
Linking Vegetation Diversity and Soils on Highway Slopes: A Case Study of the Zhengzhou–Xinxiang Section of the Beijing–Hong Kong–Macau Highway

Abstract
:1. Introduction
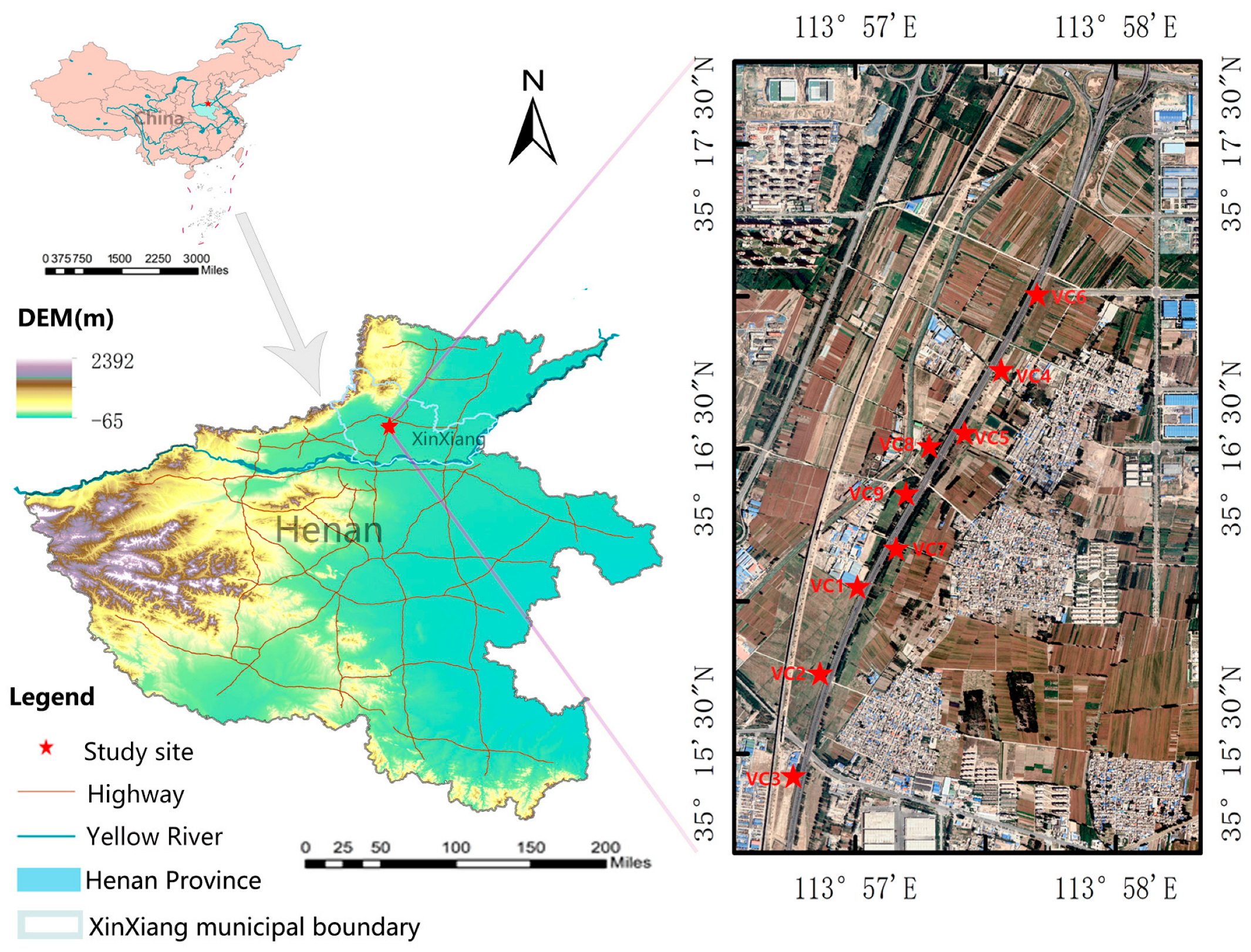
2. General Characteristics of the Study Area
3. Materials and Methods
3.1. Vegetation Community Research
3.2. Determination of Vegetation Community Diversity
3.3. Determination of Soil Physicochemical Properties
3.3.1. Soil Chemical Properties
3.3.2. Soil Physical Properties
3.4. Data Processing and Analysis
3.4.1. Redundancy Analysis
3.4.2. Coupling Analysis
4. Results
4.1. Species Composition and Vegetation Diversity of Different Vegetation Communities
4.2. Soil Physicochemical Characteristics of Different Vegetation Communities
4.2.1. Soil Chemical Characteristics
4.2.2. Soil Physical Characteristics
4.3. Analysis of the Relationship between Vegetation Community Diversity and Environmental Factors
4.4. Analysis of Vegetation Community Diversity Coupled with Environmental Factors
4.5. Analysis of the Coupling between Vegetation Community Diversity and Environmental Factors
5. Discussion
5.1. Relationship between Vegetation Community Diversity and Environmental Factors on Highway Slopes
5.2. Coupling Degree between the Diversity of Different Vegetation Communities and Environmental Factors
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Forman, R.T.T. Road Ecology: Science and Solutions; Island Press: Washington, DC, USA, 2003. [Google Scholar]
- Chen, L.L.; Mao, K.; Li, X.; Liu, L. Research Progress on Side Slopes Ecological Protection of Expressway. J. Anhui Agric. Sci. 2008, 36, 2023–2024, 2032. [Google Scholar]
- Li, S.F.; Li, Y.W.; Shi, J.L.; Zhao, T.N.; Yang, J.Y. Optimizing the formulation of external-soil spray seeding with sludge using the orthogonal test method for slope ecological protection. Ecol. Eng. 2017, 102, 527–535. [Google Scholar] [CrossRef]
- Zhao, J.H.; Gao, J.Z. Studies on Plant Invasion and Biodiversity Conservation of Ecological super highway—A case of Henan Province. In Proceedings of the International Conference on Civil, Architectural and Hydraulic Engineering (ICCAHE 2012), Zhangjiajie, China, 10–12 August 2012; pp. 1237–1241. [Google Scholar]
- Huang, W.; Liu, Z.; Zhou, C.Y.; Yang, X. Enhancement of soil ecological self-repair using a polymer composite material. Catena 2020, 188, 104443. [Google Scholar] [CrossRef]
- Berendse, F.; van Ruijven, J.; Jongejans, E.; Keesstra, S. Loss of Plant Species Diversity Reduces Soil Erosion Resistance. Ecosystems 2015, 18, 881–888. [Google Scholar] [CrossRef]
- Martins, M.D.; Angers, D.A. Different plant types for different soil ecosystem services. Geoderma 2015, 237, 266–269. [Google Scholar] [CrossRef]
