
Future Range Shifts Suggest That the Six-Spined Spruce Bark Beetle Might Pose a Greater Threat to Norway Spruce in Europe than the Eight-Spined Spruce Bark Beetle
Abstract
:1. Introduction
2. Materials and Methods
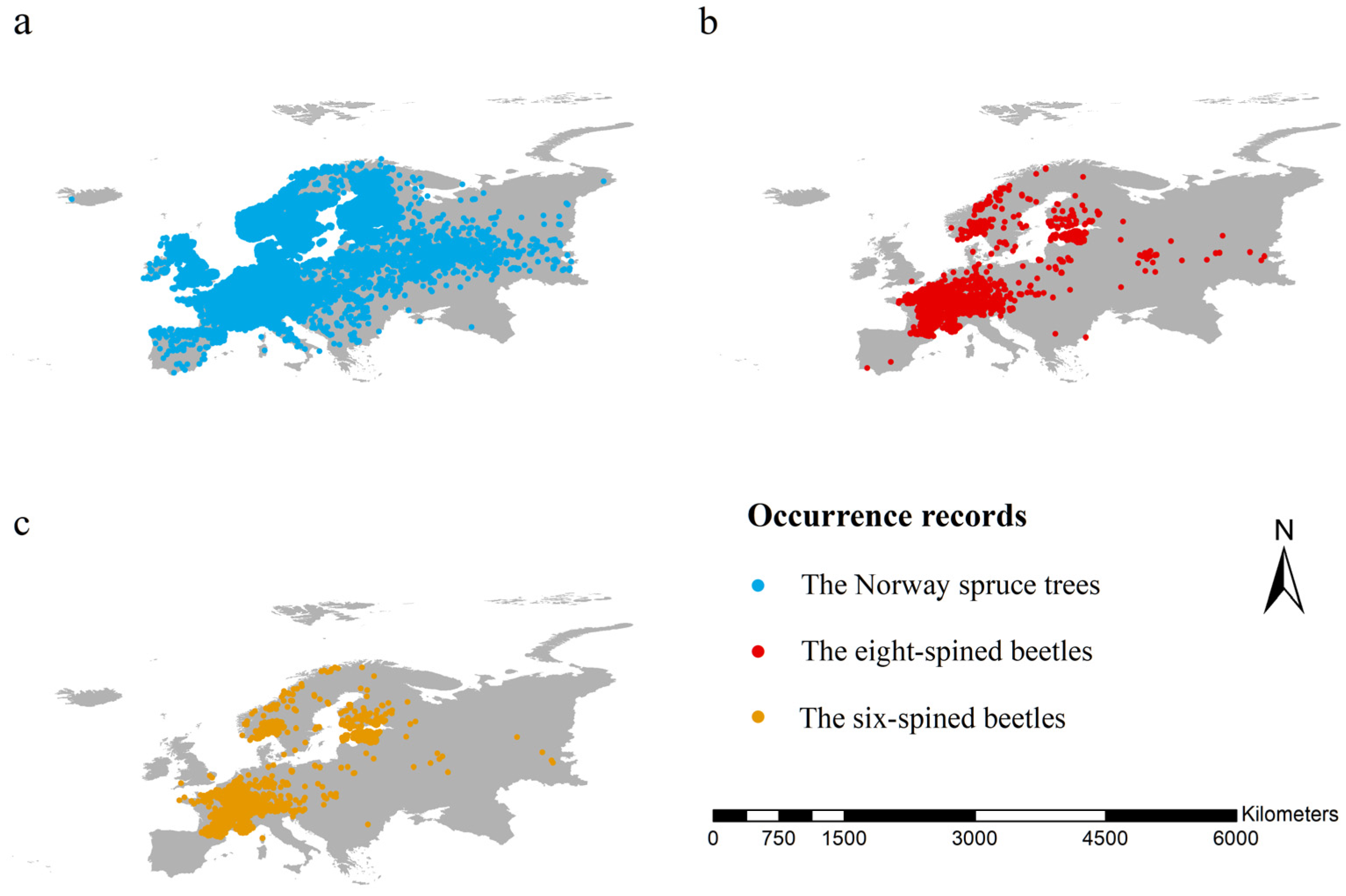
2.1. Occurrence Records
2.2. Predictors in the SDMs
2.3. Projecting the Potential Ranges of Pests and Host
2.3.1. Selection of Predictors
2.3.2. Projecting the Potential Ranges of Pests and Their Host
2.3.3. Assessment of Model Performance
2.4. Range Shifts between Current Conditions and Future Change Scenarios
2.5. Range Overlap between the Two Pests and Norway Spruce
3. Results
3.1. Assessment of the SDMs
3.2. Importance of the Predictors
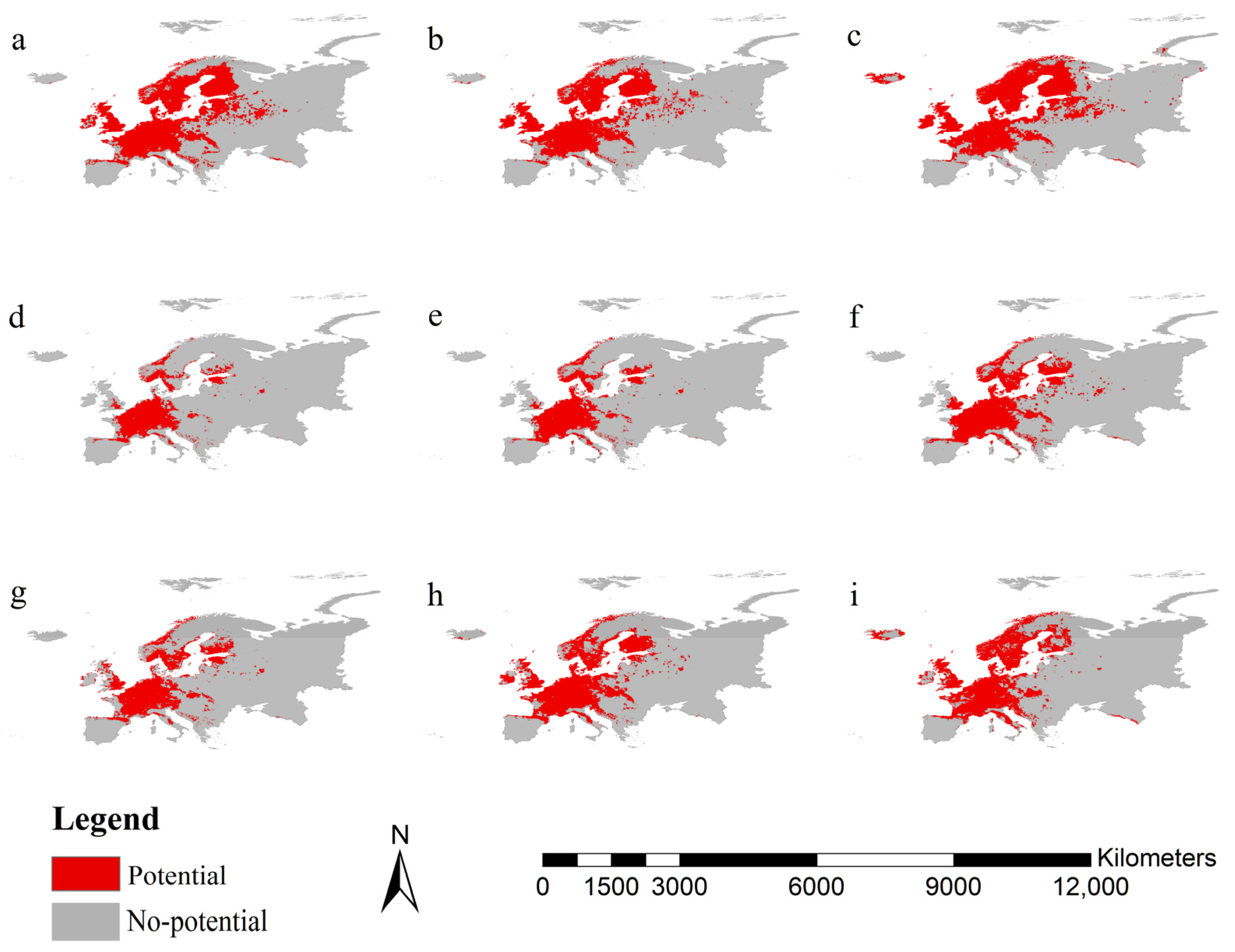
3.3. Potential Ranges of Norway Spruces and the Two Pests
3.4. Shifts in Potential Ranges
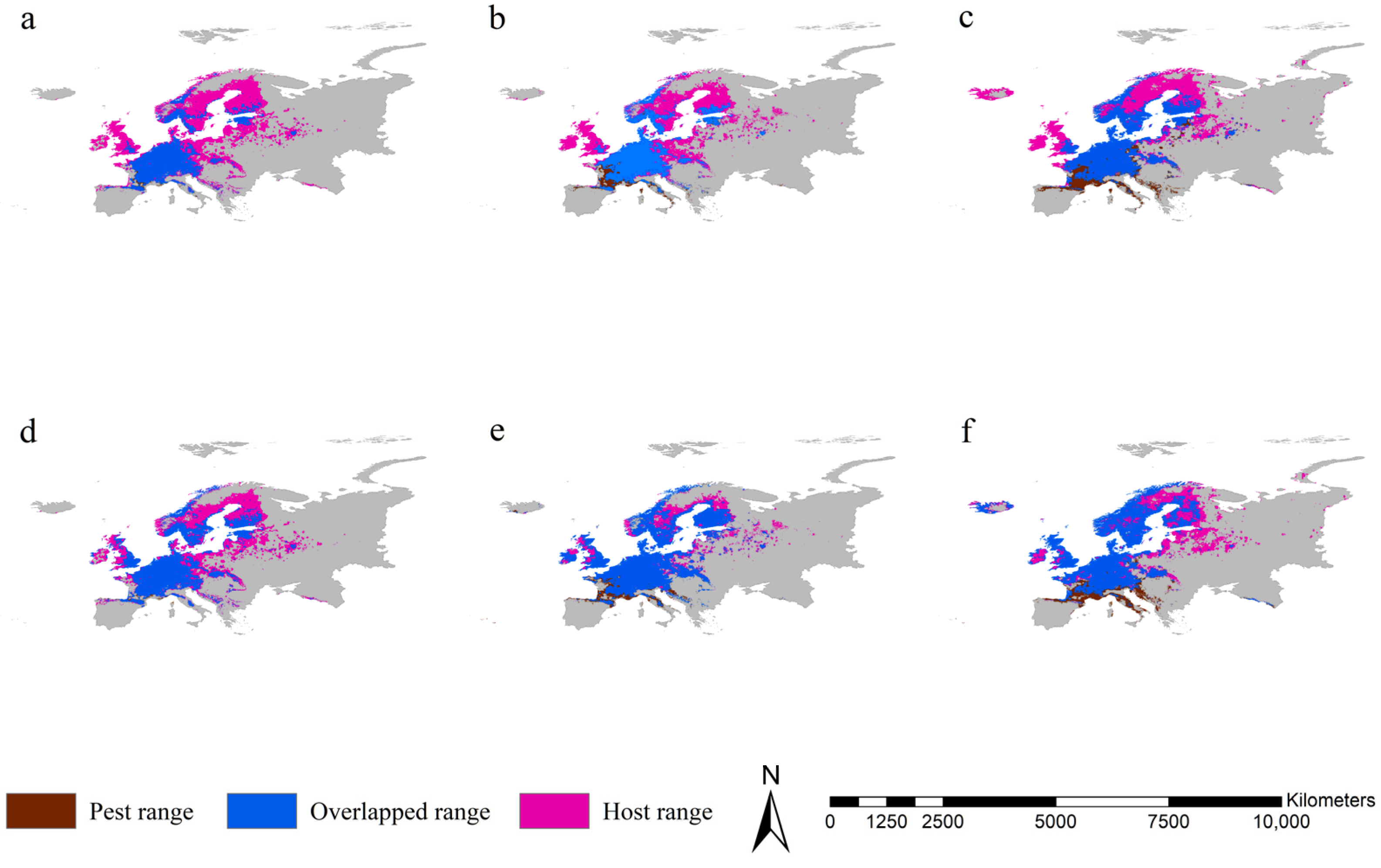
3.5. Range Overlap between the Two Pests and Norway Spruce
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- MacFarlane, D.W.; Meyer, S.P. Characteristics and distribution of potential ash tree hosts for emerald ash borer. For. Ecol. Manag. 2005, 213, 15–24. [Google Scholar] [CrossRef]
- Looney, C.; Sheehan, K.; Bai, B.; Flowers, R.; Kohler, G.; LaGasa, E.; McKorney, A.E.; Smith, D.R. The distribution of a potential new forest pest, Monsoma pulveratum (Hymenoptera: Tenthredinidae), in the pacific northwest states. Northwest Sci. 2012, 86, 342–345. [Google Scholar] [CrossRef]
- Liebhold, A.M.; McCullough, D.G.; Blackburn, L.M.; Frankel, S.J.; Holle, B.V.; Aukema, J.E. A highly aggregated geographical distribution of forest pest invasions in the USA. Divers. Distrib. 2013, 19, 1208–1216. [Google Scholar] [CrossRef]
- Srivastava, V.; Griess, V.C.; Keena, M.A. assessing the potential distribution of Asian gypsy moth in Canada: A comparison of two methodological approaches. Sci. Rep. 2020, 10, 22. [Google Scholar] [CrossRef]
- Nie, P.X.; Yang, R.J.; Cao, R.Y.; Hu, X.K.; Feng, J.M. Niche and range shifts of the fall webworm (Hyphantria cunea Dury) in Europe imply its huge invasion potential in the future. Insects 2023, 14, 316. [Google Scholar] [CrossRef]
