Forecasting Appropriate Habitats for Rare and Endangered Indocalamus Species in China in Response to Climate Change

 , , ,
, , ,
Abstract
:1. Introduction
2. Research Methods
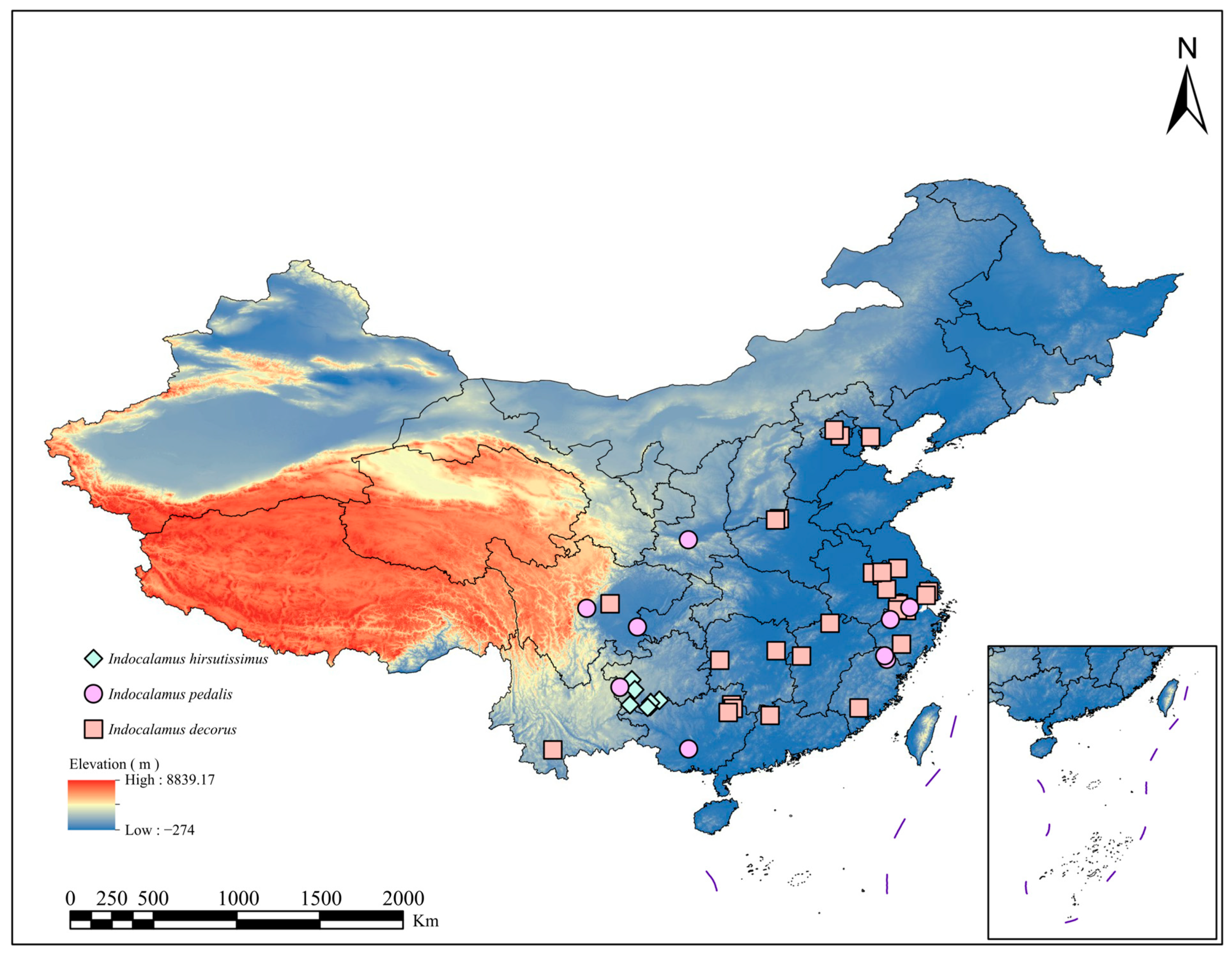
2.1. Gathering and Filtering of Sample Data
2.2. Screening of Bioclimatic Variables
2.3. Model Development, Optimization, and Evaluation
2.4. Delineation of Suitable Areas and Analysis of Low-Impact Regions
2.5. Analysis of Spatial Pattern Changes and Core Distribution Shifts
3. Results
3.1. Evaluation of Model Accuracy and Contributions of Environmental Variables
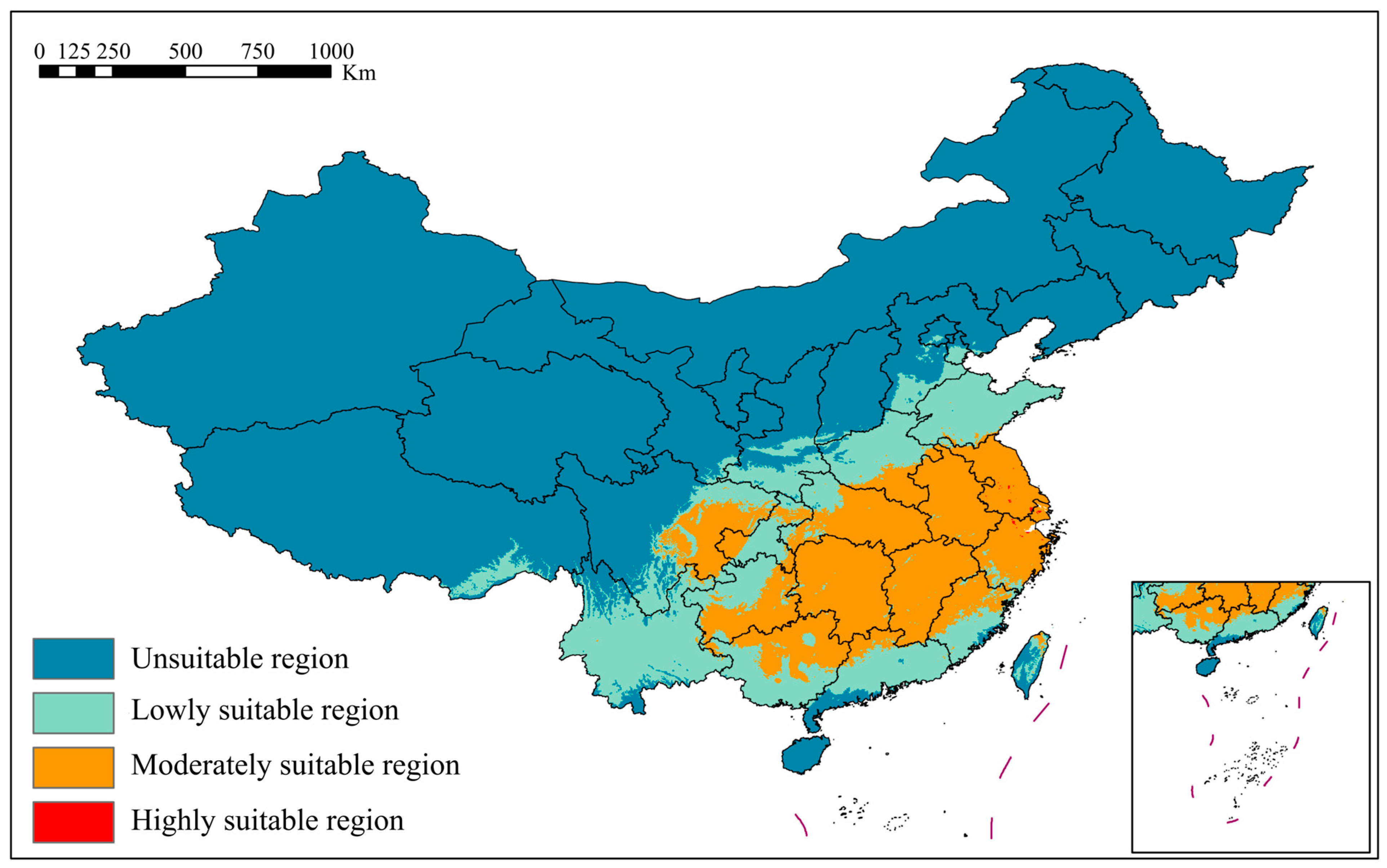
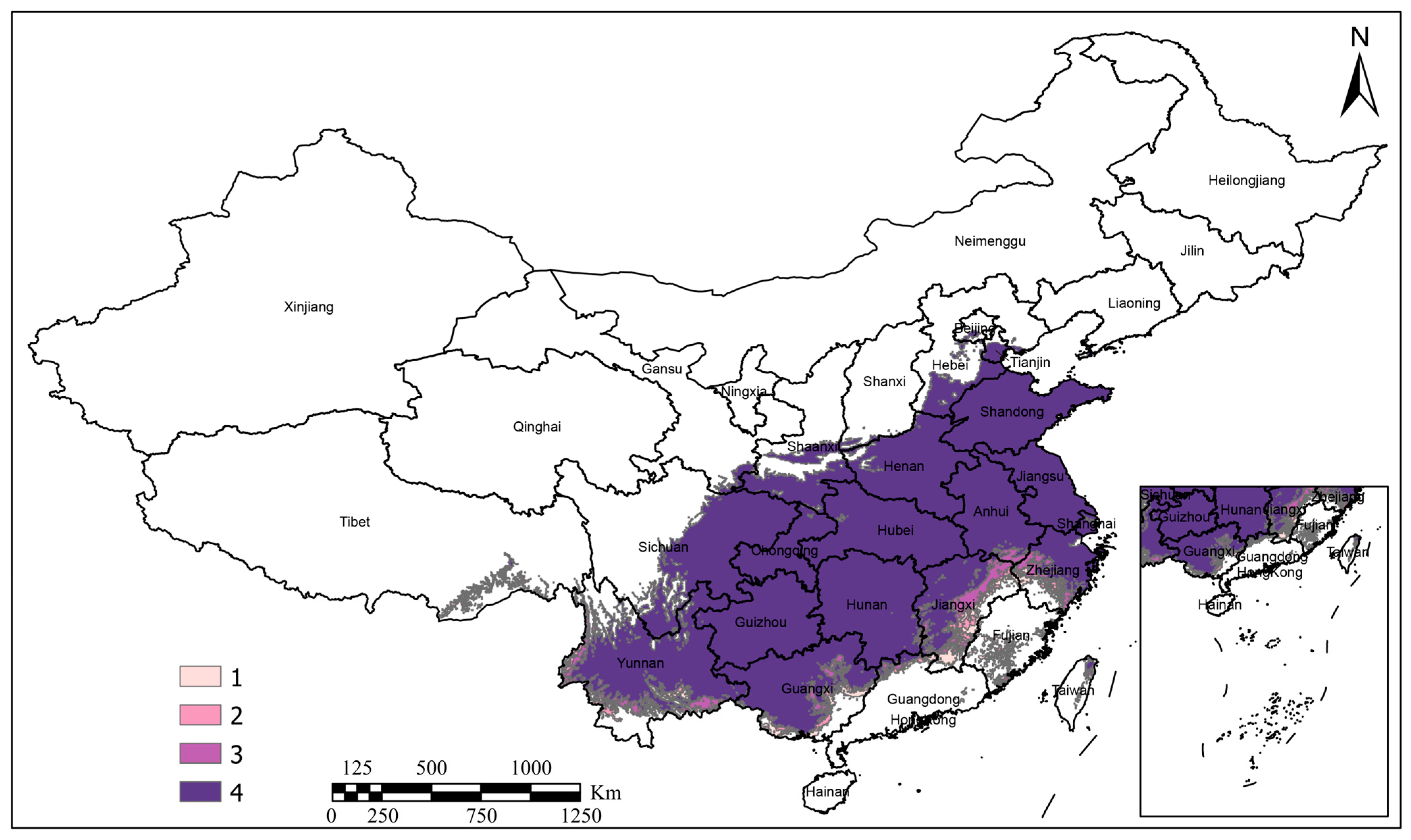
3.2. Analysis of Current Potential Suitable Distribution Areas
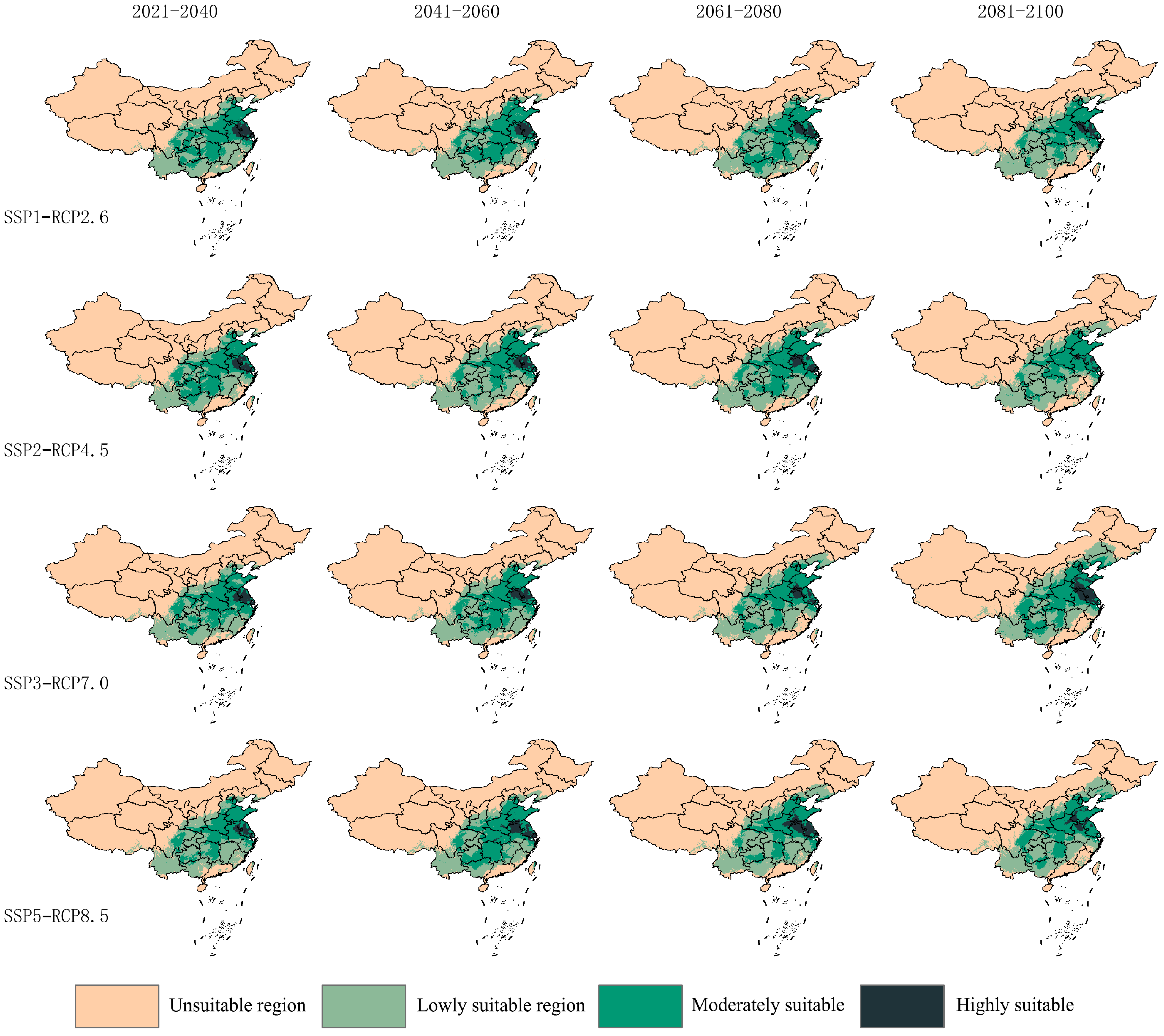
3.3. Future Potential Suitable Areas
3.4. Low-Impact Suitable Distribution Areas
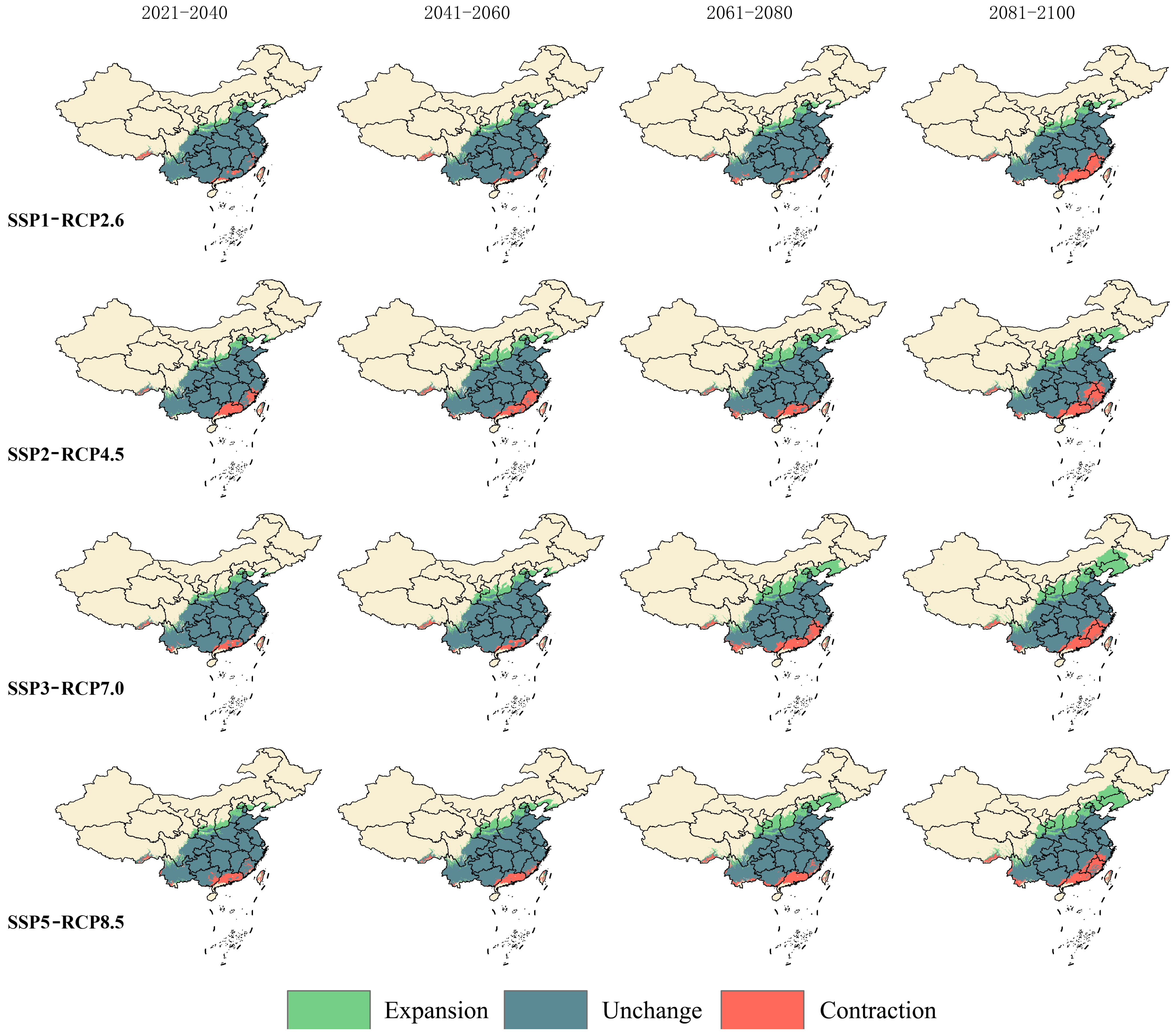
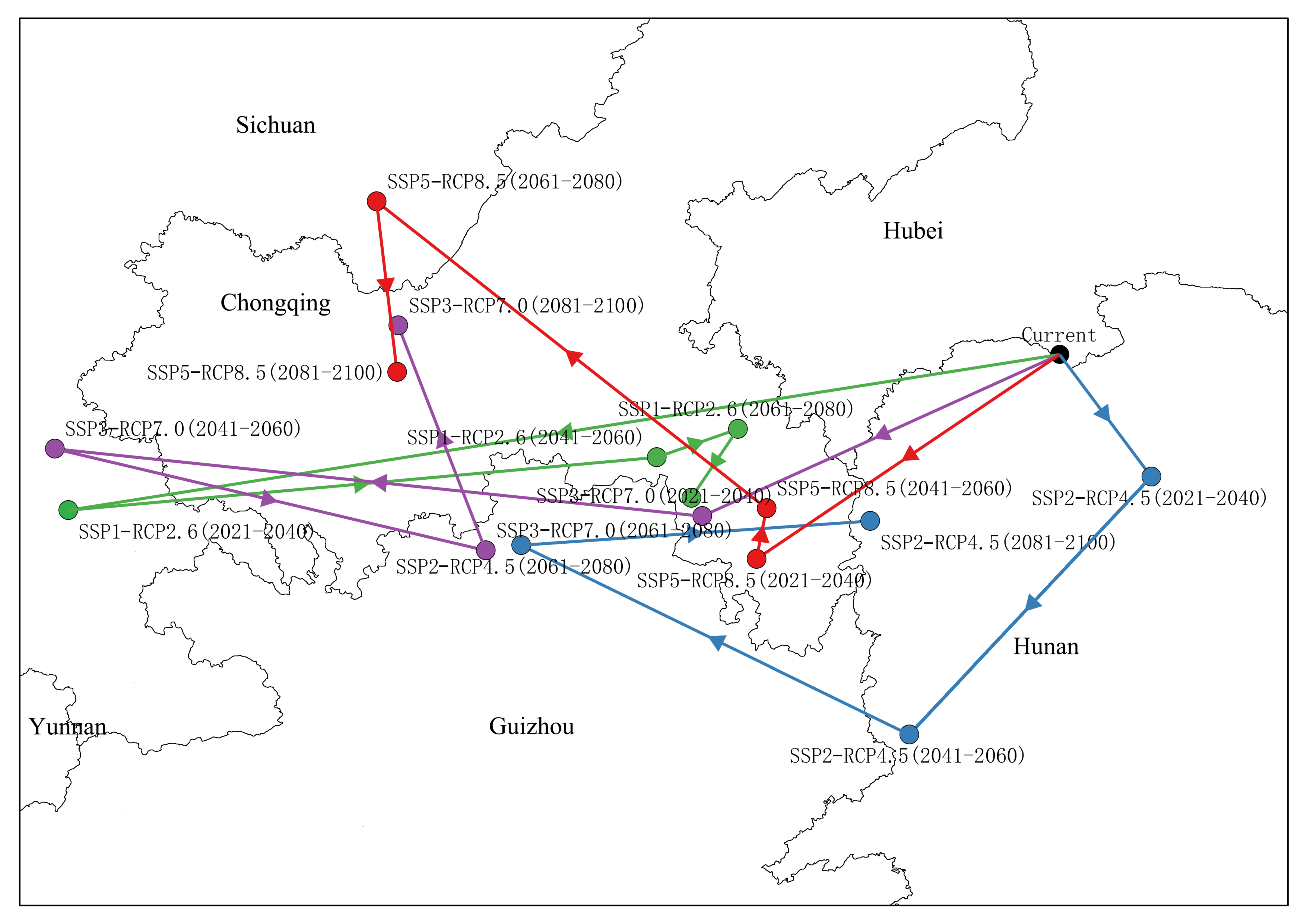
3.5. Trends in the Migration of Suitable Area Distribution Centers
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Record, S.; Fitzpatrick, M.C.; Finley, A.O.; Veloz, S.; Ellison, A.M. Should Species Distribution Models Account for Spatial Autocorrelation? A Test of Model Projections across Eight Millennia of Climate Change. Glob. Ecol. Biogeogr. 2013, 22, 760–771. [Google Scholar] [CrossRef]
- Santos-Hernández, A.F.; Monterroso-Rivas, A.I.; Granados-Sánchez, D.; Villanueva-Morales, A.; Santacruz-Carrillo, M. Projections for Mexico’s Tropical Rainforests Considering Ecological Niche and Climate Change. Forests 2021, 12, 119. [Google Scholar] [CrossRef]
- Carroll, C.; Dunk, J.R.; Moilanen, A. Optimizing Resiliency of Reserve Networks to Climate Change: Multispecies Conservation Planning in the Pacific Northwest, USA. Glob. Chang. Biol. 2010, 16, 891–904. [Google Scholar] [CrossRef]
- Braunisch, V.; Coppes, J.; Arlettaz, R.; Suchant, R.; Schmid, H.; Bollmann, K. Selecting from Correlated Climate Variables: A Major Source of Uncertainty for Predicting Species Distributions under Climate Change. Ecography 2013, 36, 971–983. [Google Scholar] [CrossRef]
- Cursach, J.; Moragues, E.; Rita, J. The Key Role of Accompanying Species in the Response of the Critically Endangered Naufraga balearica (Apiaceae) to Climatic Factors. Plant Ecol. 2018, 219, 561–576. [Google Scholar] [CrossRef]
