1. Introduction
The forest ecosystems on Earth provide about 82% of the continental biomass and more than 50% of terrestrial biodiversity [
1,
2], the economic and commercial values of which are affected directly by wood quality. The abundant variation in the traits of trees strengthens their ability to adapt to different environmental conditions and provides the potential for the selection of trees with desirable traits [
3,
4,
5]. An important reason to breed trees is to increase the quantity and quality of wood. Wood properties are closely related to the complex biological processes of wood formation under cambial activity [
6,
7,
8], and most traits are complex, quantitative, and associated and regulated by multiple genes [
9,
10]. A better understanding of the variation in these traits of trees can enhance the use of germplasm collection and elucidate the genetic basis of phenotypic variation [
11]. In addition, to maximize the genetic gain in important traits, variation in and correlations between growth and wood quality must be assessed.
Chinese fir (
Cunninghamia lanceolata (Lamb.) Hook) is an economically valuable conifer with high yield, good wood quality, and multiple uses. It is a principal indigenous tree species that occupies approximately 25% of plantations in subtropical areas of southern China. Similar to other trees in breeding programs, provenance tests of Chinese fir have been performed since 1957, and a set of activities has been carried out mainly to improve its growth and adaptability. Presently, the Chinese fir breeding program has reached the third generation, and fine germplasm resources have been obtained [
12]. In 2004, a set of 700 Chinese fir plus trees from six provinces in southern China was conserved in the ex situ gene bank of Longshan State Forest, Guangdong Province, China (25°11′ N, 113°28′ E, 285–296 m above sea level). To reveal phenotypic variation and identify individuals with superior wood quality in this genetic resource, 10 phenotypic traits were investigated in 2014. Our results will improve the application of this germplasm resource and provide a starting point for further association analyses in Chinese fir.
2. Materials and Methods
In 2004, 700 Chinese fir plus individuals were collected from six provinces in China: 105, 113, 33, 49, 71, and 329 from the provinces of Guangxi, Jiangxi, Hunan, Guizhou, Fujian, and Guangdong, respectively. The scions of these trees were synchronously grafted onto two-year-old rootstocks in the ex situ gene bank of Longshan State Forest Farm, Guangdong Province, China (25°11′ N, 113°28′ E, 285–296 m above sea level), which is located in a subtropical region with a moderate climate throughout the year and receiving ample rainfall. Each clone had at least four ramets and random distribution with a plant spacing of 3 m × 3 m.
In 2014, 10 growth and wood traits were measured in all 700 clones, with at least three randomly selected ramets per clone. The growth traits included tree height (H), diameter at breast height (DBH), bark thickness (T), and stem volume (V). H, DBH, and T were measured during field surveys using methods described by Duan et al. [
13]. V was calculated according to the formula V = 0.000 058 777 042 ×
D1.9699831 ×
H0.89646157. The wood traits were the percentage of heartwood (P), wood basic density (WBD), hygroscopicity (Hy), tracheid length (L), tracheid diameter (D), and ratio of L to D (L/D). A 5.02 mm core was drilled from each tree at breast height using a tree growth cone and then placed in a plastic tube that was not completely sealed, to prevent wet rot. P was measured using the formula P =
r2/
R2 × 100%, where
r and
R represent the length of the heartwood and the sapwood, respectively [
13]. WBD and Hy were measured using the formulae WBD = 1/(W1/W2 − 0.346) and Hy = (W1 − W2)/W2, where W1 and W2 represent the water-saturated weight and the oven-dry weight, respectively [
14]. L and D were measured using the methods described by Huang et al. [
15]. Each sample was dissociated in a Franklin solution consisting of glacial acetic acid and 30% hydrogen peroxide at a 1:1 ratio for 16 h at 100 °C. Then, the remaining tracheid material was washed with deionized water until it was neutralized. Next, a tracheid smear was made to measure the lengths and widths of tracheids under a color closed-circuit television (CCTV) video camera (Panasonic SDII, Osaka, Japan). Three smears were made for each sample, and no fewer than 30 values were obtained in total.
