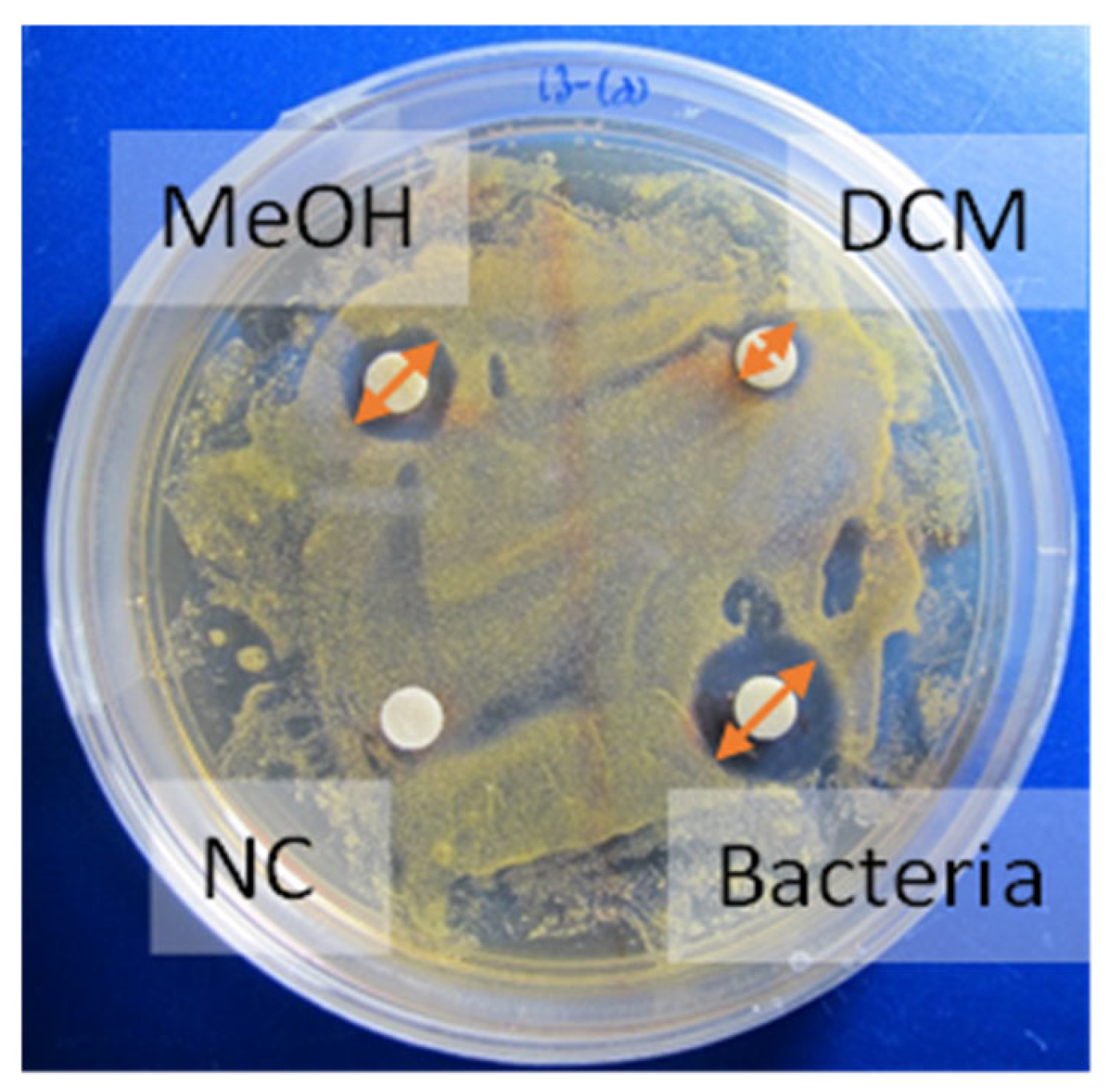
2.2. Competition Assay—Antibacterial Screening
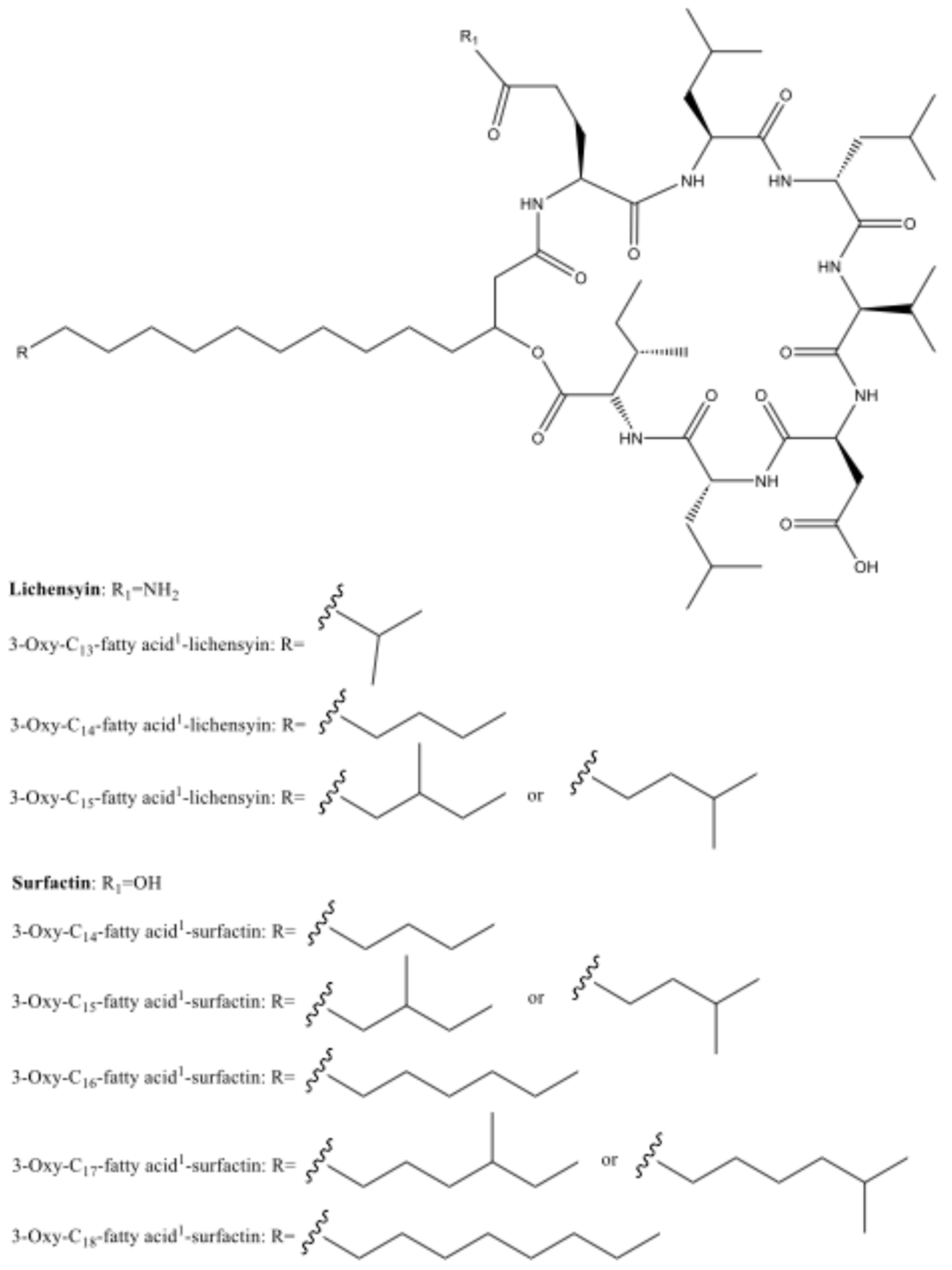
2.3. Chemical Extraction and Separations
3. Discussion
4. Materials and Methods
4.1. General Experimental Procedure
4.2. Sample Collection
4.3. Cultivation of C. nicaeensis—Associated Bacteria
4.4. Morphological Characterization of Bacterial Isolates
4.5. Molecular Identification Using rDNA 16S Phylogenetic Analysis
4.6. Competition Assay—Antibacterial Screening
4.7. Antibacterial Guided Fractionation
4.8. OTU #72 Mass Production for Compound Extraction
4.9. Antibacterial Activity Disk Diffusion Assay from the Mass Production of OTU #72
4.10. Chromatographic Separation Methods
4.11. Structure Elucidation of the Major Component of Fraction 9
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Simpson, T.L. The Epithelia. The Cell Biology of Sponges; Springer: New York, NY, USA, 1984; pp. 42–69. 662p. [Google Scholar]
- Cunningham, J.A.; Liu, A.G.; Bengtson, S.; Donoghue, P.C.J. The origin of animals: Can molecular clocks and the fossil record be reconciled? BioEssays 2017, 39, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Mcclintock, J.B.; Amsler, C.D.; Baker, B.J.; Van Soest, R.W. Ecology of Antarctic Marine Sponges: An Overview. Integr. Comp. Biol. 2005, 45, 359–368. [Google Scholar] [CrossRef] [PubMed]
- Van Soest, R.W.; Boury-Esnault, N.; Vacelet, J.; Dohrmann, M.; Erpenbeck, D.; De Voogd, N.J.; Santodomingo, N.; Vanhoorne, B.; Kelly, M.; Hooper, J.N. Global diversity of sponges (Porifera). PLoS ONE 2012, 7, e35105. [Google Scholar] [CrossRef]
- Strand, R.; Whalan, S.; Webster, N.S.; Kutti, T.; Fang, J.K.H.; Luter, H.M.; Bannister, R.J. The response of a boreal deep-sea sponge holobiont to acute thermal stress. Sci. Rep. 2017, 7, 1160. [Google Scholar] [CrossRef]
- Vogel, S. Current-induced flow through living sponges in nature. Proc. Natl. Acad. Sci. USA 1977, 74, 2069–2071. [Google Scholar] [CrossRef]
- Vacelet, J.; Boury-Esnault, N. Carnivorous sponges. Nature 1995, 373, 333–335. [Google Scholar] [CrossRef]
- Taylor, M.W.; Radax, R.; Steger, D.; Wagner, M. Sponge-Associated Microorganisms: Evolution, Ecology, and Biotechnological Potential. Microbiol. Mol. Biol. 2007, 71, 295–347. [Google Scholar] [CrossRef]
- Rua, C.P.; Trindade-Silva, A.E.; Appolinario, L.R.; Venas, T.M.; Garcia, G.D.; Carvalho, L.S.; Lima, A.; Kruger, R.; Pereira, R.C.; Berlinck, R.G.S.; et al. Diversity and antimicrobial potential of culturable heterotrophic bacteria associated with the endemic marine sponge Arenosclera brasiliensis. PeerJ 2014, 2, e419. [Google Scholar] [CrossRef]
- Thomas, T.; Moitinho-Silva, L.; Lurgi, M.; Björk, J.R.; Easson, C.; Astudillo-García, C.; Olson, J.B.; Erwin, P.M.; López-Legentil, S.; Luter, H.; et al. Diversity, structure and convergent evolution of the global sponge microbiome. Nat. Comm. 2016, 7, 11870. [Google Scholar] [CrossRef]
- Pita, L.; Rix, L.; Slaby, B.M.; Franke, A.; Hentschel, U. The sponge holobiont in a changing ocean: From microbes to ecosystems. Microbiome 2018, 6, 46. [Google Scholar] [CrossRef]
- Kaye, H.R. Sexual reproduction in four Caribbean commercial sponges. II. Oogenesis and transfer of bacterial symbionts. Invert. Repro. Dev. 1991, 19, 13–24. [Google Scholar] [CrossRef]