- Menzel, A.; Sparks, T.H.; Estrella, N.; Roy, D.B. Altered Geographic and Temporal Variability in Phenology in Response to Climate Change. Glob. Ecol. Biogeogr. 2006, 15, 498–504. [Google Scholar] [CrossRef]
- Costion, C.M.; Simpson, L.; Pert, P.L.; Carlsen, M.M.; John Kress, W.; Crayn, D. Will Tropical Mountaintop Plant Species Survive Climate Change? Identifying Key Knowledge Gaps Using Species Distribution Modelling in Australia. Biol. Conserv. 2015, 191, 322–330. [Google Scholar] [CrossRef]
- Forester, B.R.; DeChaine, E.G.; Bunn, A.G. Integrating Ensemble Species Distribution Modelling and Statistical Phylogeography to Inform Projections of Climate Change Impacts on Species Distributions. Divers. Distrib. 2013, 19, 1480–1495. [Google Scholar] [CrossRef]
- Yang, Z.; Bai, Y.; Alatalo, J.M.; Huang, Z.; Yang, F.; Pu, X.; Wang, R.; Yang, W.; Guo, X. Spatio-Temporal Variation in Potential Habitats for Rare and Endangered Plants and Habitat Conservation Based on the Maximum Entropy Model. Sci. Total Environ. 2021, 784, 147080. [Google Scholar] [CrossRef]
- Ye, X.; Zhao, G.; Zhang, M.; Cui, X.; Fan, H.; Liu, B. Distribution Pattern of Endangered Plant Semiliquidambar cathayensis (Hamamelidaceae) in Response to Climate Change after the Last Interglacial Period. Forests 2020, 11, 434. [Google Scholar] [CrossRef]
- Li, X.; Mao, F.; Du, H.; Zhou, G.; Xing, L.; Liu, T.; Han, N.; Liu, Y.; Zhu, D.; Zheng, J.; et al. Spatiotemporal Evolution and Impacts of Climate Change on Bamboo Distribution in China. J. Environ. Manag. 2019, 248, 109265. [Google Scholar] [CrossRef] [PubMed]
- Cupertino-Eisenlohr, M.A.; Vinicius-Silva, R.; Meireles, L.D.; Eisenlohr, P.V.; Meira-Neto, J.A.A.; Santos-Goncalves, A.P. Stability or Breakdown under Climate Change? A Key Group of Woody Bamboos Will Find Suitable Areas in Its Richness Center. Biodivers. Conserv. 2017, 26, 1845–1861. [Google Scholar] [CrossRef]
- Bhandawat, A.; Sharma, V.; Sharma, H.; Sood, A.; Sharma, R.K. Development and Crosstransferability of Functionally Relevant Microsatellite Markers in Dendrocalamus latiflorus and Related Bamboo Species. J. Genet. 2015, 94, 48–55. [Google Scholar] [CrossRef]
- Wu, Z.Y.; Raven, P.; Hong, D.Y. Flora of China; Botanical Garden Press: St. Louis, MO, USA, 2013. [Google Scholar]
- Department of Wildlife Conservation and Nature Reserve Management, State Forestry Administration, Institute of Botany, Chinese Academy of Sciences. Illustrated Handbook of Rare and Endangered Plants of China; China Forestry Publishing House: Beijing, China, 2013. [Google Scholar]
