1. Introduction
Practices that maintain or enhance biodiversity and structural complexity in forest ecosystems may increase adaptive capacity [
1], an increasingly emphasized management goal in light of ongoing environmental change [
2,
3]. Thus, the development, evaluation, and adaptation of approaches that conserve those elements while continuing to provide forest products are rising in importance. Retention harvesting, an approach that intentionally retains legacy features such as single, mature, overstory trees or undisturbed patches (aggregates), provides options for achieving that balance across ecosystems [
4,
5], yet much remains to be learned about the long-term effectiveness of retention harvesting in different ecosystems and potential trade-offs with meeting tree regeneration objectives [
6].
Natural disturbances result in greater young forest structural complexity compared to even-aged regeneration methods such as clearcutting [
7,
8]. Residual live trees (as well as other structural and biological attributes that persist after disturbance) provide continuity both temporally between forest generations and spatially across a landscape. Such complexity and continuity positively influence many aspects of biodiversity [
5,
9,
10] through provision of habitat for birds and mammals [
11], maintenance of ectomycorrhizal communities [
12] which influence nutrient availability and cycling, amelioration of the microenvironment [
13,
14], and other mechanisms. Accordingly, retention harvesting systems that reserve mature, overstory trees after harvest have been developed to provide similar habitat, complexity, and services as natural legacies [
4,
7,
8]. In addition to positive impacts for biodiversity evident in the short- and medium-term [
5,
12,
15], model simulations of stand development over periods of up to 240 years suggest structure in at least some actively managed forests (i.e., in the Pacific Northwest), will be more similar to natural forests if harvested within retention systems rather than clearcutting [
16].
Retention harvesting practices aim to maintain integrity, resilience, and diversity of forests while also providing forest products [
17,
18,
19]. However, application may also involve trade-offs with short-term productivity and specific regeneration goals [
20]. Retained overstory trees can result in greater competition for light [
21,
22], water, and nutrients [
23] which can have a direct negative effect on the establishment and growth of regenerating tree species [
24,
25,
26,
27,
28]. While competition for light may influence growth regardless of shade tolerance [
21], species such as
Populus tremuloides Michx. might be impacted to a greater degree both directly, because of high intolerance of shade, and indirectly, through cooler soil temperatures known to reduce root sucker initiation and annual growth [
29]. On the other hand, retained overstory trees may ameliorate the microenvironment [
13] and improve regeneration survival, facilitate access to mycorrhizae [
30], or indirectly improve growth of
P. tremuloides regeneration by reducing competition from other species such as shrubs [
31]. Understory shrubs are known to limit tree regeneration success and growth across many forest ecosystems [
32,
33,
34], particularly for shade-intolerant species [
10], and can retard succession [
31]. The effects of overstory retention on the shrub layer likely depend on site factors and the composition of the shrub community; observations in aspen mixed-conifer stands suggest that overstory retention in partial harvests decreases growth of shrubs [
35], but without necessarily conferring any benefit to target (crop tree) species [
35]. Overall, mixed results have been observed in aspen-dominated ecosystems [
36,
37] and limited information exists on aspen response and regeneration dynamics 10 years or more after partial harvest or harvesting with long-term retention (but see [
31,
38,
39]).
Understanding the biophysical effects of retained overstory on surrounding harvested areas (“forest influence” [
40,
41]) and its temporal scale [
40] is integral to achieving the ultimate goals of continuity, recolonization, and structural complexity associated with retention harvesting [
28,
42]. It is generally assumed by forest managers that forest influence is greatest within a distance approximately equivalent to the height of the dominant trees in a given forest ecosystem [
28,
41] even though evidence suggests a high degree of variation, depending on the variable of interest, time since harvest, and forest type [
42]. Across biomes, forest influence on microclimate generally extends 10–20 m into harvested areas [
42], with distances up to 44 m observed in temperate Tasmanian forests [
43], and can persist for as long as 45 years in some systems [
14]. Observations from boreal forest suggest a more limited distance of influence on understory structure than other ecosystems, with effects only extending 5–15 m into harvests [
44]. This might, in part, be explained by the shorter stature of trees in this ecosystem.