- Usher, K.M.; Kuo, J.; Fromont, J.; Sutton, D.C. Vertical transmission of cyanobacterial symbionts in the marine sponge Chondrilla australiensis (Demospongiae). Hydrobiologia 2001, 461, 9–13. [Google Scholar] [CrossRef]
- Schmitt, S.; Angermeier, H.; Schiller, R.; Lindquist, N.; Hentschel, U. Molecular microbial diversity survey of sponge reproductive stages and mechanistic insights into vertical transmission of microbial symbionts. Appl. Environ. Micro. 2008, 74, 7694–7708. [Google Scholar] [CrossRef]
- Hill, R.T. Microbes from marine sponges: A treasure trove of biodiversity for natural products discovery. In Microbial Diversity and Bioprospecting; Wiley: Hoboken, NJ, USA, 2003; pp. 177–190. [Google Scholar]
- Schröder, H.C.; Ushijima, H.; Krasko, A.; Gamulin, V.; Thakur, N.L.; Diehl-Seifert, B.; Müller, I.M.; Müller, W.E.G. Emergence and disappearance of an immune molecule, an antimicrobial lectin, in basal metazoa: A tachylectin-related protein in the sponge Suberites domuncula. J Biol. Chem. 2003, 278, 32810–32817. [Google Scholar] [CrossRef]
- Wiens, M.; Korzhev, M.; Krasko, A.; Thakur, N.L.; Perović-Ottstadt, S.; Breter, H.J.; Ushijima, H.; Diehl-Seifert, B.; Müllert, I.M.; Müller, W.E.G. Innate immune defense of the sponge Suberites domuncula against bacteria involves a MyD88-dependent signaling pathway: Induction of a perforin-like molecule. J. Biol. Chem. 2005, 280, 27949–27959. [Google Scholar] [CrossRef]
- Alain, K.; Querellou, J. Cultivating the uncultured: Limits, advances and future challenges. Extremophiles 2009, 13, 583–594. [Google Scholar] [CrossRef]
- Lavy, A.; Keren, R.; Haber, M.; Schwartz, I.; Ilan, M. Implementing sponge physiological and genomic information to enhance the diversity of its culturable associated bacteria. FEMS Microb. Ecol. 2014, 87, 486–502. [Google Scholar] [CrossRef]
- Keren, R.; Lavy, A.; Ilan, M. Increasing the richness of culturable arsenic-tolerant bacteria from Theonella swinhoei by adding of sponge skeleton to the growth medium. Microb. Ecol. 2016, 71, 873–886. [Google Scholar] [CrossRef]
- Dat, T.T.H.; Steinert, G.; Cuc, N.T.K.; Smidt, H.; Sipkema, D. Bacteria Cultivated From Sponges and Bacteria Not Yet Cultivated From Sponges—A Review. Front. Microbio. 2021, 12, 737925. [Google Scholar] [CrossRef]
- Busch, K.; Slaby, B.M.; Bach, W.; Boetius, A.; Clefsen, I.; Colaço, A.; Creemers, M.; Cristobo, J.; Federwisch, L.; Franke, A.; et al. Biodiversity, environmental drivers, and sustainability of the global deep-sea sponge microbiome. Nat. Commun. 2022, 13, 5160. [Google Scholar] [CrossRef]
- Webster, N.S.; Hill, R.T. The culturable microbial community of the Great Barrier Reef sponge Rhopaloeides odorabile is dominated by an α-Proteobacterium. Mar. Biol. 2001, 138, 843–851. [Google Scholar] [CrossRef]
- Olson, J.B.; McCarthy, P.J. Associated bacterial communities of two deep-water sponges. Aquat. Microb. Ecol. 2005, 39, 47–55. [Google Scholar] [CrossRef]
- Sipkema, D.; Schippers, K.; Maalcke, W.J.; Yang, Y.; Salim, S.; Blanch, H.W. Multiple approaches to enhance the cultivability of bacteria associated with the marine sponge Haliclona (gellius) sp. Appl. Environ. Microbiol. 2011, 77, 2130–2140. [Google Scholar] [CrossRef]