- Hof, A.R.; Svahlin, A. The potential effect of climate change on the geographical distribution of insect pest species in the Swedish boreal forest. Scand. J. For. Res. 2016, 3, 29–39. [Google Scholar] [CrossRef]
- Guan, J.; Li, M.; Ju, X.; Lin, J.; Wu, J.; Zheng, J. The potential habitat of desert locusts is contracting: Predictions under climate change scenarios. PeerJ 2021, 9, e12311. [Google Scholar] [CrossRef]
- Parkash, R.; Ramniwas, S.; Kajla, B. Climate warming mediates range shift of two differentially adapted stenothermal Drosophila species in the Western Himalayas. J. Asia-Pac. Entomol. 2013, 16, 147–153. [Google Scholar] [CrossRef]
- Lenoir, J.; Svenning, J.C. Climate-related range shifts—A global multidimensional synthesis and new research directions. Ecography 2015, 38, 15–28. [Google Scholar] [CrossRef]
- Perktas, U.; Elverici, C. Climate-driven range shifts of the sharp-tailed grouse Tympanuchus phasianellus. Acta Ornithol. 2019, 54, 213–222. [Google Scholar] [CrossRef]
- Jiang, Z.W.; Ma, L.; Mi, C.R.; Tao, S.A.; Guo, F.Y.; Du, W.G. Distinct responses and range shifts of lizard populations across an elevational gradient under climate change. Glob. Chang. Biol. 2023, 29, 2669–2680. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.M.; Wang, J.M.; Hu, X.K.; Feng, J.M. Land-use change drives present and future distributions of fall armyworm, Spodoptera frugiperda (JE Smith) (Lepidoptera: Noctuidae). Sci. Total Environ. 2020, 706, 135872. [Google Scholar] [CrossRef] [PubMed]
- Gong, X.; Chen, Y.J.; Wang, T.; Jiang, X.F.; Hu, X.K.; Feng, J.M. Double-edged effects of climate change on plant invasions: Ecological niche modeling global distributions of two invasive alien plants. Sci. Total Environ. 2020, 740, 139933. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Gao, Z.; Wang, L.Q.; Xiao, L.J.; Dong, N.; Wu, H.J.; Li, S. Projecting the potential distribution of ticks in China under climate and land use change. Int. J. Parasitol. 2021, 51, 749–759. [Google Scholar] [CrossRef] [PubMed]
- Balint, M.; Domisch, S.; Engelhardt, C.H.M.; Haase, P.; Lehrian, S.; Sauer, J. Cryptic biodiversity loss linked to global climate change. Nat. Clim. Chang. 2011, 1, 313–318. [Google Scholar] [CrossRef]