- Zhang, M.; Keenan, T.F.; Luo, X.; Serra-Diaz, J.M.; Li, W.; King, T.; Cheng, Q.; Li, Z.; Andriamiarisoa, R.L.; Raherivelo, T.N.A.N.; et al. Elevated CO2 Moderates the Impact of Climate Change on Future Bamboo Distribution in Madagascar. Sci. Total Environ. 2022, 810, 152235. [Google Scholar] [CrossRef]
- Zhao, H.; Yang, X.; Shi, S.; Xu, Y.; Yu, X.; Ye, X. Climate-Driven Distribution Changes for Bashania fargesii in the Qinling Mountains and Its Implication for Panda Conservation. Glob. Ecol. Conserv. 2023, 46, e02610. [Google Scholar] [CrossRef]
- Wu, Y.; Li, J.; Yu, L.; Wang, S.; Lv, Z.; Long, H.; Zhai, J.; Lin, S.; Meng, Y.; Cao, Z.; et al. Overwintering Performance of Bamboo Leaves, and Establishment of Mathematical Model for the Distribution and Introduction Prediction of Bamboos. Front. Plant Sci. 2023, 14, 1255033. [Google Scholar] [CrossRef]
- Vicente, J.R.; Fernandes, R.F.; Randin, C.F.; Broennimann, O.; Goncalves, J.; Marcos, B.; Pocas, I.; Alves, P.; Guisan, A.; Honrado, J.P. Will Climate Change Drive Alien Invasive Plants into Areas of High Protection Value? An Improved Model-Based Regional Assessment to Prioritise the Management of Invasions. J. Environ. Manag. 2013, 131, 185–195. [Google Scholar] [CrossRef]
- Tabor, J.A.; Koch, J.B. Ensemble Models Predict Invasive Bee Habitat Suitability Will Expand under Future Climate Scenarios in Hawai’i. Insects 2021, 12, 443. [Google Scholar] [CrossRef]
- Oduor, A.M.O.; Yang, B.; Li, J. Alien Ornamental Plant Species Cultivated in Taizhou, Southeastern China, May Experience Greater Range Expansions than Native Species under Future Climates. Glob. Ecol. Conserv. 2023, 41, e02371. [Google Scholar] [CrossRef]
- Anderson, R.P. A Framework for Using Niche Models to Estimate Impacts of Climate Change on Species Distributions. Ann. N. Y. Acad. Sci. 2013, 1297, 8–28. [Google Scholar] [CrossRef]
- Xu, Z.; Luo, Y.; Qin, S.; Zhu, G.; Li, D. Current Status and Geographic Distribution of Bamboo Specimens in Chinese Herbaria. Biodiversity 2021, 29, 897–909. [Google Scholar]
- Elith, J.; Leathwick, J.R. Species Distribution Models: Ecological Explanation and Prediction across Space and Time. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 677–697. [Google Scholar] [CrossRef]