Microsoft Excel 2010 and SAS ver. 8.1 (SAS Institute, Cary, NC, USA) were used to examine the variation in phenotypic traits, including the mean value, standard error, amplitude, and coefficient of variation (CV). Differences in phenotypic variables among and within populations were determined using analysis of variance (ANOVA) with Duncan’s multiple range tests for multiple comparisons. A
p-value for the ANOVA
F tests ≤ 0.05 was considered significant. Pearson’s correlation coefficients for all phenotypic traits—which can guide multiple-trait selective breeding—were analyzed at the levels of the individual, clone, and population. Correlations between phenotypic traits and geographical factors (longitude and latitude) were also tested at the population level. Repeatability for all traits in each population and the overall population was calculated using the formula: R = 1 − 1/F. A linear model (
yij =
u +
ai +
bj +
eij) was used to generate the components of variance of all traits in this resource by restricted maximum likelihood (REML) in the open source statistical package R [
16], where y
ij is the dependent variable, u is the overall mean,
ai is the random effect of population
i (
i = 1, ...,
m ),
N(0, б
a2),
bj is the random effect of clone
j (
j = 1, ...,
n),
N(0, б
b2), and
eijk is the residual (
N(0, б
e2)). All models assumed that the random effects were distributed normally with expectation zero and corresponding variances. V
E = б
a2, where V
E is the estimated environment variance, V
G = б
b2, where V
G is the estimated genetic variance, and б
e2 is the residual. Realized gain (G) was estimated by the formula G = (
Xi −
X)/
X × 100%, where
Xi and
X are the mean values of the selected elite clones and the overall trait, respectively.
4. Discussion
In 2004, Chinese fir from six provinces in southern China were grafted in Longshan State Forest Farm, and all were grown under the same environmental conditions. As it can maintain a recurring tree phenotype, grafting has long been used to produce elite tree clones for conservation and breeding, as well as provide evidence for a genetic effect in traits [
14,
17]. Compatibility is vitally important in grafting, and sometimes a species grafts more successfully on stock of the same species [
18]. In this study, all of the stock was two-year-old root segments of Chinese fir, and high grafting success (>90%) was seen, with no graft incompatibility. Moreover, most of the ramets remained vigorous throughout the study, as in a previous study [
14]. The high repeatability of all measured traits in this study indicated that the population was under moderate genetic control and that the selection was effective, which is also supported by the average repeatability estimations of 0.65 in wood properties in trees [
13,
14,
17]. Studies of other Chinese fir in Longshan State Forest Farm have also shown that the local environment has little effect on them [
13,
14]. Thus, the phenotypic variation observed in this study may reflect a larger degree of genotypic variation [
13,
17].
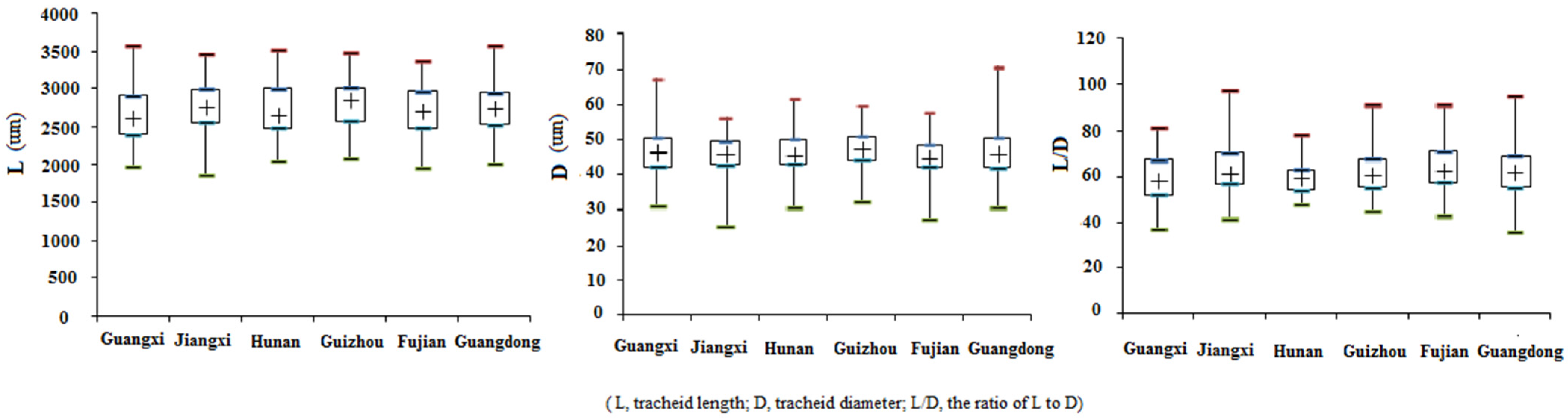
Growth traits and wood properties reflect the performance of wood. In this study, V revealed the yields of wood production, illustrating the superiority of the clones. The high CV (77.63%) for V revealed abundant phenotypic diversity in V among all individuals from different populations (
Table 1). One of the most important wood properties is WBD, which represents the degree of wood compactness and affects the quality of nearly all wood end-products [
19,
20]. P, which also plays a critical role in the improvement of wood material, was positively associated with WBD. Hy [
14], another wood trait that has multiple effects on end-products, was also tested in this experiment. Tracheids are the most abundant conducting structures in the xylem of conifers, and the morphology of tracheids significantly influences the properties of paper. Wood with a greater L/D is better for making paper [
21,
22]. There were similar high D values among the populations, but differences in L, resulting in L/D differences among the populations (
Table 3). Among the populations, Fujian had the largest average L/D. In addition, an ANOVA analysis indicated that most other traits showed moderately significant variation and that genetic parameters varied with different populations (
