1. Introduction
Endurance athletes modulate the volume and intensity of their constant training throughout the sports season in order to produce adaptations and achieve their best performance in previously established competitive periods [
1]. This exercise causes stress in the organism that induces important changes in the endocrine system to recover the initial homeostasis [
2].
Cortisol (C) is the main glucocorticoid of the organism, its secretion is produced in the adrenal glands and it is controlled through a negative feedback mechanism by the hypothalamus–pituitary–adrenal axis [
3]. It is a hormone modulated by circadian rhythms, but factors such as mental stress, dehydration or food can alter its production [
4]. In endurance activities, its blood values increase, as its catabolic function contributes to maintaining adequate energy levels through protein degradation, hydrolysis of triglycerides and even adding additional energy from carbohydrates through gluconeogenesis in the liver [
5]. At the end of physical activity, the concentrations of this hormone begin to decrease, and it may take up to 48 h to recover its basal values after maximum effort [
6].
Testosterone (T) is an anabolic hormone that participates in multiple physiological functions, intervenes in muscle protein synthesis, stimulates bone remodeling and erythropoiesis [
7] and regulates the function of lactate transporter proteins thus promoting lactate oxidation as a fuel during exercise [
8]. T is secreted by the Leydig cells in the testicles and its concentration in blood is controlled by the hypothalamic–pituitary–testicular (HPT) axis [
9]. It has been reported that athletes who undergo long-term continuous training may have reduced levels of chronic basal T, status defined as “Exercise-Hipogonadal Male Condition” [
10]. Previous studies have shown that its low levels would be caused by the negative relationship between C and T [
11,
12]. Hackney, Szczepanowska and Viru have hypothesized whether this inhibition would be caused by a dysfunction of the axis at the peripheral level, through direct inhibition of the Leydig cells in the testicles, or at the central level, reducing the release of Luteinizing Hormone (LH) in the pituitary that would affect the production of T at the testicular level [
13].
LH is secreted in the anterior pituitary gland. LH is part of a pathway comprising the hypothalamus, pituitary gland, and gonads [
14]. The release of LH is stimulated by gonadotropin-releasing hormone (GnRH) [
15]. After acute physical exercise, LH usually decreases [
16,
17]. During the training phases, a reduction in LH secretion was found in runners [
18].
The T/C ratio is a variable that relates to the anabolic/catabolic balance in athletes and is widely used for monitoring and evaluating the body’s response to chronic exercise-induced stress [
19]. Authors such as Meeusen et al. [
20] think that the ratio cannot be used as a means of control since they reported studies where decreases of 30% of the ratio did not always worsen the athletes’ performance.
Another important hormone involved during physical exercise is insulin (I), related to energy balance and blood glucose control [
21]. Horton, Grunwald, Lavely and Donahoo reported a decrease in plasma I concentration during exercise, followed by an increase during the hours after exercise to favor glycogen repletion, a decrease in carbohydrate oxidation, and an increase in fat oxidation [
22]. Aerobic endurance athletes have lower baseline values and higher insulin sensitivity than sedentary subjects to support fatty acid oxidation [
23].
In summary, hormones have been defined as important mediators in the body’s response and adaptations to exercise-induced stress. Their acute responses to different stimuli and their modifications over short periods have been extensively investigated [
24,
25]. However, few studies have shown hormonal changes in high-level endurance runners during a sports season, so the aim of this study was to determine the baseline values of T, C, LH and I and their changes throughout a sports season where training loads are modulated to obtain several peaks of performance.
3. Results
Table 2 shows ergospirometric and body composition variables in the runners during the season. In our study, VO
2 max, VT
2, RER, heart rate maximum (HRM), fat mass, fat-free mass and ∑6 skinfolds did not show significant changes during the season. Weight suffered significant decreases (
p < 0.05) at 6 and 9 months compared with initial values.
Nutritional intake of the athletes during the season is shown in
Table 3. The athletes followed a diet using established energy and macronutrient guidelines for adequate athletic performance [
32]. Energy availability (EA) is defined as the amount of energy intake (kcal day
−1) −exercise energy expenditure (kcal day
−1)] /FFM. An appropriate energy balance equates to ≥45 calories per day per kg of FFM (kcal/kg/FFM/d) [
33].