- Jin, T.; Wang, Y.; Huang, Y.; Xu, J.; Zhang, P.; Wang, N.; Liu, X.; Chu, H.; Liu, G.; Jiang, H.; et al. Taxonomic structure and functional association of foxtail millet root microbiome. GigaScience 2017, 6, 1–12. [Google Scholar] [CrossRef]
- Taylor, J.A.; Palladino, G.; Wemheuer, B.; Steinert, G.; Sipkema, D.; Williams, T.J.; Thomas, T. Phylogeny resolved, metabolism revealed: Functional radiation within a widespread and divergent clade of sponge symbionts. ISME J. 2021, 15, 503–519. [Google Scholar] [CrossRef] [PubMed]
- Malve, H. Exploring the ocean for new drug developments: Marine pharmacology. JPBS 2016, 8, 83. [Google Scholar] [CrossRef] [PubMed]
- Calado, R.; Mamede, R.; Cruz, S.; Leal, M.C. Updated trends on the biodiscovery of new marine natural products from invertebrates. Mar. Drugs 2022, 20, 389. [Google Scholar] [CrossRef]
- Varijakzhan, D.; Loh, J.Y.; Yap, W.S.; Yusoff, K.; Seboussi, R.; Lim, S.H.E.; Lai, K.S.; Chong, C.M. Bioactive compounds from marine sponges: Fundamentals and applications. Mar. Drugs 2021, 19, 246. [Google Scholar] [CrossRef]
- Li, P.; Lu, H.; Zhang, Y.; Zhang, X.; Liu, L.; Wang, M.; Liu, L. The natural products discovered in marine sponge-associated microorganisms: Structures, activities, and mining strategy. Front. Mar. Sci. 2023, 10, 1191858. [Google Scholar] [CrossRef]
- Proksch, P.; Edrada, R.; Ebel, R. Drugs from the seas—Current status and microbiological implications. Appl. Microbiol. Biotechnol. 2002, 59, 125–134. [Google Scholar]
- Imhoff, J.F.; Labes, A.; Wiese, J. Bio-mining the microbial treasures of the ocean: New natural products. Biotechnol. Adv. 2011, 29, 468–482. [Google Scholar] [CrossRef] [PubMed]
- Waters, A.L.; Hill, R.T.; Place, A.R.; Hamann, M.T. The expanding role of marine microbes in pharmaceutical development. COBIOT 2010, 21, 780–786. [Google Scholar] [CrossRef] [PubMed]
- Hentschel, U.; Schmid, M.; Wagner, M.; Fieseler, L.; Gernert, C.; Hacker, J. Isolation and phylogenetic analysis of bacteria with antimicrobial activities from the Mediterranean sponges Aplysina aerophoba and Aplysina cavernicola. FEMS Microbiol. Ecol. 2001, 35, 305–312. [Google Scholar] [CrossRef]
- Faulkner, D.J.; Unson, M.D.; Bewley, C.A. The chemistry of some sponges and their symbionts. Pure Appl. Chern 1994, 66, 1983–1990. [Google Scholar] [CrossRef]
- Haber, M.; Ilan, M. Diversity and antibacterial activity of bacteria cultured from Mediterranean Axinella spp. sponges. J. Appl. Microbiol. 2014, 116, 519–532. [Google Scholar] [CrossRef]
- Devkar, H.U.; Thakur, N.L.; Kaur, P. Marine-derived antimicrobial molecules from the sponges and their associated bacteria. Can. J. Microbiol. 2022, 69, 1–16. [Google Scholar] [CrossRef]
- Mayer, A.M.; Glaser, K.B.; Cuevas, C.; Jacobs, R.S.; Kem, W.; Little, R.D.; McIntosh, J.M.; Newman, D.J.; Potts, B.C.; Shuster, D.E. The odyssey of marine pharmaceuticals: A current pipeline perspective. TIPS 2010, 31, 255–265. [Google Scholar] [CrossRef]
- Morrow, K.M.; Fiore, C.L.; Lesser, M.P. Environmental drivers of microbial community shifts in the giant barrel sponge, Xestospongia muta, over a shallow to mesophotic depth gradient. Environ. Microbiol. 2016, 18, 2025–2038. [Google Scholar] [CrossRef]
