1. Introduction
The forest canopy is an important interface connecting the forest ecosystem and external environment, providing complex microhabitats for diverse organisms [
1,
2]. As a key feature of the forest canopy, maximum canopy height (Hmax) represents the ability of individual trees to intercept resources (i.e., light and precipitation) and has been widely used in the field of ecology [
3,
4]. Previous studies found that the Hmax pattern exhibits strong spatial geographical variability, and such variation is often used to indicate the functioning and dynamics of the forest ecosystem [
5,
6]. Nevertheless, the drivers of Hmax variation are still debated [
7,
8,
9,
10].
Various theories have been proposed to explain the variability of maximum canopy height [
7,
8,
9,
10,
11,
12]. Among them, the “hydraulic limitation hypothesis” [
13] and the “energy limitation hypothesis” [
14] are two widely discussed theories for explaining large-scale Hmax patterns [
15]. The “hydraulic limitation hypothesis” suggests that xylem water potential and available water can limit the Hmax of the canopy [
16]. In contrast, the “energy limitation hypothesis” holds that energy, either sun radiation or temperature, is an important factor regulating canopy height variation [
17]. Although many studies have been conducted to evaluate the ability of these two hypotheses to explain canopy height variation, the results are controversial because of the different data sources. For example, Moles et al. (2009) pointed out that water availability may govern global plant height distributions [
18], while Larjavaara et al. (2014) found that energy was the most significant determinant of the occurrence of the tallest individual [
14]. With light detection and ranging (LiDAR) remote sensing data, Klein et al. (2015) [
19] and Tao et al. (2016) [
20] validated the role of the hydraulic limitation hypothesis in explaining the global canopy height. The above studies show that water and energy factors shape the Hmax pattern on a continental and global scale. However, most previous researchers utilized continuous remote sensing data with a resolution greater than 1 km [
19,
20]. This may underestimate the variability in canopy height on a smaller scale and make it impossible to accurately quantify its driving forces. Therefore, local-scale analysis has the potential to provide new insights into our understanding of canopy height variation.
At the local scale, the light and water availability difference caused by topography can determine canopy height variation by controlling the spatial distribution of large individuals [
21,
22]. For example, in a valley, a lack of light will intensify the light competition among plants, causing individuals to grow taller due to the higher water availability. In contrast, on slopes, abundant light resources allow plants to reduce their investment in height and increase their investment in water absorption and transport to be competitive for water [
23]. Although both the hydraulic limitation hypothesis and the alternative energy limitation hypothesis can be used to explain the canopy height variation at the local scale, their influence would be different across forest types. For example, in certain ecosystems, such as humid temperate forests, energy can be more important than water availability due to the seasonal fluctuation in temperature and light radiation. A global analysis also suggested that canopy height variation in high-latitude areas (e.g., boreal forests) is subject to the energy limitation hypothesis, whereas lower latitude areas (e.g., tropical forests) are subject to the hydraulic limitation hypothesis [
15]. However, it remains unclear whether there is a similar directional change for the drivers of local canopy height variation from high- to low-latitude zones.
In this study, we collected airborne LiDAR data from eight dynamic forest plots, with an area range of 20–30 ha and locations ranging from temperate to tropical zones in China. We extracted the maximum canopy height (Hmax) in a 20 × 20 m quadrat and calculated the topographic wetness index (TWI) and digital terrain model-derived insolation (DTMI) to represent water and energy conditions at the local scale. We aimed to quantify the influence of water and energy on the Hmax variation at local scales and identify whether the main drivers of canopy height variation showed a directional change from temperate to tropical forests. We predicted that water limitation would play a greater role in the low-latitude forest plots, where water is more limiting to trees than is energy. In contrast, energy limitation will be more important in high-latitude forest plots because of the greater seasonal fluctuation in temperature and light radiation in these regions. Our results provide insights into the role of the “water limitation hypothesis” and the “energy limitation hypothesis” in explaining canopy height variation in forest communities.