Plasmatic concentrations of hormones are shown in
Table 4.
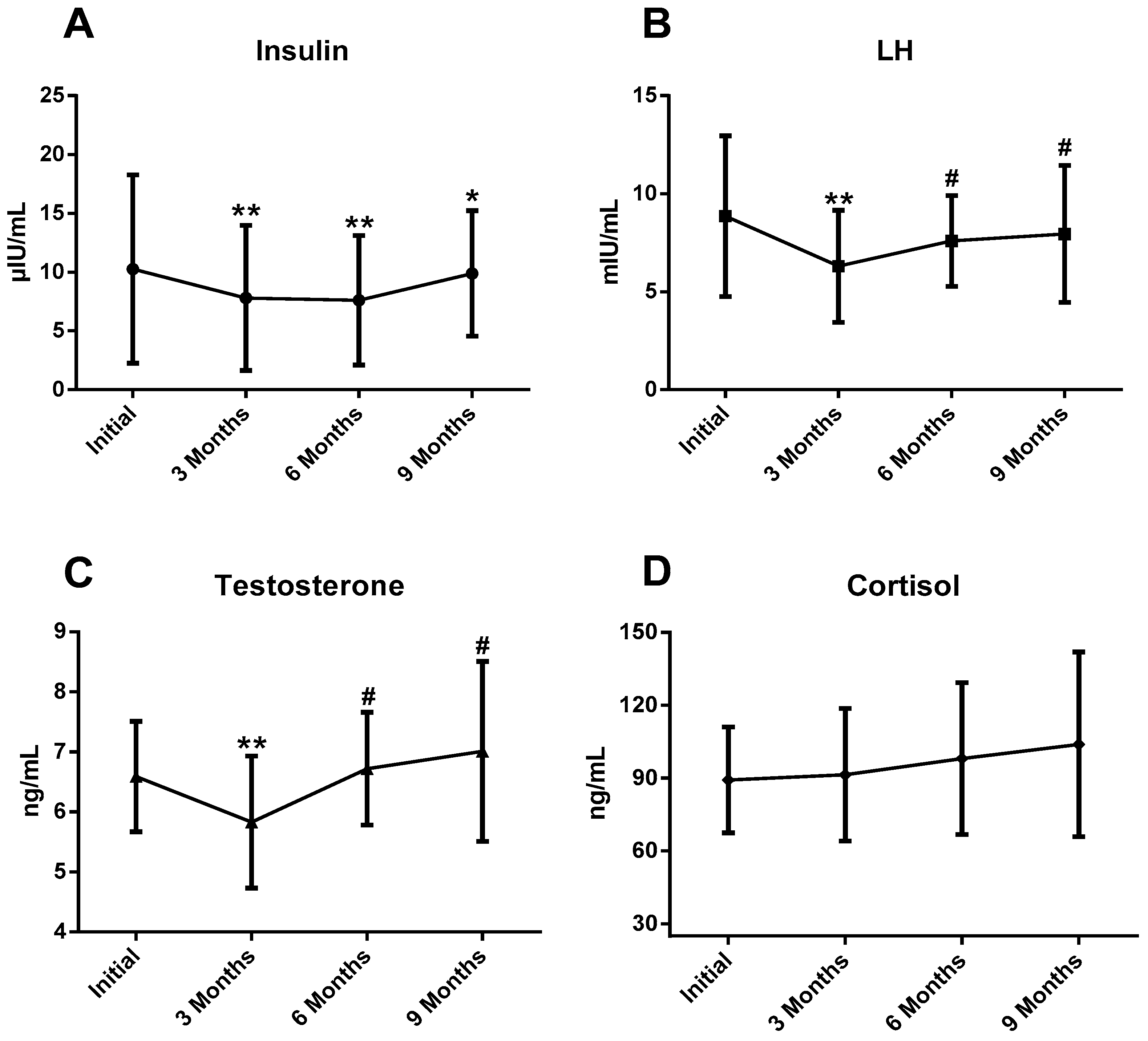
Analysis of the data revealed changes in plasmatic concentrations of total T, LH and I during the sports season. T and LH suffered very significant decreases (
p < 0.01) at 3 months compared with the initial test, and an increase (
p < 0.05) at 6 and 9 months compared with 3 months. I concentrations were significantly lower at 3, 6 (
p < 0.01) and 9 (
p < 0.05) months compared with the initial test (
Figure 2). There were no statistical differences in plasmatic C concentration and T/C ratio during the athletic season.
