1. Introduction
In eastern Canadian black spruce (
Picea mariana) boreal forests, terricolous lichens belonging to the genus
Cladonia tend to be progressively replaced by feather mosses as time since the last stand replacing fire increases [
1,
2,
3]. This gradual replacement is associated with the formation and closure of the dominant canopy, which leads to decreased light levels reaching the ground and increased humidity in the understory, conditions that are more favorable for mosses [
4,
5,
6].
However, even if terricolous lichen cover tends to decrease in older stands, these stands are also likely to contain species that are specifically described as late-successional [
7,
8,
9]. The later appearance of these species could be due to limitations in terms of dispersal or colonization capabilities [
10,
11,
12,
13] or to their association with micro-habitats that are only present in older forest, such as shaded coarse woody debris [
7,
9,
14,
15].
In western Quebec, post-fire succession is generally characterized by the progressive replacement of young, relatively dense even-aged black spruce stands with more open stands with irregular structure, mostly dominated by the same tree species after a period of
ca. 100–150 years [
16]. The presence of a flat topography, poorly drained clay soils, and a relatively cold climate also favor an accumulation of organic matter with time (process of paludification [
6,
17]). As a result of this process, old forests (>200 years) are often paludified and relatively unproductive compared with younger stands because the organic layer is a poor substrate for tree growth compared with mineral soil [