- Wedin, D.A.; Tilman, D. Influence of Nitrogen Loading and Species Composition on the Carbon Balance of Grasslands. Science 1996, 274, 1720–1723. [Google Scholar] [CrossRef] [PubMed]
- Merila, P.; Malmivaara-Lamsa, M.; Spetz, P.; Stark, S.; Vierikko, K.; Derome, J.; Fritze, H. Soil organic matter quality as a link between microbial community structure and vegetation composition along a successional gradient in a boreal forest. Appl. Soil Ecol. 2010, 46, 259–267. [Google Scholar] [CrossRef]
- Paniagua, A.; Kammerbauer, J.; Avedillo, M.; Andrews, A.M. Relationship of soil characteristics to vegetation successions on a sequence of degraded and rehabilitated soils in Honduras. Agric. Ecosyst. Environ. 1999, 72, 215–225. [Google Scholar] [CrossRef]
- Wang, G.L.; Liu, G.B.; Xu, M.X. Above- and belowground dynamics of plant community succession following abandonment of farmland on the Loess Plateau, China. Plant Soil 2009, 316, 227–239. [Google Scholar] [CrossRef]
- Gao, R.; Ai, N.; Liu, G.; Liu, C.; Qiang, F. Characteristics of understory herb communities across time during restoration in coal mine reclamation areas and their coupling with soil properties. Acta Pratac. Sin. 2022, 31, 61–68. [Google Scholar] [CrossRef]
- Fukami, T.; Nakajima, M. Complex plant-soil interactions enhance plant species diversity by delaying community convergence. J. Ecol. 2013, 101, 316–324. [Google Scholar] [CrossRef]
- Cao, W.; Wang, Y.L.; Hu, Y.G.; Xu, E.K.; Yang, H.; Tian, G.H. Coupling degree between plantation vegetation community diversity and soil properties: Xinyang-Nanyang highway as an example. Sci. Soil Water Conserv. 2018, 16, 144–150. [Google Scholar] [CrossRef]
- Wang, T.P.; Yang, X.M. Analysis of Relationship between Species Diversity and Soil Properties of Plant Community on Freeway Slope. J. Fujian For. Sci. Technol. 2010, 37, 37–41+45. [Google Scholar]
- Pan, S.L.; Gu, B.; Li, J.X. Soil-property and plant diVersity of highway rocky slopes. Acta Ecol. Sin. 2012, 32, 8. [Google Scholar] [CrossRef]
- Sun, Y.; Shi, Y.; Tang, Y.; Tian, J.; Wu, X. Correlation between plant diversity and the physicochemical properties of soil microbes. Appl. Ecol. Environ. Res. 2019, 17, 10371–10388. [Google Scholar] [CrossRef]
- Li, S.; Su, P.; Zhang, H.; Zhou, Z.; Xie, T.; Shi, R.; Gou, W. Distribution patterns of desert plant diversity and relationship to soil properties in the Heihe River Basin, China. Ecosphere 2018, 9, e02355. [Google Scholar] [CrossRef]
- Yao, M.M.; Guo, C.W.; He, F.C.; Zhang, Q.; Ren, G.H. Soil Stoichiometric Characteristics and its Relationship with Plant Diversity in Saline-Alkali Grassland of Northern Shanxi. Acta Agrestia Sin. 2021, 29, 2800–2807. [Google Scholar] [CrossRef]