- Steinert, G.; Taylor, M.W.; Deines, P.; Simister, R.L.; de Voogd, N.J.; Hoggard, M.; Schupp, P.J. In four shallow and mesophotic tropical reef sponges from Guam the microbial community largely depends on host identity. PeerJ 2016, 4, e1936. [Google Scholar] [CrossRef]
- Pyle, R.L.; Copus, J.M. Mesophotic Coral Ecosystems: Introduction and Overview. In Mesophotic Coral Ecosystems; Springer: Cham, Switzerland, 2019; pp. 3–27. [Google Scholar]
- Trainito, E.; Baldacconi, R.; Vesna, M. All-around rare and generalist: Countercurrent signals from the updated distribution of Calyx nicaeensis (Risso, 1826) (Porifera, Demospongiae). Stud. Mar. 2020, 33, 5–17. [Google Scholar]
- Cerrano, C.; Molinari, A.; Bernat, P.; Baldacconi, B.; Calcinai, B.; Macic, V. Calyx nicaeensis (Risso, 1826) (Porifera, Demospongiae). Is it a rare and threatened species? Rapp. Comm. Int. Mer Médit. 2013, 40, 771. [Google Scholar]
- Idan, T.; Shefer, S.; Feldstein, T.; Yahel, R.; Huchon, D.; Ilan, M. Shedding light on an East-Mediterranean mesophotic sponge ground community and the regional sponge fauna. Mediterr. Mar. Sci. 2018, 19, 84–106. [Google Scholar] [CrossRef]
- Faitorusso, E.; Magno, S.; Mayol, L.; Santacroce, C.; Sica, D. Calysterol: A C29 cyclopropene-containing marine sterol from the sponge Calyx nicaensis. Tetrahedron 1975, 31, 1915–1916. [Google Scholar] [CrossRef]
- Steiner, E.; Djerassi, C.; Fattorusso, E.; Magno, S.; Mayol, L.; Santacroce, C.; Sica, D. Isolation, Structure Determination and Synthesis of New Acetylenic Steroids from the Sponge Calyx nicaeensis. Helv. Chim. Acta 1977, 60, 475–481. [Google Scholar] [CrossRef]
- Itoh, T.; Sica, D.; Djerassi, C. Minor and trace sterols in marine invertebrates. 36.(24S)-24H-Isocalysterol: A new steroidal cyclopropene from the marine sponge Calyx niceaensis. J. Org. Chem. 1983, 48, 890–892. [Google Scholar] [CrossRef]
- Ha, T.T.; Djerassi, C. Minor and trace sterols in marine invertebrates. 62. novel coprostanols with unusual side chains from the marine sponge Calyx nicaeensis. Steroids 1989, 53, 387–499. [Google Scholar] [CrossRef]
- Minale, L.; Riccio, R.; Scalona, O.; Sodano, G.; Fattorusso, E.; Magno, S.; Santacroce, C. Metabolism in Porifera. VII. Conversion of [7,7-3H2]-fucosterol into calysterol by the sponge Calyx niceaensis. Experientia 1977, 33, 1550–1552. [Google Scholar] [CrossRef]
- Doss, G.A.; Margot, C.; Sodano, G.; Djerassi, C. Biosynthetic Studies of Marine Lipids. 21.l Experimental Demonstration of a Biosynthetic Interconversion of Cyclopropene Sterols in the Sponge Calyx nicaeensis. Tetrahedron Lett. 1988, 29, 6051–6054. [Google Scholar] [CrossRef]
- Doss, G.A.; Silva, C.J.; Djerassi, C. Biosynthetic Studies of Marine Lipids. 22: Calysterol: Some aspects of sterol cyclopropene biosynthesis in the sponge Calyx nicaeensis. Tetrahedron 1989, 45, 1273–1282. [Google Scholar] [CrossRef]
- Dogan, E.; Demir, O.; Sertdemir, M.; Saracli, M.A.; Konuklugil, B. Screening of the selected marine sponges from the coasts of Turkey for antimicrobial activity. IJMS 2018, 47, 1193–1198. [Google Scholar]
- Li, Y.; Yang, S.; Mu, B. The surfactin and lichenysin isoforms produced by Bacillus licheniformis HSN 221. Anal. Lett. 2010, 43, 929–940. [Google Scholar] [CrossRef]