3. Results
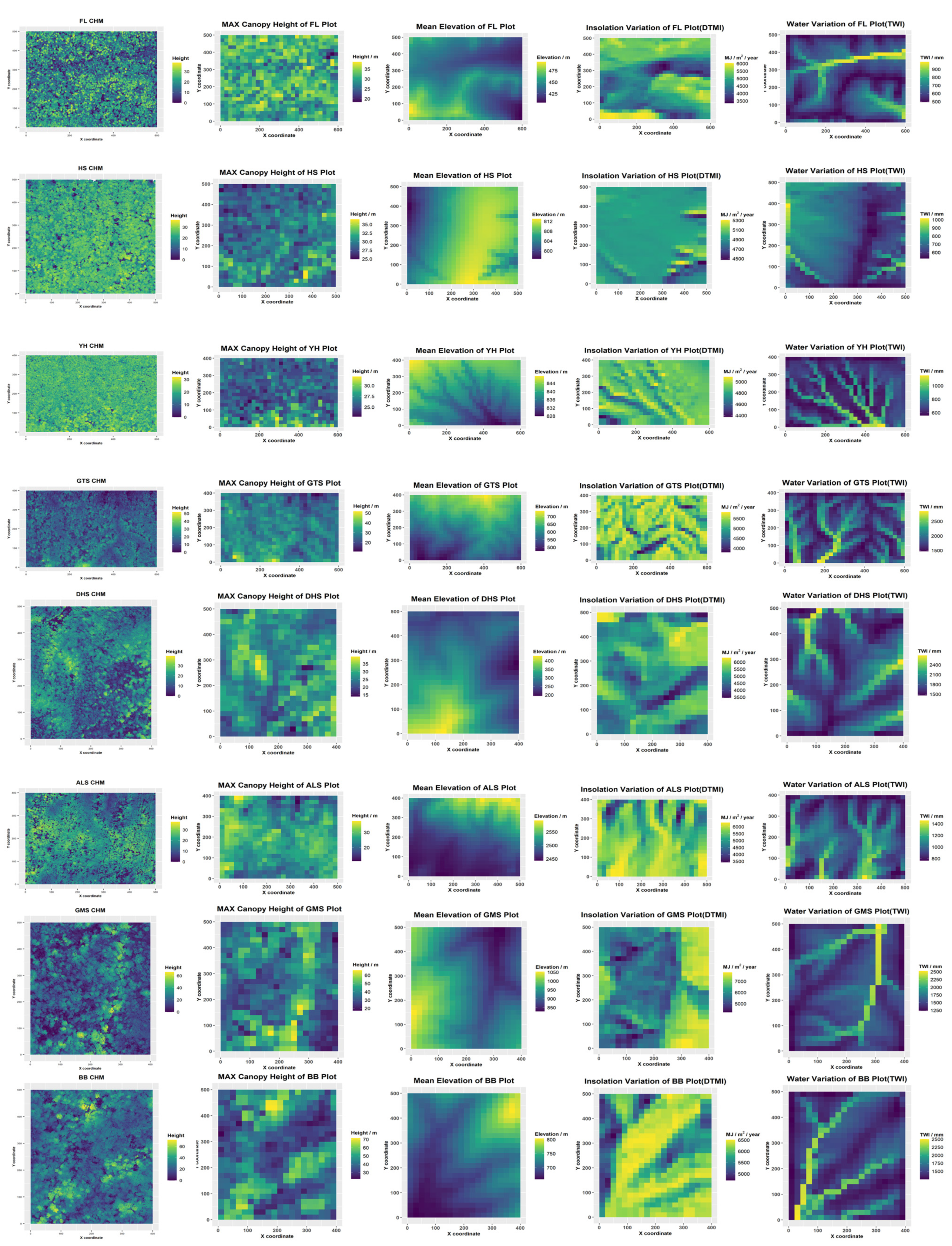
The quadrat-level maximum canopy height ranged from 10 to 70 m across all forest plots (
Table 2). The GMS and BB plots had the greatest quadrat maximum canopy height of around 70 m, while the mean Hmax of the eight plots was approximately 40 m. The quadrat canopy height of the HS and YH plots was highly centralized, compared with that of the six other plots. More information such as CHM and DEM of each plot is in
Figure A1.
The LMM included plot as the random intercept and the slope was the best model, with the lowest AIC and highest R-square value of 0.633 (
Table 3). TWI was highly significantly positively related to all quadrat maximum canopy heights (coefficient = 4.21,
p < 0.01), and the DTMI was weakly and non-significantly negatively related to the Hmax (coefficient = −0.35,
p > 0.1).