- Araújo, M.B.; Guisan, A. Five (or so) Challenges for Species Distribution Modelling. J. Biogeogr. 2006, 33, 1677–1688. [Google Scholar] [CrossRef]
- Marmion, M.; Luoto, M.; Heikkinen, R.K.; Thuiller, W. The Performance of State-of-the-Art Modelling Techniques Depends on Geographical Distribution of Species. Ecol. Model. 2009, 220, 3512–3520. [Google Scholar] [CrossRef]
- Zhao, Z.; Guo, Y.; Wei, H.; Ran, Q.; Gu, W. Predictions of the Potential Geographical Distribution and Quality of a Gynostemma pentaphyllum Base on the Fuzzy Matter Element Model in China. Sustainability 2017, 9, 1114. [Google Scholar] [CrossRef]
- Brown, J.L. SDMtoolbox: A Python-Based GIS Toolkit for Landscape Genetic, Biogeographic and Species Distribution Model Analyses. Methods Ecol. Evol. 2014, 5, 694–700. [Google Scholar] [CrossRef]
- Brown, J.L.; Bennett, J.R.; French, C.M. SDMtoolbox 2.0: The next Generation Python-Based GIS Toolkit for Landscape Genetic, Biogeographic and Species Distribution Model Analyses. PeerJ 2017, 5, e4095. [Google Scholar] [CrossRef]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J. A Statistical Explanation of MaxEnt for Ecologists. Divers. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Pearson, R.G.; Raxworthy, C.J.; Nakamura, M.; Townsend Peterson, A. Predicting Species Distributions from Small Numbers of Occurrence Records: A Test Case Using Cryptic Geckos in Madagascar. J. Biogeogr. 2007, 34, 102–117. [Google Scholar] [CrossRef]
- Dou, P.; Dong, Y.; Chen, L.; Yang, H.-Q. Modeling the Potential Distribution of Different Types of Dendrocalamus sinicus, the Strongest Woody Bamboo in the World, with MaxEnt Model. Peerj 2022, 10, e13847. [Google Scholar] [CrossRef]
- Meena, R.K.; Bhandari, M.S.; Thakur, P.K.; Negi, N.; Pandey, S.; Kant, R.; Sharma, R.; Sahu, N.; Avtar, R. MaxEnt-Based Potential Distribution Mapping and Range Shift under Future Climatic Scenarios for an Alpine Bamboo Thamnocalamus spathiflorus in Northwestern Himalayas. Land 2024, 13, 931. [Google Scholar] [CrossRef]
- Chen, L.-J.; Xie, Y.-Q.; He, T.-Y.; Chen, L.-Y.; Rong, J.-D.; Chen, L.-G.; Zheng, Y.-S. Projecting the Impacts of Climate Change, Soil, and Landscape on the Geographic Distribution of Ma Bamboo (Dendrocalamus latiflorus Munro) in China. Forests 2024, 15, 1321. [Google Scholar] [CrossRef]