- Ma, M.J.; Baskin, C.C.; Yu, K.L.; Ma, Z.; Du, G.Z. Wetland drying indirectly influences plant community and seed bank diversity through soil pH. Ecol. Indic. 2017, 80, 186–195. [Google Scholar] [CrossRef]
- Liang, J.A.; Wang, X.A.; Yu, Z.D.; Dong, Z.M.; Wang, J.C. Effects of Vegetation Succession on Soil Fertility within Farming-Plantation Ecotone in Ziwuling Mountains of the Loess Plateau in China. Sci. Agric. Sin 2010, 9, 1481–1491. [Google Scholar] [CrossRef]
- Chen, L.X. Soil Experiment Practice Course; Northeast University Press: Harbin, China, 2005. [Google Scholar]
- Meng, Q.; Wang, S.; Fu, Z.; Deng, Y.; Chen, H. Soil types determine vegetation communities along a toposequence in a dolomite peak-cluster depression catchment. Plant Soil 2022, 475, 5–22. [Google Scholar] [CrossRef]
- Wang, M.; Dong, Z.; Luo, W.; Lu, J.; Li, J. Spatial variability of vegetation characteristics, soil properties and their relationships in and around China’s Badain Jaran Desert. Environ. Earth Sci. 2015, 74, 6847–6858. [Google Scholar] [CrossRef]
- Liu, Z.Y.; Wang, Y.C.; Luo, P.Z.; He, G.X. Coupling relationships between plant diversity and soil characteristics in rocky desertification areas of western Hunan. J. For. Environ. 2021, 41, 471–477. [Google Scholar] [CrossRef]
- Xu, O.; Wei, T.X.; Li, F.; Li, Y.Y. Modeling the degree of coupling and interaction between plant community diversity and soil properties on highway slope. J. Beijing For. Univ. 2016, 38, 91–100. [Google Scholar] [CrossRef]
- Wang, Y.J. Research on the Impact of Returning Farmland to Forest on the Coupling of Agricultural Land Resources and Industrial Systems in Loess Hilly Areas. Master’s Thesis, North West Agriculture and Forestry University, Shaanxi, China, 2010. [Google Scholar]
- Pang, C.Q.; Qin, J.T.; Li, H.X.; Liu, J.H. Effects of Rice Straw Incorporation and Permanent Fallow on Soil Nutrient of Paddy Field in Northeastern Jiangxi Province. Soils 2013, 45, 604–609. [Google Scholar] [CrossRef]
- Da, Z.J.; Ai, Y.W.; Song, T.; Guo, P.J.; Wang, Q.; Li, W. Spatial and Seasonal Variability of Soil Water in Road Slopes. Bull. Soil Water Conserv. 2011, 31, 72–75. [Google Scholar] [CrossRef]
- Li, Y.Y.; Shao, M.G.; Zhang, X.C. Spatial Distribution of Soil Moisture and Available Phosphorus Content on Eroded Sloping Land. J. Soil Water Conserv. 2001, 15, 41–44. [Google Scholar] [CrossRef]
- Heilman, G.E.; Strittholt, J.R.; Slosser, N.C.; Dellasala, D.A. Forest fragmentation of the conterminous United States: Assessing forest intactness through road density and spatial characteristics: Forest fragmentation can be measured and monitored in a powerful new way by combining remote sensing, geographic information systems, and analytical software. Bioscience 2002, 52, 411–422. [Google Scholar] [CrossRef]