Simple linear regressions between the plasmatic hormones and km trained are shown in
Table 5 and
Table 6. Plasmatic concentrations of I and C were inversely (r = 0.363; β = −0.577;
p = 0.017) and positively (r = 0.202; β = 0.310;
p = 0.043) associated, respectively, with the amount of km trained per week.
There was a significant association between km trained at a higher intensity than VT2 and I (r = −0.580; β = −0.442; p = 0.000).
4. Discussion
The purpose of our longitudinal study was to observe the changes in plasma basal concentrations of LH, T, C and I in high-level endurance runners, as well as the possible changes that occur during a sports season in relation to the training performed.
VO
2 max and VT
2 did not show significant changes during the season in our athletes. High values of VO
2 max are required in endurance athletes, although it is not a determinant variable among homogeneous groups [
34]. Body composition and running economy are other variables related to performance in endurance runners [
35,
36].
The findings of this research agree with those observed in other studies in which they found that training throughout a sports season produces adaptations in the endocrine system with the aim of improving the athletes’ performance [
2]. The basal concentrations of the different hormones in the study showed significant changes during the sports season although they remained within the normal reference values for humans [
37,
38].
C did not suffer significant changes during the season, since athletes were examined in the laboratory without having carried out intense exercise the previous days. Mäestu, Jürimäe and Jürimäe reported that C did not change when training volume was increased [
39]. In our study, C showed a positive association with the volume of km performed per week. Purge, Jürimäe and Jürimäe also observed a significant relation between C and mean training volume in elite male rowers [
40]. It is known that during long-term aerobic exercise hypercortisolemia occurs which contributes to maintaining adequate energy levels during training [
9]. The C prevents the re-esterification of fatty acids released by the catecholamine-induced lipolysis [
41]. The activation of catabolic processes is an essential tool for adaptation in high-stress conditions [
42].
Cortisol is usually elevated in energy-deficient conditions. Increases in cortisol circulation have been observed in studies of severe caloric restriction or fasting [
43]. As mentioned above, during endurance exercise C concentrations increase, and previous studies have reported an inhibitory effect of C on T synthesis [
9,
44] that could interfere with athletes’ recovery and performance, since it participates in protein synthesis and erythropoiesis [
45], and could even negatively affect their health due to low bone mineral density and infertility [
10].
It has been reported that chronic endurance training may have negative effects on the basal concentration of T, which leads to chronic low levels of this hormone as a consequence of the accumulation of aerobic training over years [
46]. In our research, athletes suffered a significant decrease in basal T levels at 3 months accompanied by a significant decrease in LH, a fact that would indicate that there is an alteration of the HPT axis at the central level, decreasing the secretion of GnRH from the hypothalamus that would affect the release of LH and, consequently, the Leydig cells would not be stimulated for T synthesis [
47,
48]. This could be caused by the fact that in this period the runners trained the highest volume of km per week of the entire season, which forced them to carry out longer training sessions with less recovery between sessions that would promote maintaining high concentrations of C during this phase. MacConnie [
48] reported a decrease in LH pulse frequency in highly trained runners. Several studies have reported a negative relationship between T concentrations and high volumes of aerobic training, where the HPT axis is altered in runners who performed more than 100 km/week as occurred with our athletes [
49,
50]. Flynn et al., observed a decrease in T after the training volume had been increased by 88% for two weeks in swimmers [
51].