- Tang, C.Q.; Matsui, T.; Ohashi, H.; Dong, Y.-F.; Momohara, A.; Herrando-Moraira, S.; Qian, S.; Yang, Y.; Ohsawa, M.; Luu, H.T.; et al. Identifying Long-Term Stable Refugia for Relict Plant Species in East Asia. Nat. Commun. 2018, 9, 4488. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Mi, Z.-Y.; Lu, C.; Zhang, X.-F.; Chen, L.-J.; Wang, S.-Q.; Niu, J.-F.; Wang, Z.-Z. Predicting Potential Distribution of Ziziphus spinosa (Bunge) H.H. Hu Ex F.H. Chen in China under Climate Change Scenarios. Ecol. Evol. 2022, 12, e8629. [Google Scholar] [CrossRef]
- Xing, D.; Hao, Z. The Principle of Maximum Entropy and Its Application in Ecological Research. Biodiversity 2011, 19, 295–302. [Google Scholar]
- Keliang, Z.; Sun, L.; Tao, J. Impact of Climate Change on the Distribution of Euscaphis japonica (Staphyleaceae) Trees. Forests 2020, 11, 525. [Google Scholar] [CrossRef]
- Warren, D.L.; Matzke, N.J.; Cardillo, M.; Baumgartner, J.B.; Beaumont, L.J.; Turelli, M.; Glor, R.E.; Huron, N.A.; Simões, M.; Iglesias, T.L.; et al. ENMTools 1.0: An R Package for Comparative Ecological Biogeography. Ecography 2021, 44, 504–511. [Google Scholar] [CrossRef]
- Phillips, S.J.; Dudík, M. Modeling of Species Distributions with Maxent: New Extensions and a Comprehensive Evaluation. Ecography 2008, 31, 161–175. [Google Scholar] [CrossRef]
- Cahill, A.E.; Aiello-Lammens, M.E.; Fisher-Reid, M.C.; Hua, X.; Karanewsky, C.J.; Yeong Ryu, H.; Sbeglia, G.C.; Spagnolo, F.; Waldron, J.B.; Warsi, O.; et al. How Does Climate Change Cause Extinction? Proc. R. Soc. B Biol. Sci. 2013, 280, 20121890. [Google Scholar] [CrossRef]
- Warren, B.H.; Bakker, F.T.; Bellstedt, D.U.; Bytebier, B.; Claßen-Bockhoff, R.; Dreyer, L.L.; Edwards, D.; Forest, F.; Galley, C.; Hardy, C.R.; et al. Consistent Phenological Shifts in the Making of a Biodiversity Hotspot: The Cape Flora. BMC Evol. Biol. 2011, 11, 39. [Google Scholar] [CrossRef]
- Lobo, J.M.; Jiménez-Valverde, A.; Real, R. AUC: A Misleading Measure of the Performance of Predictive Distribution Models. Glob. Ecol. Biogeogr. 2008, 17, 145–151. [Google Scholar] [CrossRef]
- Warren, D.L.; Seifert, S.N. Ecological Niche Modeling in Maxent: The Importance of Model Complexity and the Performance of Model Selection Criteria. Ecol. Appl. 2011, 21, 335–342. [Google Scholar] [CrossRef] [PubMed]