- Behera, H.T.; Mojumdar, A.; Ray, L. Biology, genetic aspects and oxidative stress response of actinobacteria and strategies for bioremediation of toxic metals. In Microbial Biodegradation and Bioremediation; Elsevier: Amsterdam, The Netherlands, 2022; pp. 181–192. [Google Scholar]
- Schmitt, S.; Tsai, P.; Bell, J.; Fromont, J.; Ilan, M.; Lindquist, N.; Perez, T.; Rodrigo, A.; Schupp, P.J.; Vacelet, J.; et al. Assessing the complex sponge microbiota: Core, variable and species-specific bacterial communities in marine sponges. ISME J. 2012, 6, 564–576. [Google Scholar] [CrossRef]
- Laatsch, H. Marine bacterial metabolites. In Frontiers in Marine Biotechnology; Horizon Bioscience: Norfolk, UK, 2006; pp. 225–288. [Google Scholar]
- Fenical, W.; Jensen, P.R. Developing a new resource for drug discovery: Marine actinomycete bacteria. Nat. Chem. Biol. 2006, 2, 666–673. [Google Scholar] [CrossRef] [PubMed]
- Umezawa, H. New antibiotics, bleomycin A and B. J. Antibiot. 1966, 19, 200–209. [Google Scholar]
- Takahashi, Y. Exploitation of new microbial resources for bioactive compounds and discovery of new actinomycetes. Actinomycetologica 2004, 18, 54–61. [Google Scholar] [CrossRef]
- Reddy, G.S.N.; Manasa, B.P.; Singh, S.K.; Shivaji, S. Paenisporosarcina indica sp. nov., a psychrophilic bacterium from a glacier, and reclassification of Sporosarcina antarctica Yu et al., 2008 as Paenisporosarcina antarctica comb. nov. and emended description of the genus Paenisporosarcina. IJSEM 2013, 63 Pt 8, 2927–2933. [Google Scholar] [CrossRef]
- Schultz, J.; Rosado, A.S. Extreme environments: A source of biosurfactants for biotechnological applications. Extremophiles 2020, 24, 189–206. [Google Scholar] [CrossRef]
- Lasek, R.; Szuplewska, M.; Mitura, M.; Decewicz, P.; Chmielowska, C.; Pawlot, A.; Sentkowska, D.; Czarnecki, J.; Bartosik, D. Genome structure of the opportunistic pathogen Paracoccus yeei (Alphaproteobacteria) and identification of putative virulence factors. Front. Microbiol. 2018, 9, 2553. [Google Scholar] [CrossRef]
- Lorenz, N.; Reiger, M.; Toro-Nahuelpan, M.; Brachmann, A.; Poettinger, L.; Plener, L.; Lassak, J.; Jung, K. Identification and initial characterization of prophages in Vibrio campbellii. PLoS ONE 2016, 11, e0156010. [Google Scholar] [CrossRef]
- Bottone, E.J. Bacillus cereus, a volatile human pathogen. Clin. Microbiol. Rev. 2010, 23, 382–398. [Google Scholar] [CrossRef]
- Collins FW, J.; O’Connor, P.M.; O’Sullivan, O.; Rea, M.C.; Hill, C.; Ross, R.P. Formicin—A novel broad-spectrum two component lantibiotic produced by Bacillus paralicheniformis APC 1576. Microbiology 2016, 162, 1662–1671. [Google Scholar] [CrossRef] [PubMed]
- Mearns-Spragg, A.; Bregu, M.; Boyd, K.G.; Burgess, J.G. Cross-species, induction and enhancement of antimicrobial activity produced by epibiotic bacteria from marine algae and invertebrates, after exposure to terrestrial bacteria. LAM 1998, 27, 142–146. [Google Scholar] [CrossRef] [PubMed]
- Kanagasabhapathy, M.; Nagata, S. Cross-species induction of antibacterial activity produced by epibiotic bacteria isolated from Indian marine sponge Pseudoceratina purpurea. World J. Microbiol. Biotechnol. 2008, 24, 687–691. [Google Scholar] [CrossRef]