- Liu, B.Y.; Jiao, Z.H.; Ma, J.; Gao, X.; Xiao, J.H.; Hayat, M.A. Modelling the potential distribution of arbovirus vector Aedes aegypti under current and future climate scenarios in Taiwan, China. Pest Manag. Sci. 2019, 75, 3076–3083. [Google Scholar] [CrossRef]
- Sirami, C.; Caplat, P.; Popy, S.; Clamens, A.; Arlettaz, R.; Jiguet, F. Impacts of global change on species distributions: Obstacles and solutions to integrate climate and land use. Glob. Ecol. Biogeogr. 2017, 26, 385–394. [Google Scholar] [CrossRef]
- Dempster, J.P.; Pollard, E. Fluctuations in resource availability and insect populations. Oecologia 1981, 50, 412–416. [Google Scholar] [CrossRef] [PubMed]
- Ylioja, T.; Roininen, H.; Ayres, M.P.; Rousi, M.; Price, P.W. Host-driven population dynamics in an herbivorous insect. Proc. Natl. Acad. Sci. USA 1999, 96, 10735–10740. [Google Scholar] [CrossRef]
- Halsch, C.A.; Shapiro, A.M.; Thorne, J.H.; Waetjen, D.P.; Forister, M.L. A winner in the Anthropocene: Changing host plant distribution explains geographical range expansion in the gulf fritillary butterfly. Ecol. Entomol. 2020, 45, 652–662. [Google Scholar] [CrossRef]
- Silva, D.P.; Varela, S.; Nemésio, A.; De Marco, P. Adding biotic interactions into paleodistribution models: A host-cleptoparasite complex of neotropical orchid bees. PLoS ONE 2015, 10, e0129890. [Google Scholar] [CrossRef]
- Hanspach, J.; Schweiger, O.; Kuhn, I.; Plattner, M.; Pearman, P.B.; Zimmermann, N.E.; Settele, J. Host plant availability potentially limits butterfly distributions under cold environmental conditions. Ecography 2014, 37, 301–308. [Google Scholar] [CrossRef]
- Christiansen, E.; Bakke, A. The spruce bark beetle of Eurasia. In Plenum Publishing Corporation; Berryman, A.A., Ed.; Dynamics of Forest Insect Populations: New York, NY, USA, 1988; pp. 479–503. [Google Scholar]
- Pfeffer, A. Zentral-und Westpaläarktische Borken-und Kernkäfer (Coleoptera: Scolytidae, Platypodidae); Pro Entomologia, Naturhistorisches Museum Basel: Basel, Switzerland, 1995; 310p. [Google Scholar]