- Arenas-Castro, S.; Gonçalves, J.F.; Moreno, M.; Villar, R. Projected Climate Changes Are Expected to Decrease the Suitability and Production of Olive Varieties in Southern Spain. Sci. Total Environ. 2020, 709, 136161. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Hu, J.; Hu, H.; Zhou, Y. Predictive Habitat Suitability Modeling of Deep-Sea Framework-Forming Scleractinian Corals in the Gulf of Mexico. Sci. Total Environ. 2020, 742, 140562. [Google Scholar] [CrossRef]
- Li, W.; Yu, Y.; Liu, P.; Tang, R.; Dai, Y.; Li, L.; Zhang, L. Identifying Climate Refugia and Its Potential Impact on Small Population of Asian Elephant (Elephas maximus) in China. Glob. Ecol. Conserv. 2019, 19, e00664. [Google Scholar] [CrossRef]
- Pan, J.; Fan, X.; Luo, S.; Zhang, Y.; Yao, S.; Guo, Q.; Qian, Z. Predicting the Potential Distribution of Two Varieties of Litsea coreana (Leopard-Skin Camphor) in China under Climate Change. Forests 2020, 11, 1159. [Google Scholar] [CrossRef]
- Zhao, Q.; Zhang, Y.; Li, W.-N.; Hu, B.-W.; Zou, J.-B.; Wang, S.-Q.; Niu, J.-F.; Wang, Z.-Z. Predicting the Potential Distribution of Perennial Plant Coptis chinensis Franch. in China under Multiple Climate Change Scenarios. Forests 2021, 12, 1464. [Google Scholar] [CrossRef]
- Smith, A.B.; Godsoe, W.; Rodríguez-Sánchez, F.; Wang, H.-H.; Warren, D. Niche Estimation Above and Below the Species Level. Trends Ecol. Evol. 2019, 34, 260–273. [Google Scholar] [CrossRef]
- Zurell, D.; Franklin, J.; König, C.; Bouchet, P.J.; Dormann, C.F.; Elith, J.; Fandos, G.; Feng, X.; Guillera-Arroita, G.; Guisan, A.; et al. A Standard Protocol for Reporting Species Distribution Models. Ecography 2020, 43, 1261–1277. [Google Scholar] [CrossRef]
- Yan, H.; Feng, L.; Zhao, Y.; Feng, L.; Wu, D.; Zhu, C. Prediction of the Spatial Distribution of Alternanthera philoxeroides in China Based on ArcGIS and MaxEnt. Glob. Ecol. Conserv. 2020, 21, e00856. [Google Scholar] [CrossRef]
- Yan, H.; Feng, L.; Zhao, Y.; Feng, L.; Zhu, C.; Qu, Y.; Wang, H. Predicting the Potential Distribution of an Invasive Species, Erigeron canadensis L., in China with a Maximum Entropy Model. Glob. Ecol. Conserv. 2020, 21, e00822. [Google Scholar] [CrossRef]
- Sun, S.; Zhang, Y.; Huang, D.; Wang, H.; Cao, Q.; Fan, P.; Yang, N.; Zheng, P.; Wang, R. The Effect of Climate Change on the Richness Distribution Pattern of Oaks (Quercus L.) in China. Sci. Total Environ. 2020, 744, 140786. [Google Scholar] [CrossRef]
- Feng, L.; Sun, J.; Shi, Y.; Wang, G.; Wang, T. Predicting Suitable Habitats of Camptotheca acuminata Considering Both Climatic and Soil Variables. Forests 2020, 11, 891. [Google Scholar] [CrossRef]