- Muscholl-Silberhorn, A.; Thiel, V.; Imhoff, J.F. Abundance and bioactivity of cultured sponge-associated bacteria from the Mediterranean Sea. Microb. Ecol. 2008, 55, 94–106. [Google Scholar] [CrossRef] [PubMed]
- Chelossi, E.; Pantile, R.; Pronzato, R.; Milanese, M.; Hentschel, U. Bacteria with antimicrobial properties isolated from the Mediterranean sponges Chondrilla nucula and Petrosia ficiformis. AME 2007, 49, 157–163. [Google Scholar] [CrossRef]
- Gnanamanickam, S.S.; Smith, D.A. Selective toxicity of isoflavonoid phytoalexins to gram-positive bacteria. Phytopathology 1980, 70, 894. [Google Scholar] [CrossRef]
- Nikaido, H. Multidrug Efflux Pumps of Gram-Negative Bacteria. J. Bacteriol. 1996, 178, 5853–5859. [Google Scholar] [CrossRef]
- Abdelmohsen, U.R.; Bayer, K.; Hentschel, U. Diversity, abundance and natural products of marine sponge-associated actinomycetes. Nat. Prod. Rep. 2014, 31, 381–399. [Google Scholar] [CrossRef]
- Hellio, C.; Bremer, G.; Pons, A.M.; Le Gal, Y.; Bourgougnon, N. Inhibition of the development of microorganisms (bacteria and fungi) by extracts of marine algae from Brittany, France. Appl. Microbiol. Biotechnol. 2000, 54, 543–549. [Google Scholar] [CrossRef]
- Angelova, B.; Schmauder, H.-P. Lipophilic compounds in biotechnology-interactions with cells and technological problems. J. Biotech. 1999, 67, 13–32. [Google Scholar] [CrossRef]
- Hino, M.; Fujie, A.; Iwamoto, T.; Hori, Y.; Hashimoto, M.; Tsurumi, Y.; Sakamoto, K.; Takase, S.; Hashimoto, S. Chemical diversity in lipopeptide antifungal antibiotics. JIMB 2001, 27, 157–162. [Google Scholar] [CrossRef] [PubMed]
- Das, P.; Mukherjee, S.; Sen, R. Antimicrobial potential of a lipopeptide biosurfactant derived from a marine Bacillus circulans. J. Appl. Microbiol. 2008, 104, 1675–1684. [Google Scholar] [CrossRef] [PubMed]
- Sachdev, D.P.; Cameotra, S.S. Biosurfactants in agriculture. Appl. Microbiol. Biotechnol. 2013, 97, 1005–1016. [Google Scholar] [CrossRef]
- Yeak KY, C.; Perko, M.; Staring, G.; Fernandez-Ciruelos, B.M.; Wells, J.M.; Abee, T.; Wells-Bennik, M.H. Lichenysin Production by Bacillus licheniformis Food Isolates and Toxicity to Human Cells. Front. Microbiol. 2022, 13, 831003. [Google Scholar] [CrossRef]
- Zammuto, V.; Rizzo, M.G.; De Pasquale, C.; Ferlazzo, G.; Caccamo, M.T.; Magazù, S.; Guglielmino, S.P.P.; Gugliandolo, C. Lichenysin-like Polypeptide Production by Bacillus licheniformis B3-15 and Its Antiadhesive and Antibiofilm Properties. Microorganisms 2023, 11, 1842. [Google Scholar] [CrossRef]
- Rangarajan, V.; Clarke, K.G. Process development and intensification for enhanced production of Bacillus lipopeptides. BGER 2015, 31, 46–68. [Google Scholar]
- Coronel León, J.; Manresa Presas, M.Á.; Marqués Villavecchia, A.M. Lichenysin production and application in the pharmaceutical field. In Recent Advances in Pharmaceutical Sciences VI; Research Signpost: Kerala, India, 2016; Chapter 9; pp. 147–163. [Google Scholar]
- Coronel-León, J.; Marqués, A.M.; Bastida, J.; Manresa, A. Optimizing the production of the biosurfactant lichenysin and its application in biofilm control. J. Appl. Microbiol. 2016, 120, 99–111. [Google Scholar] [CrossRef]