- Kammerbauer, H.; Selinger, H.; Römmelt, R.; Jöns, A.Z.; Knoppik, D.; Hock, B. Toxic effects of exhaust emissions on spruce Picea abies and their reduction by the catalytic converter. Environ. Pollut. 1986, 42, 133–142. [Google Scholar] [CrossRef]
- Ball, J.E.; Jenks, R.; Aubourg, D. An assessment of the availability of pollutant constituents on road surfaces. Sci. Total Environ. 1998, 209, 243–254. [Google Scholar] [CrossRef]
- Angold, P.G. The impact of a road upon adjacent heathland vegetation: Effects on plant species composition. J. Appl. Ecol. 1997, 34, 409–417. [Google Scholar] [CrossRef]
- Spellerberg, I. Ecological effects of roads and traffic: A literature review. Glob. Ecol. Biogeogr. 1998, 7, 317–333. [Google Scholar] [CrossRef]
- Andersen, T.; Elser, J.J.; Hessen, D.O. Stoichiometry and population dynamics. Ecol. Lett. 2004, 7, 884–900. [Google Scholar] [CrossRef]
- Olde, H.V.; Wassen, M.J.; Verkroost, A.W.M.; Ruiter, P.C.D. Species richnessproductivity patterns differ between n, p, and klimited wetlands. Ecology 2003, 84, 2191–2199. [Google Scholar] [CrossRef]
- Wang, L.; Wang, P.; Sheng, M.; Tian, J. Ecological stoichiometry and environmental influencing factors of soil nutrients in the karst rocky desertification ecosystem, southwest China. Glob. Ecol. Conserv. 2018, 16, e00449. [Google Scholar] [CrossRef]
- Wu, G.L.; Zhang, Z.N.; Wang, D.; Shi, Z.H.; Zhu, Y.J. Interactions of soil water content heterogeneity and species diversity patterns in semi-arid steppes on the Loess Plateau of China. J. Hydrol. 2014, 519, 1362–1367. [Google Scholar] [CrossRef]
- Zhao, Z.C.; Xia, Z.X.; Xiong, S.Y.; Wu, B.; Xu, W.N. Evolution of Soil Properties in Vegetation Restoration Process on Disturbed Slopes. Bull. Soil Water Conserv. 2013, 33, 82–86. [Google Scholar] [CrossRef]
- Gamfeldt, L.; Snll, T.; Bagchi, R.; Jonsson, M.; Bengtsson, J. Higher levels of multiple ecosystem services are found in forests with more tree species. Nat. Commun. 2011, 4, 1340. [Google Scholar] [CrossRef]
- Hu, D.; Lv, G.H.; Wang, H.F.; Yang, Q.; Cai, Y. Response of desert plant diversity and stability to soil factors based on water gradient. Acta Ecol. Sin. 2021, 41, 6738–6748. [Google Scholar]
- Hao, J.F.; Zhou, R.H.; Yao, S.L.; Yu, J.; Cheng, C.G. Effects of the second generation wild boar grazing on species diversity and soil physicochemical properties of coniferous-broad-leaved mixed forest in Jiajin Mountain, China. Chin. J. Plant Ecol. 2022, 46, 197–207. [Google Scholar] [CrossRef]