- López-Carrasco, C.; López-Sánchez, A.; San Miguel, A.; Roig, S. The Effect of Tree Cover on the Biomass and Diversity of the Herbaceous Layer in a Mediterranean Dehesa. Grass Forage Sci. 2015, 70, 639–650. [Google Scholar] [CrossRef]
- Ayalew, B.; Hylander, K.; Adugna, G.; Zewdie, B.; Zignol, F.; Tack, A.J.M. Impact of Climate and Management on Coffee Berry Disease and Yield in Coffee’s Native Range. Basic Appl. Ecol. 2024, 76, 25–34. [Google Scholar] [CrossRef]
- Büntgen, U.; Palosse, A.; Dolezal, J.; Liebhold, A. Introducing the Concepts of Range-Pinning and Allee Effects to Explain Reduced Temperature Sensitivity of Global Treeline Dynamics. Glob. Chang. Biol. 2024, 30, e17288. [Google Scholar] [CrossRef]
- Zhou, A.; Ge, B.; Chen, S.; Kang, D.; Wu, J.; Zheng, Y.; Ma, H. Leaf Ecological Stoichiometry and Anatomical Structural Adaptation Mechanisms of Quercus Sect. Heterobalanus in Southeastern Qinghai-Tibet Plateau. BMC Plant Biol. 2024, 24, 325. [Google Scholar] [CrossRef]
- Chmura, D.J.; Banach, J.; Kempf, M.; Kowalczyk, J.; Mohytych, V.; Szeligowski, H.; Buraczyk, W.; Kowalkowski, W. Growth and Productivity of European Beech Populations Show Plastic Response to Climatic Transfer at the North-Eastern Border of the Species Range. For. Ecol. Manag. 2024, 565, 122043. [Google Scholar] [CrossRef]
- Henley, B.J.; McGregor, H.V.; King, A.D.; Hoegh-Guldberg, O.; Arzey, A.K.; Karoly, D.J.; Lough, J.M.; DeCarlo, T.M.; Linsley, B.K. Highest Ocean Heat in Four Centuries Places Great Barrier Reef in Danger. Nature 2024, 632, 320–326. [Google Scholar] [CrossRef]
- Masson-Delmotte, V. The Physical Science Basis of Climate Change Empowering Transformations, Insights from the IPCC AR6 for a Climate Research Agenda Grounded in Ethics. PLoS Clim. 2024, 3, e0000451. [Google Scholar] [CrossRef]
- Ghosh, S.; Matthews, B.; Petchey, O.L. Temperature and Biodiversity Influence Community Stability Differently in Birds and Fishes. Nat. Ecol. Evol. 2024. [Google Scholar] [CrossRef] [PubMed]
- Hein, S.R.; Farleigh, K.; Berg, D.J. Riverscape Genomics of the Endangered Freshwater Mussel Lampsilis rafinesqueana. Freshw. Biol. 2024, 69, 1438–1453. [Google Scholar] [CrossRef]
- Hannah, L.; Midgley, G.F.; Millar, D. Climate Change-Integrated Conservation Strategies. Glob. Ecol. Biogeogr. 2002, 11, 485–495. [Google Scholar] [CrossRef]