- Day, K.R.; Leather, S.R. Threats to forestry by insect pests in Europe. In Forests and Insects; Watt, A.D., Stork, N.E., Hunter, M.D., Eds.; Chapman & Hall: London, UK, 1997; pp. 177–205. [Google Scholar]
- Six, D.L.; Bracewell, R. Dendroctonus. In Bark Beetles: Biology and Ecology of Native and Invasive Species; Vega, F.E., Hofstetter, R.W., Eds.; Academic Press: New York, NY, USA, 2015; pp. 305–350. [Google Scholar]
- Muller, M.; Olsson, P.O.; Eklundh, L.; Jamali, S.; Ardo, J. Features predisposing forest to bark beetle outbreaks and their dynamics during drought. For. Ecol. Manag. 2022, 523, 120480. [Google Scholar] [CrossRef]
- Schebeck, M.; Schopf, A.; Ragland, G.J.; Stauffer, C.; Biedermann, P.H.W. Evolutionary ecology of the bark beetles Ips typographus and Pityogenes chalcographus. Bull. Entomol. Res. 2023, 113, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Eidmann, H.H. Impact of bark beetles on forests and forestry in Sweden. J. Appl. Ent. 1992, 114, 193–200. [Google Scholar] [CrossRef]
- Marini, L.; Lindelöw, A.; Jönsson, A.M.; Wulff, S.; Schroeder, L.M. Population dynamics of the spruce bark beetle: A long-term study. Oikos 2013, 122, 1768–1776. [Google Scholar] [CrossRef]
- Marini, L.; Økland, B.; Jönsson, A.M.; Bentz, B.; Carroll, A.; Forster, B.; Gregoire, J.C.; Hurling, R.; Nageleisen, L.M.; Netherer, S. Climate drivers of bark beetle outbreak dynamics in Norway spruce forests. Ecography 2017, 40, 1426–1435. [Google Scholar] [CrossRef]
- Schroeder, M.; Cocos, D. Performance of the tree-killing bark beetles Ips typographus and Pityogenes chalcographus in non-indigenous lodgepole pine and their historical host Norway spruce. Agric. For. Entomol. 2017, 20, 347–357. [Google Scholar] [CrossRef]
- Seidl, R.; Schelhaas, M.J.; Lexer, M.J. Unraveling the drivers of intensifying forest disturbance regimes in Europe. Glob. Chang. Biol. 2011, 17, 2842–2852. [Google Scholar] [CrossRef]
- Markovic, C.; Stojanovic, A. Differences in bark beetle (Ips typographus and Pityogenes chalcographus) abundance in a strict spruce reserve and the surrounding spruce forests of Serbia. Phytoparasitica 2010, 38, 31–37. [Google Scholar] [CrossRef]