17]. These relatively unproductive uneven-aged forests are known to host a particularly rich liverwort flora [
18] and a high abundance of epiphytic lichens [
19], but their suitability for terricolous lichens is still poorly investigated.
Many studies have been undertaken on terricolous lichen communities in Canadian boreal forests (e.g. [
1,
20,
21]). However, few examined the relative contribution of the time elapsed since the last fire (time for species colonization and growth) and microhabitat quality and/or availability in shaping lichen communities. This question is important for determining the proper forest management strategies to maintain species diversity and abundance of terricolous lichen communities. For instance, if some lichen species are restricted to old forests primarily because they need a long period of time to establish and grow in the location, it will be necessary to emphasize a landscape management approach where forest patches containing source populations are maintained at all times in the forest mosaic. Alternatively, if terricolous lichen communities are mainly restricted to old forests because of microhabitat requirements, then a stand-scale approach where forest practices mimic or maintain these specific microhabitats could be successful. Improving our understanding of factors controlling lichen abundance in this ecosystem may also contribute to the preservation of the forest-dwelling ecotype of woodland caribou (
Rangifer tarundus caribou), an endangered subspecies that relies mainly on terricolous lichens, particularly during winter [
22,
23].
In this study, we document the structure and composition of terricolous lichen communities along a post-fire chronosequence (43 to 355 years old) in black spruce forests of western Quebec. We address two questions: (1) do species composition, richness, and abundance vary according to time since the last fire? and (2) is it time for colonization and growth or specific microhabitats that are the most important in explaining lichen composition, abundance, and richness?
4. Discussion
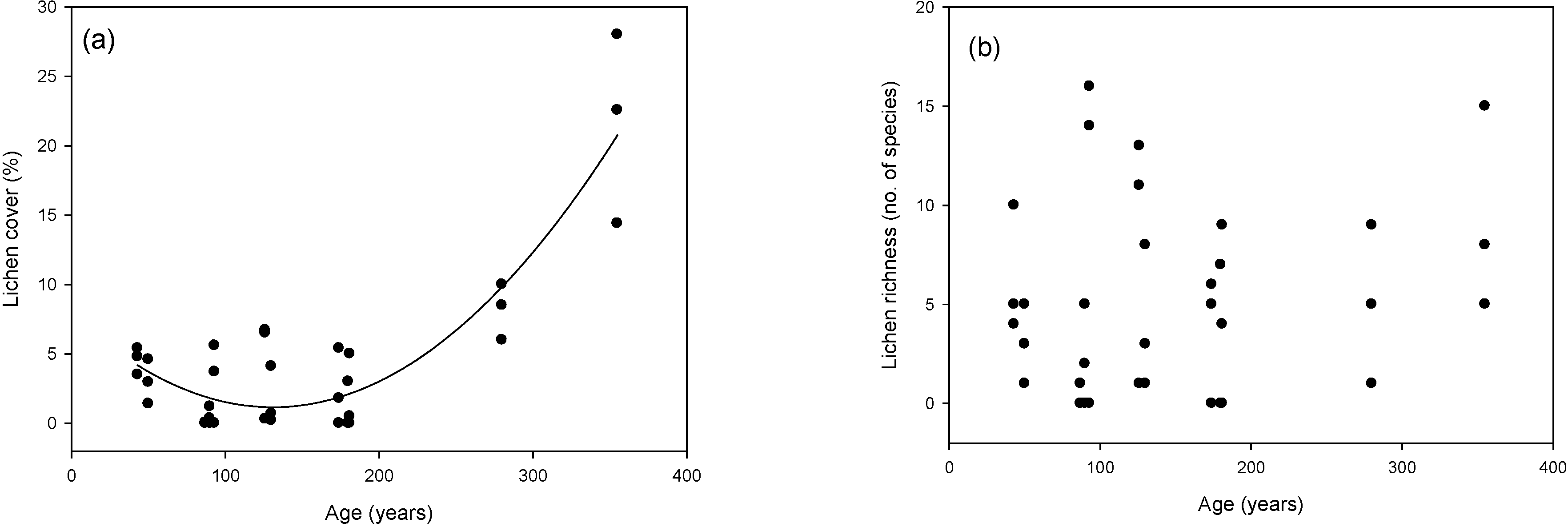
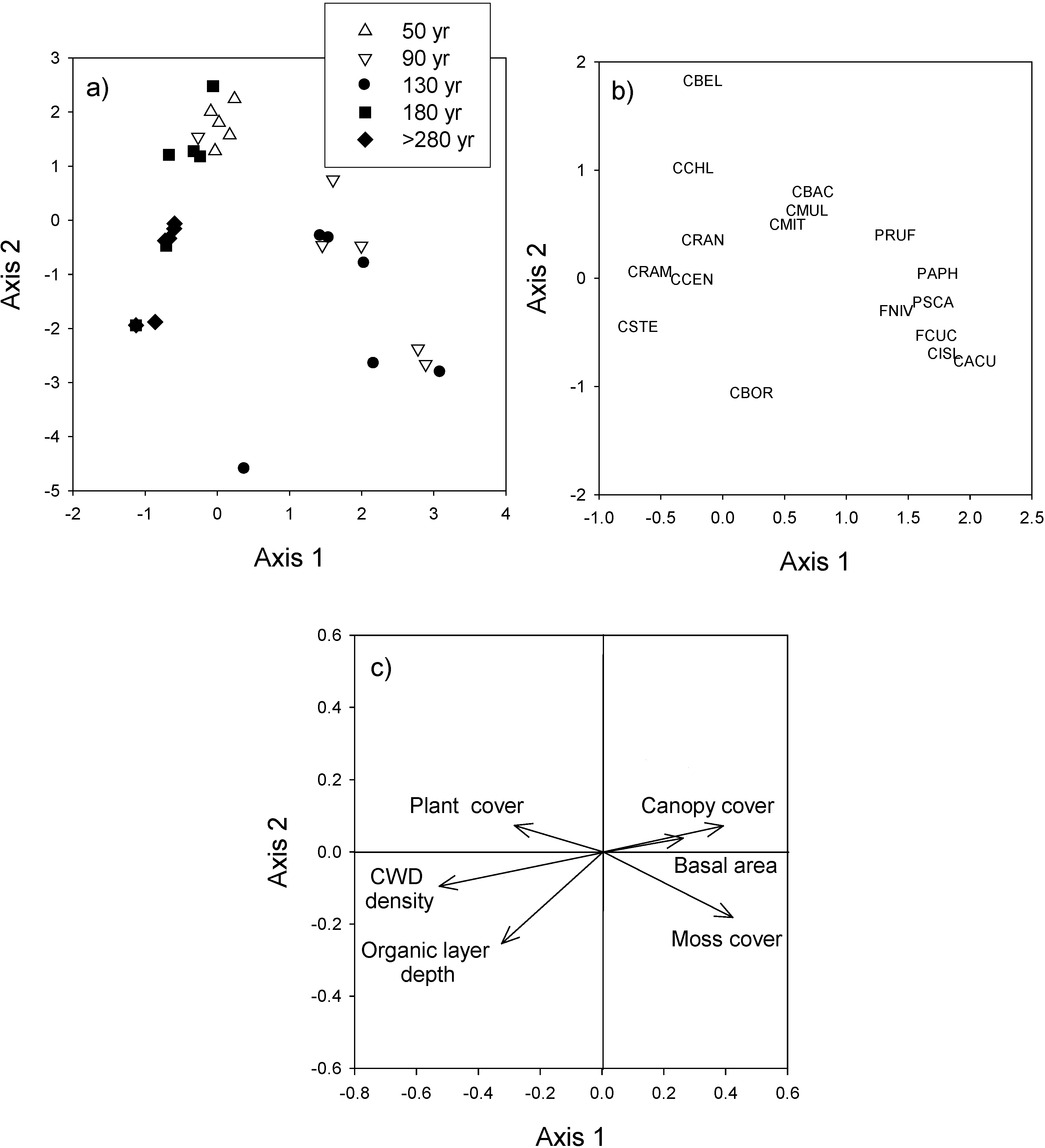
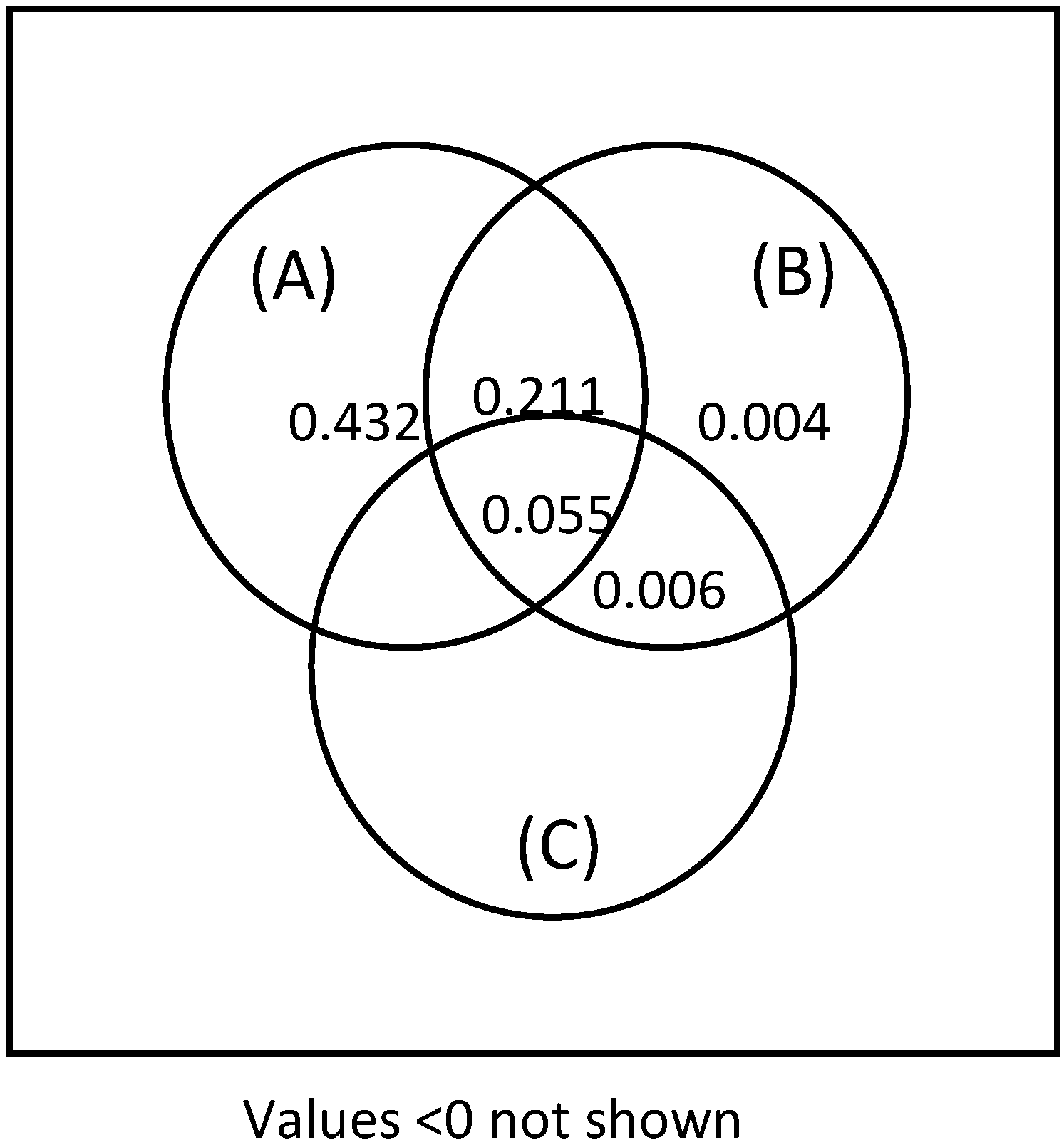
This study addresses a fundamental question for forest ecosystem management: is it stand age or stand characteristics that have the most influence on lichen species composition and abundance? Our results indicate that the time elapsed since the last fire was the factor that contributed the most to explaining terricolous lichen abundance and species composition. To our knowledge, this question has not been addressed before for terricolous lichens. Differences in composition arise principally from large foliose species (e.g. Flavocetraria cuculata, C. islandica, and F. nivellis) that typified forests from the 90- and 130-year age classes, and from high cover of caribou lichens associated with the youngest (Cladonia arbuscula and C. mitis) and oldest (C. rangiferina and C. stellaris)age classes.