- Li, X.R.; Zhang, J.G.; Liu, L.C.; Chen, H.S.; Shi, Q.H. Plant diversity in the process of succession of artificiai vegetation types and environment in an arid desertregion of China. Chin. J. Plant Ecol. 2000, 257–261. [Google Scholar] [CrossRef]
- Hagen-Thorn, A.; Callesen, I.; Armolaitis, K.; Nihlgård, B. The impact of six European tree species on the chemistry of mineral topsoil in forest plantations on former agricultural land. For. Ecol. Manag. 2004, 195, 373–384. [Google Scholar] [CrossRef]
- Derong, X.; Kun, T.; Liquan, Z. Relationship between plant diversity and soil fertility in Napahai wetland of Northwestern Yunnan Plateau. Acta Ecol. Sin. 2008, 28, 3116–3124. [Google Scholar]
- Lagomarsino, A.; Moscatelli, M.C.; Tizio, A.D.; Mancinelli, R.; Grego, S.; Marinari, S. Soil biochemical indicators as a tool to assess the short-term impact of agricultural management on changes in organic C in a Mediterranean environment. Ecol. Indic. 2009, 9, 518–527. [Google Scholar] [CrossRef]
- Yang, D.D.; Luo, C.D.; Guan, Y.B.; Liang, J. The Dynamic of Soil Nutrient under Forest and Grass Composite Pattern in Area of Conversion of Farmland to Forests. Sci. Silv. Sin. 2007, 43, 101–105. [Google Scholar]
- Li, T.T.; Tang, Y.B.; Zhou, R.H.; Yu, F.Y.; Dong, H.J.; Wang, M.; Hao, J.F. Understory plant diversity and its relationship with soil physicochemical properties in different plantations in Yunding Mountain. Acta Ecol. Sin. 2021, 41, 1168–1177. [Google Scholar] [CrossRef]
- Xu, L.; Feng, F.; Liu, Y.; Yang, Y.P.; Zheng, W.D. Relationship between plant species diversity and soil chemical properties in coal gangue dump: Early stage of ecological restoration in Lingwu Mining Area. Coal Sci. Technol. 2020, 48, 97–104. [Google Scholar] [CrossRef]
- Zhao, F.Y.; Guo, Y.J.; Cao, B.; Liang, L.Z. Grey Incidence Analysis of Vegetation Diversity Characteristics and Soil Factors on Highway Side Slopes in Yanqing County. Bull. Soil Water Conserv. 2010, 30, 64–68+90. [Google Scholar] [CrossRef]
- Lin, L.; Dai, L.; Lin, Z.B.; Wu, J.T.; Yan, W.; Wang, Z.J. Plant Diversity and Its Relationship with Soil Physicochemical Properties of Urban Forest Communities in Central Guizhou. Ecol. Environ. Sci. 2021, 30, 2130–2141. [Google Scholar] [CrossRef]
- Nan, G.W.; Han, L.; He, X.Y.; Rong, H.Q.; Ma, L. Dynamic changes of species diversity in herb layer of the Robinia pseudoacacia plantation. J. For. Environ. 2022, 42, 491–497. [Google Scholar] [CrossRef]
- Huo, H.; Feng, Q.; Su, Y.H. Shrub communities and environmental variables responsible for species distribution patterns in an alpine zone of the Qilian Mountains, northwest China. J. Mount. Sci. 2015, 12, 166–176. [Google Scholar] [CrossRef]