However, in a recent review suggesting that running mileage alone is not enough to predict the low T concentrations [
52], it was proposed that the alterations in the endocrine–reproductive hormonal system observed in endurance runners are related to the development of low energy availability (LEA) [
33]. In healthy and active women, it has been established that an adequate energy intake is ≥45 kcal/kg FFM [
53]; whether intake in men is similar is currently under debate [
52]. In our study, runners reported an energy intake lower than 45 kcal/kg FFM initially and especially at 3 months, a period when the runners ran more km/week, and basal T and LH concentrations were the lowest of the season. De Souza [
53] concluded that it is difficult to consume the energy required by athletes who perform chronic strenuous exercise, resulting in an energy deficit that causes alterations in the hypothalamic- pituitary- gonadal axis.
Another factor that could contribute to the decrease in basal T values is the variation in the annual circadian rhythms that this hormone suffers from as a consequence of exposure to the sun, thus, higher peak concentrations have been verified during the summer months, and lower levels during the winter as occurred in our research, since the third month of the study corresponded to the month of January in our region [
54]. Low circulating vitamin D concentrations have been associated with a lower total T concentration [
55]. Lombardi et al. [
56] confirmed the importance of sun exposure and solar irradiance in the vitamin D and T concentrations in professional soccer players during two sports seasons, also, significant correlations between vitamin D and T were reported.
In the second part of the season, at 6 and 9 months, we observed an increase in the basal concentrations of T and LH. It seems that hormonal changes as a consequence of LEA are reversible when the subjects have adequate energy available [
57], as occurred in the runners of our study when they performed less km/week and they had adequate energy intake. In addition, the target competitions to be carried out in these periods were shorter distances and higher intensities (maximum 16 min), where the athletes trained less km per week and with greater intensity (more weekly sessions >VT
2). Previous studies have reported that high-intensity training produces an increase in T [
11,
58], which could be a consequence of the large reduction in night time C concentrations that occur during these training sessions compared to aerobic training [
59]. T could play an essential role in muscle metabolism during the tapering phase [
39,
60]. T seems to increase the ability of the muscle to refill its glycogen stores through increased activity of muscle glycogen synthetase [
61].
This would be very important for adequate regeneration after prolonged exercise and during intensive training periods [
62]. Other factors mentioned above are important, such as the annual variation suffered by this hormone; the measures taken at 6 and 9 months correspond to the months of April and July, respectively, where there are more hours of sun exposure and solar irradiance, during those periods which could have favored the increase in vitamin D concentrations and basal T concentrations [
63].
As for the I, there was a decrease in its basal values with respect to the initial ones throughout the season, more visible in the periods of greater volume of km trained, also, remarkable negative associations were revealed with the number of km trained per week. Jürimäe, Purge and Jürimäe [
64] reported an inverse correlation between I and training volume in elite rowers (r = −0.399,
p < 0.05). Other study reported a significantly lowered maximal exercise-induced level of I [
41]. Reduced insulin levels have been observed in energy-deficient athletes [
43].
During prolonged training, the I level decreases because the catecholamine increase inhibits the I secretion [
65]. This phenomenon could favor glucose homeostasis with increased glucose availability for the central nervous system [
41]. It has been widely reported that training improves insulin sensitivity [
66], which can be considered a positive adaptation produced in runners to enhance the use of fatty acids as fuel [
23]. Previous studies with athletes have reported a decrease in the plasma I concentration during exercise, followed by an increase during the hours after the end of exercise, in that time there is a decrease in the oxidation of carbohydrates and an increase in the oxidation of fats to favor the repletion of glycogen [
22].
The small number of participants and the absence of control of I sensitivity are limitations of the present study. Basal vitamin D concentrations could not be analyzed during the season to observe its circannual rhythm and the possible relationship with the hours of sun and its irradiation. Using Dual-energy X-ray absorptiometry (DXA) or Bioelectrical Impedance Analysis (BIA) are methods that could provide more accurate data on the body composition of subjects (fat mass and fat-free mass) that would have allowed a better assessment of changes in muscle mass and its relationship with dietary intake and hormonal concentrations. Blood samples were not drawn for lactate collection during the treadmill running test.

 ,
, 





{kind=link}
{kind=link}