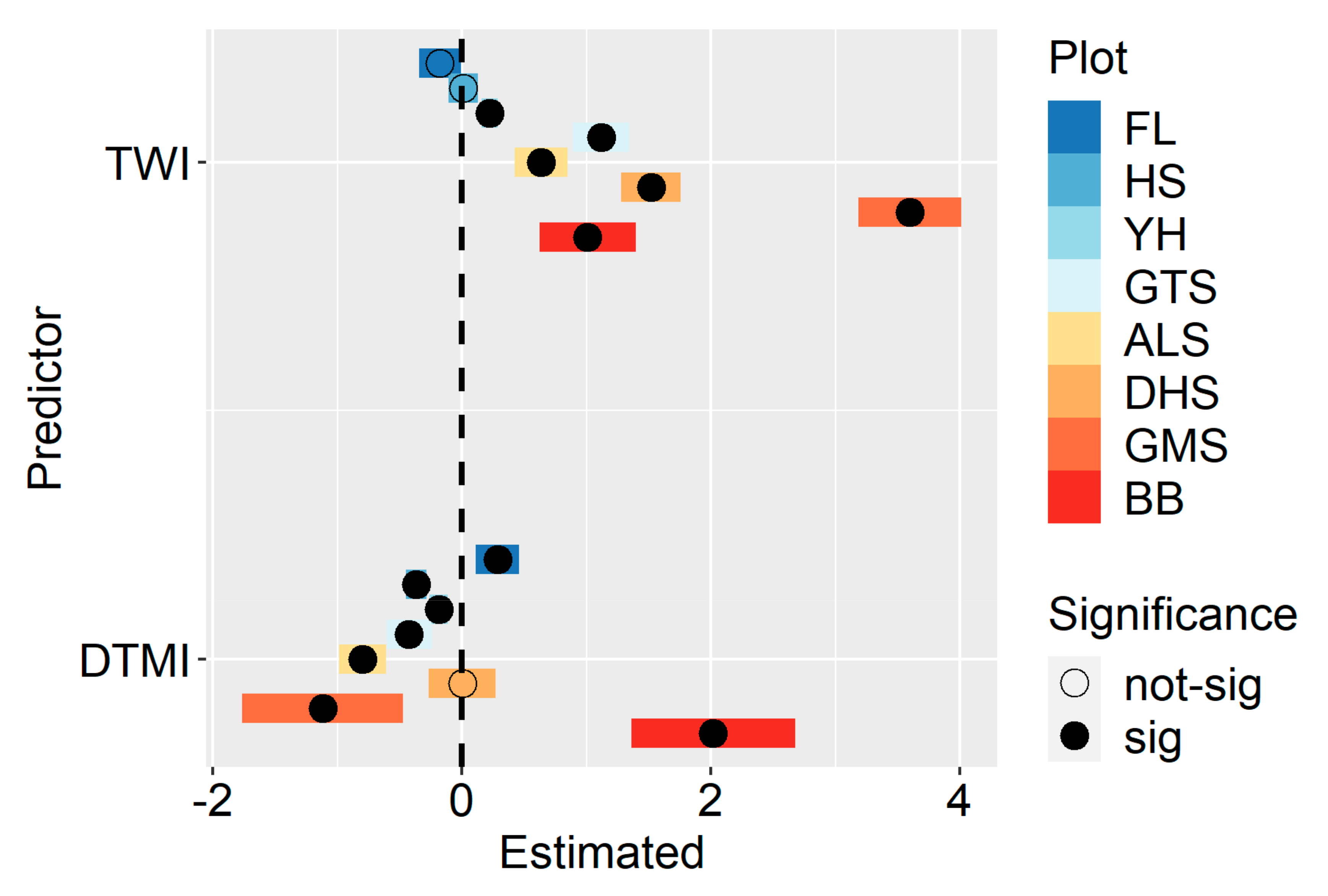
We found that TWI showed a significant, positive influence except in the FL and HS plots. DTMI had a significant influence except in the DHS plot. The effect of DTMI was positive in the FL and BB plots but negative in the other five plots. Interestingly, we found that the effect of TWI changed from a non-significant, negative effect to a significant, positive effect when changing locations from temperate to tropical forests. The effect of DTMI showed a similar directional change pattern from positive to negative when the forest types changed from temperate to tropical forests, except in the BB forest plots (
Figure 2). The detailed information is in
Table A2.
We found that DTMI had higher importance in temperate forests, while TWI had a stronger explanatory power in subtropical and tropical forests (
Figure 3). The DTMI accounted for more than 90% of the total variation in the two temperate forest plots (i.e., FL and HS plots), while DTMI explained less than 10% of the variation in the subtropical and tropical forest plots (except for ALS). Unexpectedly, we found TWI was slightly more important than DTMI in the YH plot, which is a typical secondary temperate forest. The detailed information is in
Table A3.