- Hue, N.; Serani, L.; Laprvote, O. Structural investigation of cyclic peptidolipids from Bacillus subtilis by high-energy tandem mass spectrometry. Rapid Commun. Mass Spectrom. 2001, 15, 203–209. [Google Scholar] [CrossRef]
- Anuradha, S.N. Structural and molecular characteristics of lichenysin and its relationship with surface activity. In Biosurfactants; Springer: New York, NY, USA, 2010; pp. 304–315. [Google Scholar]
- Zhao, H.; Shao, D.; Jiang, C.; Shi, J.; Li, Q.; Huang, Q.; Rajoka, M.S.R.; Yang, H.; Jin, M. Biological activity of lipopeptides from Bacillus. Appl. Microbiol. Biotechnol. 2017, 101, 5951–5960. [Google Scholar] [CrossRef]
- Santos, V.S.V.; Silveira, E.; Pereira, B.B. Toxicity and applications of surfactin for health and environmental biotechnology. J. Toxicol. Environ. Health 2018, 21 Part B, 382–399. [Google Scholar] [CrossRef]
- Thakur, S.; Singh, A.; Sharma, R.; Aurora, R.; Jain, S.K. Biosurfactants as a novel additive in pharmaceutical formulations: Current trends and future implications. Curr. Drug Metab. 2020, 21, 885–901. [Google Scholar] [CrossRef] [PubMed]
- Maget-Dana, R.; Ptak, M. Interactions of surfactin with membrane models. Biophys. J. 1995, 68, 1937–1943. [Google Scholar] [CrossRef]
- Carrillo, C.; Teruel, J.A.; Aranda, F.J.; Ortiz, A. Molecular mechanism of membrane permeabilization by the peptide antibiotic surfactin. BBA—Biomembr. 2003, 1611, 91–97. [Google Scholar] [CrossRef]
- Zhen, C.; Ge, X.F.; Lu, Y.T.; Liu, W.Z. Chemical structure, properties and potential applications of surfactin, as well as advanced strategies for improving its microbial production. AIMS Microbiol. 2023, 9, 195. [Google Scholar] [CrossRef]
- Wang, M.; Noor, S.; Huan, R.; Liu, C.; Li, J.Y.; Shi, Q.; Zhang, Y.J.; Wu, C.; He, H. Comparison of the diversity of cultured and total bacterial communities in marine sediment using culture-dependent and sequencing methods. PeerJ 2020, 8, e10060. [Google Scholar] [CrossRef]
- Marchesi, J.R.; Sato, T.; Weightman, A.J.; Martin, T.A.; Fry, J.C.; Hiom, S.J.; Wade, W.G. Design and Evaluation of Useful Bacterium-Specific PCR Primers That Amplify Genes Coding for Bacterial 16S rRNA. Appl. Environ. Microbiol. 1998, 64, 795–799. [Google Scholar] [CrossRef]
- Kim, O.S.; Cho, Y.J.; Lee, K.; Yoon, S.H.; Kim, M.; Na, H.; Park, S.C.; Jeon, Y.S.; Lee, J.H.; Yi, H.; et al. Introducing EzTaxon-e: A prokaryotic 16s rRNA gene sequence database with phylotypes that represent uncultured species. IJSEM 2012, 62 Pt 3, 716–721. [Google Scholar] [CrossRef]
- Tamura, K.; Nei, M. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. MBE 1993, 10, 512–526. [Google Scholar]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. MBE 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Hall, B.G. Building phylogenetic trees from molecular data with MEGA. MBE 2013, 30, 1229–1235. [Google Scholar] [CrossRef]
- Riyanti Balansa, W.; Liu, Y.; Sharma, A.; Mihajlovic, S.; Hartwig, C.; Leis, B.; Rieuwpassa, F.J.; Ijong, F.G.; Wägele, H.; König, G.M.; et al. Selection of sponge-associated bacteria with high potential for the production of antibacterial compounds. Sci. Rep. 2020, 10, 19614. [Google Scholar]