- Forgiarini, C.; Meimberg, H.; Curto, M.; Stiehl-Alves, E.M.; Vijayan, T.; Engl, P.T.; Bräuchler, C.; Kollmann, J.; de Souza-Chies, T.T. Low Genetic Differentiation despite High Habitat Fragmentation in an Endemic and Endangered Species of Iridaceae from South America: Implications for Conservation. Bot. J. Linn. Soc. 2024, boae036. [Google Scholar] [CrossRef]
- Tomiolo, S.; Ward, D. Species Migrations and Range Shifts: A Synthesis of Causes and Consequences. Perspect. Plant Ecol. Evol. Syst. 2018, 33, 62–77. [Google Scholar] [CrossRef]
- Huang, E.; Chen, Y.; Fang, M.; Zheng, Y.; Yu, S. Environmental Drivers of Plant Distributions at Global and Regional Scales. Glob. Ecol. Biogeogr. 2021, 30, 697–709. [Google Scholar] [CrossRef]
- Wilting, A.; Cord, A.; Hearn, A.J.; Hesse, D.; Mohamed, A.; Traeholdt, C.; Cheyne, S.M.; Sunarto, S.; Jayasilan, M.-A.; Ross, J.; et al. Modelling the Species Distribution of Flat-Headed Cats (Prionailurus planiceps), an Endangered South-East Asian Small Felid. PLoS ONE 2010, 5, e9612. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| LIA Statistics | Shared Socio-Economic Pathways (SSPs) | |||
|---|---|---|---|---|
| entry 1 | SSP1-RCP 2.6 | SSP2-RCP4.5 | SSP3-RCP7.0 | SSP5-RCP 8.5 |
| Geographic area (×104 km2) | 237.61 | 229.96 | 229.69 | 231.89 |
| Percentage of current suitable area (%) | 94.36 | 91.33 | 91.22 | 92.05 |
| Percentage of SSP1-2.6 area (%) | 100.00 | 96.78 | 96.66 | 97.59 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xie, Y.; Huang, H.; Chen, L.; Xiao, J.; Weng, F.; Liu, J.; He, T.; Chen, L.; Rong, J.; Chen, L.; et al. Forecasting Appropriate Habitats for Rare and Endangered Indocalamus Species in China in Response to Climate Change. Forests 2024, 15, 1693. https://doi.org/10.3390/f15101693
Xie Y, Huang H, Chen L, Xiao J, Weng F, Liu J, He T, Chen L, Rong J, Chen L, et al. Forecasting Appropriate Habitats for Rare and Endangered Indocalamus Species in China in Response to Climate Change. Forests. 2024; 15(10):1693. https://doi.org/10.3390/f15101693
Chicago/Turabian StyleXie, Yanqiu, Hui Huang, Lijia Chen, Jihong Xiao, Feifan Weng, Jiaying Liu, Tianyou He, Lingyan Chen, Jundong Rong, Liguang Chen, and et al. 2024. "Forecasting Appropriate Habitats for Rare and Endangered Indocalamus Species in China in Response to Climate Change" Forests 15, no. 10: 1693. https://doi.org/10.3390/f15101693