- Göthlin, E.; Schroeder, L.M.; Lindelöw, A. Attacks by Ips typographus and Pityogenes chalcographus on windthrown spruces (Picea abies) during the two years following a storm felling. Scand. J. For. Res. 2000, 15, 542–549. [Google Scholar] [CrossRef]
- Eriksson, M.; Lilja, S.; Roininen, H. Dead wood creation and restoration burning: Implications for bark beetles and beetle induced tree deaths. For. Ecol. Manag. 2006, 231, 205–213. [Google Scholar] [CrossRef]
- Jurc, M.; Perko, M.; Dzeroski, S.; Demsar, D.; Hrasovec, B. Spruce bark beetles (Ips typographus, Pityogenes chalcographus, Col.: Scolytidae) in the Dinaric mountain forests of Slovenia: Monitoring and modeling. Ecol. Model. 2006, 194, 219–226. [Google Scholar] [CrossRef]
- Berthelot, S.; Fruhbrodt, T.; Hajek, P.; Nock, C.A.; Dormann, C.F.; Bauhus, J.; Frund, J. Tree diversity reduces the risk of bark beetle infestation for preferred conifer species, but increases the risk for less preferred hosts. J. Ecol. 2021, 109, 2649–2661. [Google Scholar] [CrossRef]
- Yang, R.J.; Cao, R.Y.; Gong, X.; Feng, J.M. Large shifts of niche and range in the golden apple snail (Pomacea canaliculate), an aquatic invasive species. Ecosphere 2023, 14, e4391. [Google Scholar] [CrossRef]
- Brown, J.L. SDMtoolbox: A python-based GIS toolkit for landscape genetic, biogeographic and species distribution model analyses. Methods Ecol. Evol. 2014, 5, 694–700. [Google Scholar] [CrossRef]
- Brown, J.L.; Bennett, J.R.; French, C.M. SDMtoolbox 2.0: The next generation Python-based GIS toolkit for landscape genetic, biogeographic and species distribution model analyses. PeerJ 2017, 5, e4095. [Google Scholar] [CrossRef] [PubMed]
- Fick, S.E.; Hijmans, R.J. WorldClim2: New 1 km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Zhang, M.Z.; Xu, Z.F.; Han, Y.; Guo, W.D. Evaluation of CMIP6 models toward dynamical downscaling over 14 CORDEX domains. Clim. Dyn. 2022, 50, 803–817. [Google Scholar] [CrossRef]
- Duarte, A.; Wolcott, D.M.; Chow, T.E.; Ricca, M.A. Identifying potential habitat for the endangered Aleutian shield fern using topographical characteristics. J. Fish Wildl. Manag. 2012, 3, 303–310. [Google Scholar] [CrossRef]