- Takahashi, K.; Murayama, Y. Effects of topographic and edaphic conditions on alpine plant species distribution along a slope gradient on Mount Norikura, central Japan. Ecol. Res. 2014, 29, 823–833. [Google Scholar] [CrossRef]
- Gao, G.G.; Hu, Y.K.; Li, K.H.; Xiao, H.L.; Gong, Y.M.; Yin, W. Relationships between Species Diversity of Plant Communities and Soil Factors in Alpine-cold Grassland. Bull. Soil Water Conserv. 2009, 29, 118–122. [Google Scholar] [CrossRef]
- Zhang, F.; Li, Y.C.; Wang, X.; Zhu, J.X. Effect of rangeland degradation on biomass allocation in alpine meadows on the Qinghai-Tibet Plateau, China. Pratac. Sci. 2021, 38, 1451–1458. [Google Scholar]
- Wen, P.Y.; Jin, G.Z. Effects of topography on species diversity in a typical mixed broadleaved-Korean pine forest. Acta Ecol. Sin. 2018, 39, 945–956. [Google Scholar] [CrossRef]
- Oztas, T.; Koc, A.; Comakli, B. Changes in vegetation and soil properties along a slope on overgrazed and eroded rangelands. J. Arid Environ. 2003, 55, 93–100. [Google Scholar] [CrossRef]
- Qin, S.; Fan, Y.; Liu, H.B.; Wang, Z.Y. Study on the Relations between Topographical Factors and the Spatial Distributions of Soil Nutrients. Res. Soil Water Conserv. 2008, 15, 46–49+52. [Google Scholar]
- Dorairaj, D.; Osman, N. Present practices and emerging opportunities in bioengineering for slope stabilization in Malaysia: An overview. PeerJ 2021, 9, e10477. [Google Scholar] [CrossRef]
- Odum, E.P. Productivity and Biodiversity: A Two-Way Relationship. Bull. Ecol. Soc. Am. 1998, 79, 125. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| VC | Name of Vegetation Community | Slope | Orientation |
|---|---|---|---|
| VC1 | Guandi B. papyrifera | 32° | West Slope |
| VC2 | Guandi U. pumila + A. fruticosa | 46° | West Slope |
| VC3 | Guandi A. fruticosa + Artemisia annua | 46° | West Slope |
| VC4 | Zhangdi B. papyrifera + A. fruticosa | 31° | East Slope |
| VC5 | Zhangdi U. pumila + A. fruticosa | 31° | East Slope |
| VC6 | Zhangdi A. fruticosa | 35° | East Slope |
| VC7 | Yuandi B. papyrifera | 30° | East Slope |
| VC8 | Yuandi Humulus scandens + B. papyrifera | 31° | West Slope |
| VC9 | Yuandi Setaria viridis (L.) Beauv. + A. fruticosa | 31° | West Slope |
| Coupling Degree (C) | 0 ≤ C ≤ 0.4 | 0.4 ≤ C ≤ 0.5 | 0.5 ≤ C ≤ 0.6 | 0.6 ≤ C ≤ 0.7 | 0.7 ≤ C ≤ 0.8 | 0.8 ≤ C ≤ 0.9 | 0.9 ≤ C ≤ 1.0 |
|---|---|---|---|---|---|---|---|
| Type of coordination | Serious incoordination | Middle incoordination | Light incoordination | Light coordination | Middle coordination | Favorable coordination | Superior coordination |
| VC | R | H | E | D |
|---|---|---|---|---|
| VC1 | 4.33 ± 1.53 cd | 1.14 ± 0.25 c | 0.80 ± 0.07 a | 0.62 ± 0.09 b |