4. Discussion
Understanding the patterns and determinants of forest canopy height variation can provide critical insights into the drivers of ecosystem functions [
33]. Despite an extensive body of research on canopy height variation, few studies have focused on the patterns and determinants of canopy height variation at the local scale. In this study, based on near-surface LiDAR data from eight large permanent forest plots in China, we accurately measured canopy height variation at the local scale and quantified the relative contributions of energy and water limitations to canopy height variation from temperate to tropical forests.
In this study, we found that TWI, rather than DTMI, was the major driver of maximum canopy height variation when we combined all plots, supporting the hydraulic limitation hypothesis. Previous studies demonstrated that canopy height variation is mainly affected by water limitation at both global and regional scales [
19,
20,
34]. Our results further highlighted the dominant role of water availability in controlling forest canopy height at the local scale. The positive relationship between TWI and Hmax suggested that higher water availability can increase the maximum canopy height. Higher maximum canopy height and vertical heterogeneity could lead to greater use of niche space for light and provide multidimensional space that can be occupied by different species [
33]. Therefore, our results suggest that ensuring the water availability of plants, especially large trees, is an important prerequisite for effectively maintaining biodiversity at the local scale. The altitude of the ALS plot is 2509 m, which causes the water vapor in the cloud to act as an important supplement of water, thus alleviating the effect of water limitation on the canopy height of the ALS plot. In the context of global climate change, more frequent and devastating droughts have reduced the soil water content, streamflow, and groundwater recharge [
35,
36]. Further studies are required to investigate how drought influences biodiversity maintenance by altering the complexity of the canopy structure.
When we focused on individual plots, we found that the TWI had limited influence on the canopy height variation in the temperate forest plots. This is because trees grow mainly in the summer when precipitation is high in the temperate forests of East Asia. The abundant water supplies contribute to the weak relationship between large trees and water availability. We also found that the positive effect of TWI increased significantly from temperate to tropical forests, suggesting that the spatial distribution of large trees is more related to the local water variability in subtropical and tropical forests. This result is similar to that found in previous studies, i.e., that water availability can determine plant species distribution in tropical forests [
37,
38]. Species with different heights can develop corresponding adaptive strategies to water availability through hydraulic regulation [
39]. Therefore, trees growing in areas with higher water availability can invest more resources into height growth rather than hydraulic conductivity [
40]. Furthermore, two studies have found that species co-existing within a community often have specific habitat preferences [
41,
42]. Our study supported this view by identifying the relationship between tree height and water availability; that is, species with greater height tend to grow in areas with high water availability.
Although the relationship between DTMI and canopy height variation was non-significant when we combined all plots, we found that DTMI had a significant influence on canopy height variation in most individual forest plots. This result highlights the necessity of local-scale analysis for our understanding of canopy height variation. Most importantly, we found that DTMI had higher importance in temperate forests than in subtropical and tropical forests. This finding supports the idea that energy is an important factor regulating canopy height variation in high-latitude forests because of the low temperature and high heterogeneity of solar radiation [
26,
43]. However, different from previous results supporting the energy limitation hypothesis, our study demonstrated that the effect of energy on canopy height variation can be both positive and negative in temperate forests. The first possible reason for this finding may stem from the geographical characteristics of temperate forests. Temperate forests are located near the middle of a full latitudinal gradient from tropical forests to boreal forests and contain flora from temperate, subtropical, and subarctic zones. Global analysis suggests that the drivers of canopy height variation change from energy to water when the forest type shifts from boreal to tropical [
15]. Therefore, temperate forests are in a transition zone from energy-limited to water-limited geography, and the complex flora composition makes it possible for forest canopy heights to respond differently to energy and water limitations. The second possible explanation is the local habitat heterogeneity. We found a large elevation difference in the FL plot. Due to the undulating terrain, the energy at lower elevations is usually lower than that at higher elevations. As a result, the vertical growth of plants can benefit from higher energy availability when growing in higher elevation areas, resulting in a positive relationship between canopy height and energy availability. In contrast, the HS plot had a relatively flat terrain and a relatively uniform and low energy distribution. Thus, plants grown in relatively energy-poor areas can mitigate the adverse effects of the energy deficit by increasing their height, and then their canopy heights are greater than those grown in energy-sufficient areas. This may explain why we observed an inverse relationship between canopy height and energy availability.


 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}