- Nauyoma, L.T.; Wassenaar, T.D.; Mfune, J.K.E.; van der Waal, C.; Kwembeya, E.G. Field surveys and revegetation experiments show that simulation of topographical habitat features could improve the chances of successful restoration for the threatened succulent Juttadinteria albata. Restor. Ecol. 2019, 27, 730–737. [Google Scholar] [CrossRef]
- Beckerath, X.V.; Benadi, G.; Gilg, O.; Sittler, B.; Yannic, G.; Klein, A.M.; Eitzinger, B. Long-term monitoring reveals topographical features and vegetation that explain winter habitat use of an Arctic rodent. Arct. Sci. 2022, 8, 349–361. [Google Scholar] [CrossRef]
- Michalak, J.L.; Lawler, J.J.; Roberts, D.R.; Carroll, C. Distribution and protection of climatic refugia in North America. Conserv. Biol. 2018, 32, 1414–1425. [Google Scholar] [CrossRef]
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- Dormann, C.F.; Elith, J.; Bacher, S.; Buchmann, C.; Carl, G.; Carre, G.; Marquez, J.R.G.; Jaime, R.; Gruber, B.; Lafourcade, B.; et al. Collinearity: A review of methods to deal with it and a simulation study evaluating their performance. Ecography 2013, 36, 27–46. [Google Scholar] [CrossRef]
- Thuiller, W.; Lafourcade, B.; Engler, R.; Araujo, M.B. BIOMOD—A platform for ensemble forecasting of species distributions. Ecography 2009, 32, 369–373. [Google Scholar] [CrossRef]
- Barbet-Massin, M.; Jiguet, F.; Albert, C.H.; Thuiller, W. Selecting pseudo-absences for species distribution models: How, where and how many? Methods Ecol. Evol. 2012, 3, 327–338. [Google Scholar] [CrossRef]
- Liu, C.; Newell, G.; White, M. On the selection of thresholds for predicting species occurrence with presence–only data. Ecol. Evol. 2016, 6, 337–348. [Google Scholar] [CrossRef]
- Nie, P.X.; Feng, J.M. Global niche and range shifts of Batrachochytrium dendrobatidis, a highly virulent amphibian-killing fungus. Fungal. Biol. 2022, 126, 809–816. [Google Scholar] [CrossRef]
- Faccoli, M.; Bernardinelli, I. Composition and elevation of spruce forests affect susceptibility to bark beetle attacks: Implications for forest management. Forests 2014, 5, 88–102. [Google Scholar] [CrossRef]