| VC2 | 8.00 ± 2.65 a | 1.58 ± 0.33 abc | 0.77 ± 0.04 ab | 0.72 ± 0.07 ab |
| VC3 | 8.67 ± 2.31 a | 1.48 ± 0.34 abc | 0.71 ± 0.21 ab | 0.66 ± 0.19 ab |
| VC4 | 4.67 ± 0.58 bcd | 1.19 ± 0.15 bc | 0.77 ± 0.04 ab | 0.63 ± 0.05 b |
| VC5 | 4.67 ± 2.08 bcd | 1.22 ± 0.40 bc | 0.82 ± 0.09 a | 0.65 ± 0.12 ab |
| VC6 | 6.67 ± 0.58 abc | 1.62 ± 0.09 ab | 0.85 ± 0.05 a | 0.77 ± 0.01 ab |
| VC7 | 3.00 ± 1.00 d | 0.65 ± 0.21 d | 0.62 ± 0.09 b | 0.37 ± 0.13 c |
| VC8 | 7.67 ± 2.52 ab | 1.30 ± 0.37 bc | 0.64 ± 0.08 b | 0.64 ± 0.14 ab |
| VC9 | 9.67 ± 1.16 a | 1.88 ± 0.06 a | 0.83 ± 0.03 a | 0.82 ± 0.02 a |
| VC | AN/(mg/kg) | TN/(g/kg) | TP/((g/kg) | TK/((g/kg) | AP/(mg/kg) | AK/(mg/kg) | OM/(g/kg) | pH |
|---|---|---|---|---|---|---|---|---|
| VC1 | 43.22 ± 9.75 a | 0.95 ± 0.15 a | 0.62 ± 0.01 a | 17.01 ± 0.23 cde | 2.91 ± 0.58 ab | 216.5 ± 17.52 a | 23.2 ± 5.3 a | 8.58 ± 0.03 e |
| VC2 | 32.65 ± 12.25 abc | 0.74 ± 0.27 ab | 0.53 ± 0.05 bc | 17.22 ± 0.09 bcd | 1.86 ± 0.93 bc | 154 ± 13.7 bcd | 17.39 ± 6.52 ab | 8.75 ± 0.1 cd |
| VC3 | 24.49 ± 6.6 bcd | 0.56 ± 0.13 bc | 0.49 ± 0.03 c | 16.78 ± 0.23 e | 1.48 ± 0.74 c | 128.5 ± 21.63 cd | 12.82 ± 3.53 bc | 8.67 ± 0.07 cde |
| VC4 | 23.29 ± 3.25 cd | 0.52 ± 0.01 bc | 0.54 ± 0.04 bc | 17.45 ± 0.41 ab | 1.38 ± 0.18 c | 146.33 ± 7.08 cd | 11.27 ± 0.36 bc | 8.8 ± 0.09 bc |
| VC5 | 19.69 ± 7.93 cd | 0.46 ± 0.14 c | 0.5 ± 0.02 c | 17.59 ± 0.07 ab | 1.4 ± 0.05 c | 135.17 ± 15.28 cd | 9.94 ± 3.18 c | 8.62 ± 0.13 de |
| VC6 | 24.25 ± 1.5 bcd | 0.53 ± 0.08 bc | 0.58 ± 0.01 ab | 17.47 ± 0.25 ab | 2.8 ± 0.67 ab | 162.83 ± 17.28 bc | 11.61 ± 1.36 bc | 8.74 ± 0.03 cd |
| VC7 | 19.21 ± 2.73 d | 0.47 ± 0.08 c | 0.49 ± 0.02 c | 17.6 ± 0.15 a | 1.57 ± 0.61 c | 117.33 ± 27.89 d | 10.11 ± 1.74 bc | 8.95 ± 0.09 a |
| VC8 | 23.05 ± 6.28 cd | 0.56 ± 0.17 bc | 0.54 ± 0.05 bc | 17.37 ± 0.1 abc | 1.93 ± 0.71 bc | 152.83 ± 44.74 bcd | 12.83 ± 4.28 bc | 8.92 ± 0.08 ab |
| VC9 | 36.98 ± 10.55 ab | 0.83 ± 0.22 a | 0.6 ± 0.02 a | 16.97 ± 0.2 de | 3.07 ± 0.84 a | 184.67 ± 9.57 ab | 20.5 ± 7.21 a | 8.64 ± 0.03 de |
| VC | WM (%) | SBD (g/cm−3) | SWC (%) | CP (%) | PG (%) | FS (%) | CS (%) |
|---|---|---|---|---|---|---|---|
| VC1 | 9.17 ± 1.96 bcd | 1.24 ± 0.06 b | 1.21 ± 0.08 c | 8.94 ± 0.54 a | 44.61 ± 1.24 a | 45.14 ± 1.75 b | 1.32 ± 0.16 ab |
| VC2 | 5.51 ± 0.38 e | 1.35 ± 0.09 a | 1.1 ± 0.08 d | 6.37 ± 1.65 b | 35.33 ± 10.21 ab | 55.65 ± 9.7 a | 2.65 ± 2.16 ab |
| VC3 | 4.56 ± 0.05 e | 1.38 ± 0.03 a | 1.05 ± 0.02 d | 6.63 ± 0.6 1 b | 35.03 ± 2.58 ab | 55.37 ± 3.12 ab | 2.97 ± 0.34 ab |
| VC4 | 9.63 ± 0.39 bcd | 1.4 ± 0.03 a | 1.03 ± 0.03 d | 8.11 ± 1.2 a b | 42.13 ± 5.45 ab | 48.57 ± 4.89 ab | 1.19 ± 1.39 ab |