Even if the limiting effect of lichen dispersal cannot be evaluated directly in this study because distance from the closest fire escapes were not evaluated, it is likely that the mode of dispersal of different species groups explains some part of the variations in species composition between age classes. For most
Cladonia species, dispersal strategy is either by soredia, small vegetative propagules that contain the mycobiont and the photobiont, or by spores [
38]. These modes of dispersal are considered efficient modes for lichen species dispersal [
39,
40,
41]. The main dispersal strategy for species belonging to the
Cladina group (
Cladonia mitis,
C. rangiferina,
C. stellaris) has been suggested to be through thallus fragments [
42,
43,
44,
45], which have a higher rate of successful establishment but a lower dispersal capability than other diaspores [
42,
46]. Heinken [
47] studied the dispersal of thallus fragments of seven terricolous lichen species and found the distance of dispersal was shorter than one meter by wind and about 10 m by animal dispersal. It is likely that the time elapsed since the last fire had a positive effect on
Cladina colonization by increasing the probability that large thallus fragments successfully reach the sites, particularly in the case of
C. stellaris and
C. rangiferina, which are particularly abundant in older (>280 years) forests.
In a general manner, lichen development rates have been considered to be relatively slow, and many years can elapse before the appearance of a mature thallus [
48] and the production of propagules [
49,
50]. Environmental factors such as snow accumulation on the diaspores can also have an effect through the reduction of the growing season for terricolous lichens [
48]. Overall, it can be reasonably expected that time since the last disturbance will have a positive effect on the probability of lichen establishment and growth in a site. The association between
C. stellaris and late successional forests [
51,
52] has previously been explained by the low growth rate of this species compared with the other caribou lichen species studied [
53]. However, as den Herder
et al. [
54] found in Finland, higher growth rates for
C stellaris compared with
C. mitis and
C. rangiferina were previously found in our study area, including in dense forest stands where the abundance of
C. stellaris is naturally relatively low [
55]. Thus, it is unlikely that the association between
C. stellaris and older forests in this study can be attributed to a lower growth rate for this species. An important effect of time since last disturbance, distinct from microhabitat availability, has also been suggested for other organisms potentially limited by dispersal in boreal forests such as liverworts [
18,
56,
57] and epiphytic lichens [
10,
19]. However, the exact biological mechanism underlying these statistical relationships remains poorly documented at the moment due to a poor documentation of traits associated with dispersal for these organisms [
58].
Despite the overall importance of stand age, the presence of suitable micro-habitats was an important prerequisite for some species. Thickness of the organic layer was important for overall lichen cover, species composition, and species richness. The establishment and growth of terricolous lichens may be favored by the accumulation of the organic layer and the increase in moss cover [
23] because these substrates increase the length of hydration periods [
59,
60]. The increase in organic matter depth may favor the abundance of
Cladonia stellaris over the other mat-forming species. Indeed, Kershaw and Rouse [
61] found that
C. stellaris metabolizes at a higher water saturation level than
C. rangiferina, and this latter is thus associated with dryer microsites than
C. stellaris [
62].
In this study, stand structural variables played a minor role for lichen abundance and species richness. Structural variables have been considered to influence lichen growth and were considered important in explaining the composition of terricolous lichen communities in many studies [
5,
43,
46,
51]. For example, Sulyma and Coxson [
63] have proposed that a decrease in ventilation in forest stands where canopy leaf area index is high (dense stands) can have the effect of increasing air humidity and favoring the establishment of feather mosses at the expense of lichens. Despite the weak relationships that we observed, it is possible that terricolous lichens were favored by the more diverse light conditions, lower humidity levels, and greater ventilation present in stands older than 180 years.
Finally, it is important to emphasize that this study took place on clay soils, which is the most representative site type in the region [
29]. Terricolous lichens are known to exhibit different community structures on different types of surficial deposits [
64], and thus the relationships with forest continuity could be different on soil types with coarser or finer textures, something that should be investigated by future studies.


{kind=link}
{kind=link}
{kind=link}