- Ruhnau, B. Eigenvector-centrality-a node-centrality? Soc. Netw. 2000, 22, 357–365. [Google Scholar] [CrossRef]
- Barry, A.L. Procedure for testing antimicrobial agents in agar media: Theoretical considerations. In Antibiotics in Laboratory Medicine; Willams and Wilkins Company: Baltimore, MD, USA, 1986; pp. 1–26. [Google Scholar]
- Kochva, Z. Characterization of the Bacterial Community Inhabiting Sponges of the Genus Amphimedon and Locating Bacterial Symbionts Which Produce Natural Compounds. Master’s Thesis, Tel Aviv University, Tel Aviv, Israel, 2009. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HPLC of Fraction | Fraction | Weight (mg) | (-)HR ESI MS m/z (%) | Identity |
|---|---|---|---|---|
| M-12 and B-6 | 5 | 10.2 | 1020.6819 (8.3) | 3-oxy-C14-fatty acid1-surfactin |
| 1034.6986 (57.8) | 3-oxy-C15-fatty acid1-surfactin | |||
| 1048.7141 (21.5) | 3-oxy-C16-fatty acid1-surfactin | |||
| 1051.7315 (12.3) | unknown | |||
| 6 | 12.8 | 1005.6753 (8.2) | 3-oxy-C13-fatty acid1-lichensyin | |
| 1034.6908 (20.4) | 3-oxy-C15-fatty acid1-surfactin | |||
| 1048.7079 (60.8) | 3-oxy-C16-fatty acid1-surfactin | |||
| 1065.7411 (10.3) | unknown | |||
| 7 | 7.5 | 1005.6635 (12.7) | 3-oxy-C13-fatty acid1-lichensyin | |
| 1019.6785 (10.0) | 3-oxy-C14-fatty acid1-lichensyin | |||
| 1034.6807 (12.1) | 3-oxy-C15-fatty acid1-surfactin | |||
| 1048.7025 (20.9) | 3-oxy-C16-fatty acid1-surfactin | |||
| 1062.7135 (29.4) | 3-oxy-C17-fatty acid1-surfactin | |||
| 1065.7251 (11.0) | unknown | |||
| 1076.7235 (3.0) | 3-oxy-C18-fatty acid1-surfactin | |||
| 8 | 8.9 | 1005.6625 (5.8) | 3-oxy-C13-fatty acid1-lichensyin | |
| 1019.6788 (33.7) | 3-oxy-C14-fatty acid1-lichensyin | |||
| 1034.6793 (12.0) | 3-oxy-C15-fatty acid1-surfactin | |||
| 1048.6937 (38.3) | 3-oxy-C16-fatty acid1-surfactin | |||
| 1062.6959 (11.0) | 3-oxy-C17-fatty acid1-surfactin | |||
| 9 | 24.2 | 1005.6644 (2.4) | 3-oxy-C13-fatty acid1-lichensyin | |
| 1019.6813 (6.1) | 3-oxy-C14-fatty acid1-lichensyin | |||
| 1033.6973 (71.5) | 3-oxy-C15-fatty acid1-lichensyin | |||
| 1048.6980 (13.9) | 3-oxy-C16-fatty acid1-surfactin | |||
| 1062.7126 (2.4) | 3-oxy-C17-fatty acid1-surfactin | |||
| 10 | 1.0 | 1019.6771 (4.1) | 3-oxy-C14-fatty acid1-lichensyin | |
| 1033.6932 (80.2) | 3-oxy-C15-fatty acid1-lichensyin | |||
| 1048.6935 (12.3) | 3-oxy-C16-fatty acid1-surfactin | |||
| 1062.7090 (2.7) | 3-oxy-C17-fatty acid1-surfactin | |||
| M-11 | 8 * | 4.7 | 1019.6756 (86.9) | 3-oxy-C14-fatty acid1-lichensyin |
| 1033.6915 (13.2) | 3-oxy-C15-fatty acid1-lichensyin |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Itelson, L.; Merav, M.; Haymi, S.; Carmeli, S.; Ilan, M. Diversity and Activity of Bacteria Cultured from a Cup—The Sponge Calyx nicaeensis. Mar. Drugs 2024, 22, 440. https://doi.org/10.3390/md22100440
Itelson L, Merav M, Haymi S, Carmeli S, Ilan M. Diversity and Activity of Bacteria Cultured from a Cup—The Sponge Calyx nicaeensis. Marine Drugs. 2024; 22(10):440. https://doi.org/10.3390/md22100440
Chicago/Turabian StyleItelson, Lynne, Mayan Merav, Shai Haymi, Shmuel Carmeli, and Micha Ilan. 2024. "Diversity and Activity of Bacteria Cultured from a Cup—The Sponge Calyx nicaeensis" Marine Drugs 22, no. 10: 440. https://doi.org/10.3390/md22100440