- Nielsen, D.G.; Muilenburg, V.L.; Herms, D.A. Interspecific variation in resistance of Asian, European, and North American birches (Betula spp.) to bronze birch borer (Coleoptera: Buprestidae). Environ. Entomol. 2011, 40, 648–653. [Google Scholar] [CrossRef] [PubMed]
- Dilts, E.; Steele, M.O.; Engler, J.D.; Pelton, E.M.; Jepsen, S.J.; McKnight, S.J.; Taylor, A.R.; Fallon, C.E.; Black, S.H.; Cruz, E.E.; et al. Host plants and climate structure habitat associations of the western monarch butterfly. Front. Ecol. Evol. 2019, 7, 188. [Google Scholar] [CrossRef]
- Dang, Y.Q.; Zhang, Y.L.; Wang, X.Y.; Xin, B.; Quinn, N.F.; Duan, J.J. Retrospective analysis of factors affecting the distribution of an invasive wood-boring insect using native range data: The importance of host plants. J. Pest Sci. 2021, 94, 981–990. [Google Scholar] [CrossRef]
- Ding, F.Y.; Fu, J.Y.; Jiang, D.; Hao, M.M.; Lin, G. Mapping the spatial distribution of Aedes aegypti and Aedes albopictus. Acta Trop. 2018, 178, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Dickens, B.L.; Sun, H.; Jit, M.; Cook, A.R.; Carrasco, L.R. Determining environmental and anthropogenic factors which explain the global distribution of Aedes aegypti and Ae. albopictus. BMJ Glob. Health 2018, 3, e000801. [Google Scholar] [CrossRef]
- Root, R.B. Organization of a plant–arthropod association in simple and diverse habitats: Fauna of collards (Brassica oleracea). Ecol. Monogr. 1973, 43, 95–120. [Google Scholar] [CrossRef]
- Rocca, F.D.; Milanesi, P. The new dominator of the world: Modeling the global distribution of the Japanese beetle under land use and climate change scenarios. Land 2022, 11, 567. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Norway Spruce | The Eight-Spined Beetle | The Six-Spined Beetle | ||||||
|---|---|---|---|---|---|---|---|---|
| Category | Predictors | IVs | Category | Predictors | IVs | Category | Predictors | IVs |
| Climate | Bio4 | 0.5482 | Host availability | Host availability | 0.8194 | Host availability | Host availability | 0.7331 |
| Climate | Bio1 | 0.2407 | Climate | Bio10 | 0.0746 | Climate | Bio11 | 0.0856 |