| VC5 | 6.59 ± 0.64 de | 1.42 ± 0.04 a | 1.02 ± 0.04 d | 6.37 ± 0.85 b | 33.94 ± 6.14 b | 56.77 ± 6.55 a | 2.92 ± 0.48 ab |
| VC6 | 15.76 ± 5.3 a | 1.39 ± 0.06 a | 1.06 ± 0.06 d | 7.94 ± 1.33 ab | 39.91 ± 5.26 ab | 51.54 ± 5.4 ab | 0.61 ± 1.01 b |
| VC7 | 10.38 ± 2.11 bc | 0.99 ± 0.01 d | 1.51 ± 0.01 a | 6.56 ± 2.26 b | 34.03 ± 10.7 ab | 56 ± 10.19 a | 3.41 ± 2.75 a |
| VC8 | 7.4 ± 1.16 cde | 1.11 ± 0.06 c | 1.36 ± 0.08 b | 8.34 ± 0.23 ab | 40.09 ± 2.21 ab | 50.66 ± 1.04 ab | 0.91 ± 1.37 b |
| VC9 | 12.18 ± 1.28 ab | 1.21 ± 0.05 b | 1.23 ± 0.07 c | 7.11 ± 0.76 ab | 40.86 ± 3.95 ab | 50.93 ± 3.54 ab | 1.1 ± 1.26 ab |
| Correlation | R | H | E | D | MV |
|---|---|---|---|---|---|
| AN | 0.739 | 0.728 | 0.705 | 0.724 | 0.724 |
| TN | 0.726 | 0.712 | 0.694 | 0.711 | 0.711 |
| TP | 0.685 | 0.744 | 0.741 | 0.757 | 0.732 |
| TK | 0.646 | 0.663 | 0.671 | 0.682 | 0.665 |
| AP | 0.693 | 0.713 | 0.726 | 0.728 | 0.715 |
| AK | 0.711 | 0.732 | 0.748 | 0.749 | 0.735 |
| OM | 0.726 | 0.703 | 0.695 | 0.715 | 0.710 |
| pH | 0.645 | 0.652 | 0.630 | 0.680 | 0.652 |
| WM | 0.659 | 0.683 | 0.731 | 0.729 | 0.701 |
| SBD | 0.661 | 0.743 | 0.773 | 0.773 | 0.738 |
| SWC | 0.690 | 0.665 | 0.658 | 0.681 | 0.674 |
| SG | 0.752 | 0.714 | 0.695 | 0.714 | 0.719 |
| CP | 0.647 | 0.664 | 0.675 | 0.684 | 0.667 |
| PG | 0.657 | 0.683 | 0.702 | 0.710 | 0.688 |
| SF | 0.694 | 0.718 | 0.702 | 0.720 | 0.709 |
| CS | 0.672 | 0.701 | 0.698 | 0.715 | 0.697 |
| MV | 0.688 | 0.701 | 0.703 | 0.717 | 0.702 |
| VC | Coupling Degree (C) | Type of Coordination |
|---|---|---|
| VC1 | 0.676 | Light coordination |
| VC2 | 0.725 | Middle coordination |
| VC3 | 0.638 | Light coordination |
| VC4 | 0.794 | Middle coordination |
| VC5 | 0.725 | Middle coordination |
| VC6 | 0.741 | Middle coordination |
| VC7 | 0.623 | Light coordination |
| VC8 | 0.699 | Light coordination |
| VC9 | 0.698 | Light coordination |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, W.; Zhu, N.; Meng, Z.; Lv, C.; Chen, Y.; Wang, G. Linking Vegetation Diversity and Soils on Highway Slopes: A Case Study of the Zhengzhou–Xinxiang Section of the Beijing–Hong Kong–Macau Highway. Forests 2023, 14, 1863. https://doi.org/10.3390/f14091863
Cao W, Zhu N, Meng Z, Lv C, Chen Y, Wang G. Linking Vegetation Diversity and Soils on Highway Slopes: A Case Study of the Zhengzhou–Xinxiang Section of the Beijing–Hong Kong–Macau Highway. Forests. 2023; 14(9):1863. https://doi.org/10.3390/f14091863
Chicago/Turabian StyleCao, Wei, Niuniu Zhu, Zhenyu Meng, Chenxi Lv, Yue Chen, and Guojie Wang. 2023. "Linking Vegetation Diversity and Soils on Highway Slopes: A Case Study of the Zhengzhou–Xinxiang Section of the Beijing–Hong Kong–Macau Highway" Forests 14, no. 9: 1863. https://doi.org/10.3390/f14091863