Table 1,
Table 2 and
Table 3). Further ANOVA analysis within populations showed that there was abundant variation among the clones, which provides potential opportunities for genetic improvement of Chinese fir with large potential for selection. Chinese fir is indigenous to southern China. The trees investigated in this study were all from subtropical climate zones, which may weaken their genetic structure to an extent. Further analysis of the trait-geographical factor correlations at the population level (
Table 8) showed that most traits had no significant correlation with longitude or latitude. However, it is important to characterize the variation in these traits in response to geographical environmental gradients in order to understand the evolutionary adaption of Chinese fir. Much more study of greater scope is needed.
The most important traits of trees are quantitative traits, and either strong linkage disequilibrium of related genes or pleiotropy effects explain the common correlation between two quantitative traits in a natural population. The correlations among growth and wood properties of Chinese fir are important for its genetic improvement. The correlation analysis of all 10 phenotypic traits in this study showed that there were strong correlations for each trait at both the clone and individual levels. All of the growth traits showed significant positive correlations with each other, as well as with P, Hy, L, and L/D, showing that they may have similar genetic bases and may be biologically related [
17]. There were negative correlations between WBD and all of the growth traits (which is consistent with the results of previous investigations [
13,
14]), and DBH showed the highest degree of negative correlation. At the population level, WBD also had a significant negative correlation with DBH. Although WBD had significant negative correlations with most of the traits, it is one of the important indicators of wood quality. Rapid growth with relatively high WBD increases the value of the output [
23], and seems to represent the correlated response of traits to selection. Thus, considering the DBH and WBD among all clones, 98 relatively fast-growing clones with relatively high WBD were identified. The gain in DBH was 14.80% and that in WBD was 8.82%. In addition, there was a 6.65% gain in V. There was a positive correlation between Hy and growth, and a strong negative relationship between Hy and WBD, resulting in reductions (−10.77%) in Hy. Therefore, it is desirable to use only a few traits as a selection index in a selective breeding program [
24]. The traits of Chinese fir can be predicted early [
25], as the juvenile wood and mature material properties are closely related. A previous study showed that it was feasible to predict WBD and L of mature wood using a density of around 10 in juvenile wood [
26]. These results suggest that our selection is reliable and that it provides a basis for the prediction of mature wood material.
5. Conclusions
Variation is essential for the adaptability of a population and is the basis for the evolutionary potential of a species. In our study, 10 phenotypic traits were investigated and abundant variation was found with the phenotypic variation coefficients all exceeded 10%. The variation in most growth traits was significantly different among the populations. Trees from Guangxi, Guizhou, and Guangdong had better growth performance. All traits differed significantly within populations and had high value of repeatability, providing possibility to choose plus trees. The correlation between characters is important in the selective breeding program and the correlation analysis of all 10 phenotypic traits in this study showed that there were strong correlations for each trait at both the clone and individual levels. All of the growth traits showed significant positive correlations with each other but negative correlation with WBD, and DBH showed the highest degree of negative correlation. However, rapid growth with relatively high WBD can increase the value of the output. Thus, using DBH and WBD as criteria, 98 relatively fast-growing genotypes with relatively high WBD were identified. And the gain in DBH was 14.80% and that in WBD was 8.82%. Overall, the results will improve the application of this germplasm resource.






{kind=link}
{kind=link}