| Climate | Bio5 | 0.1479 | Climate | Bio4 | 0.0683 | Land-use | Cropland | 0.0513 |
| Land-use | Primf | 0.0852 | Topography | Ele | 0.0531 | Land-use | Paster | 0.0377 |
| Land-use | Cropland | 0.0419 | Land-use | Primf | 0.0358 | Climate | Bio15 | 0.0319 |
| Climate | Bio18 | 0.0291 | Climate | Bio9 | 0.0299 | Land-use | Secdf | 0.0251 |
| Topography | Ele | 0.0208 | Climate | Bio15 | 0.0253 | Climate | Bio16 | 0.0205 |
| Climate | Bio2 | 0.0159 | Climate | Bio17 | 0.0245 | Land-use | Primn | 0.0195 |
| Land-use | Urban | 0.0154 | Land-use | Cropland | 0.0213 | Topography | Slope | 0.0168 |
| Climate | Bio14 | 0.0108 | Land-use | Urban | 0.0207 | Land-use | Urban | 0.0128 |
| Land-use | Secdf | 0.0089 | Climate | Bio2 | 0.0154 | Climate | Bio8 | 0.0121 |
| Climate | Bio8 | 0.0057 | Land-use | Paster | 0.0148 | Topography | Ele | 0.0094 |
| Land-use | Paster | 0.0045 | Land-use | Primn | 0.0148 | Land-use | Rang | 0.0086 |
| Land-use | Secdn | 0.0045 | Climate | Bio18 | 0.0144 | Climate | Bio2 | 0.0079 |
| Land-use | Rang | 0.0040 | Land-use | Secdn | 0.0130 | Land-use | Primf | 0.0071 |
| Land-use | Primn | 0.0023 | Land-use | Rang | 0.0071 | Topography | Aspect | 0.0035 |
| Climate | Bio15 | 0.0018 | Climate | Bio8 | 0.0067 | |||
| Topography | Slope | 0.0012 | Topography | Aspect | 0.0038 | |||
| Topography | Aspect | 0.0001 | ||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, R.; Feng, J. Future Range Shifts Suggest That the Six-Spined Spruce Bark Beetle Might Pose a Greater Threat to Norway Spruce in Europe than the Eight-Spined Spruce Bark Beetle. Forests 2023, 14, 2048. https://doi.org/10.3390/f14102048
Cao R, Feng J. Future Range Shifts Suggest That the Six-Spined Spruce Bark Beetle Might Pose a Greater Threat to Norway Spruce in Europe than the Eight-Spined Spruce Bark Beetle. Forests. 2023; 14(10):2048. https://doi.org/10.3390/f14102048
Chicago/Turabian StyleCao, Runyao, and Jianmeng Feng. 2023. "Future Range Shifts Suggest That the Six-Spined Spruce Bark Beetle Might Pose a Greater Threat to Norway Spruce in Europe than the Eight-Spined Spruce Bark Beetle" Forests 14, no. 10: 2048. https://doi.org/10.3390/f14102048