The smaller patch size of retained aggregates may decrease the forest influence provided by those aggregates over time relative to intact forest. Close proximity to harvest edges can limit the longevity of overstory trees because of increased exposure to wind and other stressors, particularly in aggregated retention where retained trees are more exposed [
45,
46,
47]. For instance, windthrow occurs more frequently and up to greater depths in retained aggregates relative to larger intact forest stands in temperate forests of Tasmania, Australia [
47]. Similarly, observations in Canadian boreal mixedwoods suggest higher rates of mortality in overstory trees in cut areas following partial harvests compared to intact stands, both as a result of stress from exposure (primarily hardwoods) and windthrow (mostly conifers) [
46]. This mortality can contribute downed, dead wood and influence the coverage and exposure of mineral soil, all factors determining the availability of appropriate substrate and conditions for seedling germination. The survivorship of mature trees also affects seed availability, seed dispersal (as species differ in their response to edge conditions [
46]), and the microclimate. Thus, seedling germination and establishment are subject to forest influence and differ according to the arrangement and proximity of mature overstory [
48] at short- to medium-terms after harvest [
10]. Moreover, while mostly similar between aggregates and intact forest, some evidence suggests a greater depth of microclimatic influence into harvested areas associated with intact forest [
43]. This could be related to greater sunlight penetration of aggregate edges (and therefore less shade) resulting from the mortality discussed above, a lesser degree of shade over the course of a day because of sun positioning and smaller patch size, differing influences on airflow, or a combination of the above [
43]. Overall, understanding forest influence as it relates to overstory retention is key to quantifying potential trade-offs and evaluating the success of retention harvesting practices at achieving balance among ecological, economic, and social objectives.
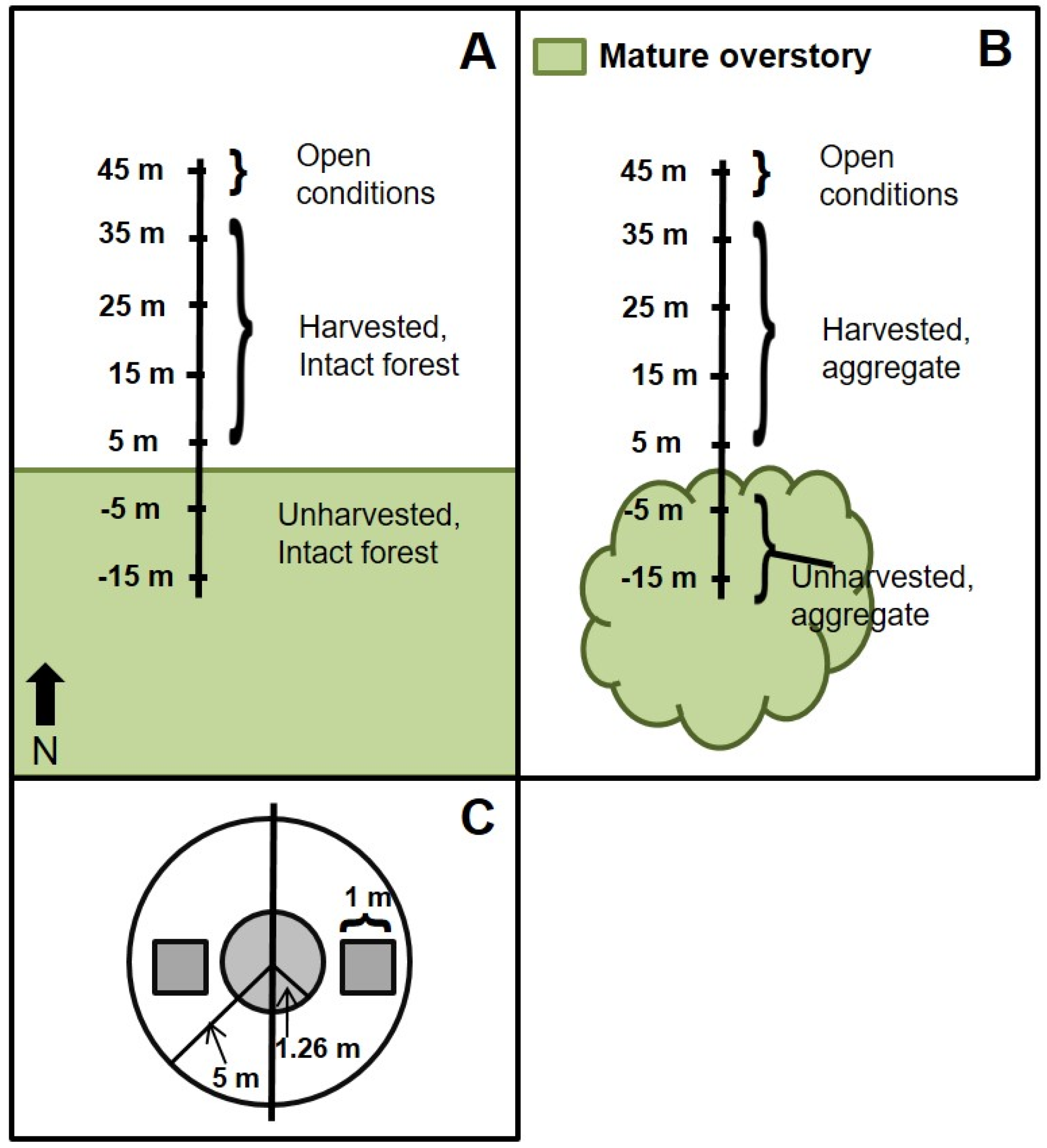
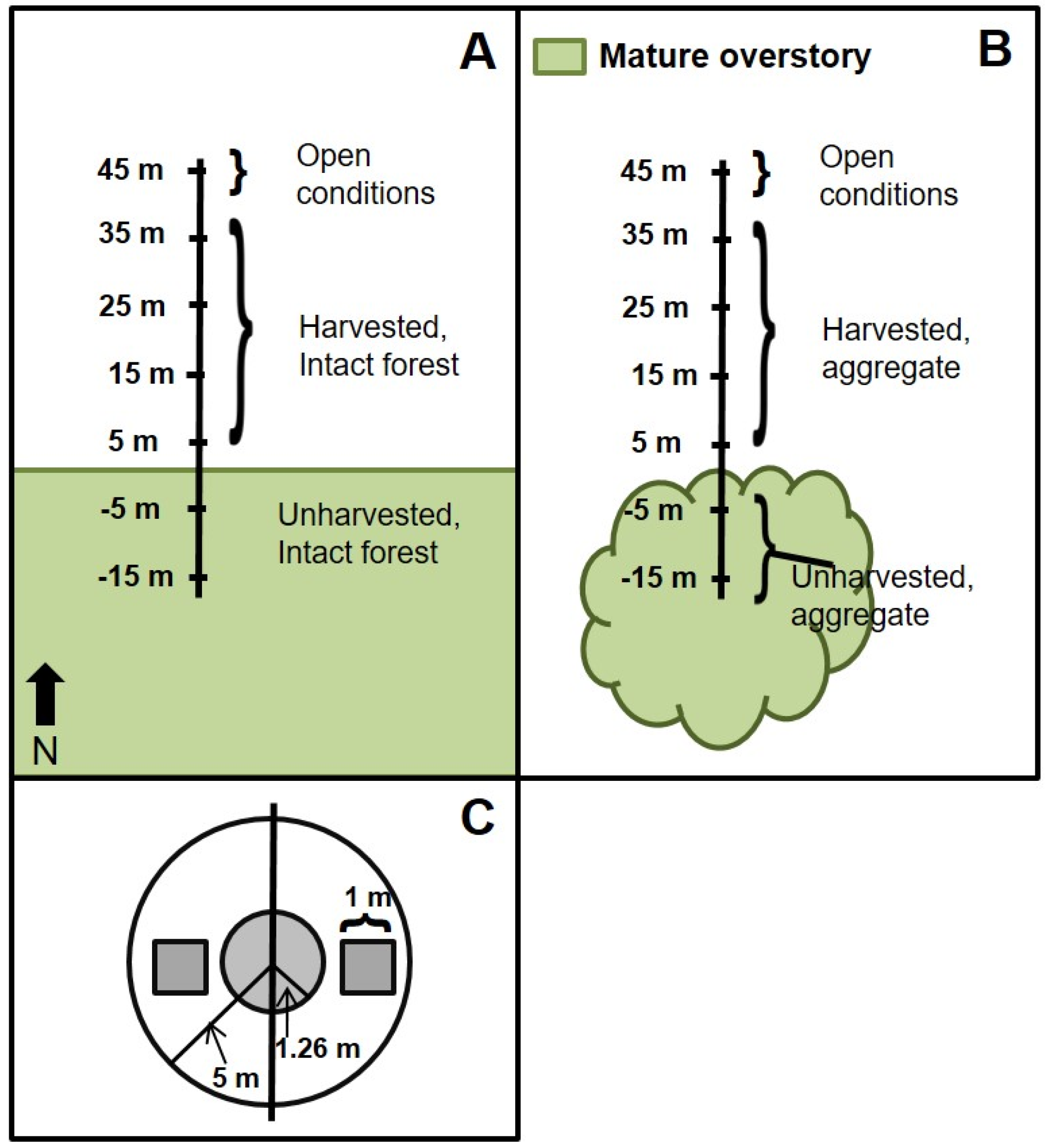
Our study assesses the influence of aggregated retention on the structure and productivity (standing biomass) of tree regeneration and the woody understory. This was accomplished using a dataset that includes replicates from multiple sites dominated by
P. tremuloides, a species that regenerates prolifically via root sprouts following disturbance such as harvest [
29]. Based on observations of forest influence on general plant species composition [
49] and the functional diversity and functional composition in the woody understory of the same
P. tremuloides-dominated forest ecosystem (Curzon et al. unpublished), as well as observations of forest structure in boreal forest [
44], we hypothesized that intact forest and aggregates would both influence structural composition of adjacent regenerating forests. We also hypothesized that the magnitude and extent of forest influence into regenerating forest would be greater adjacent to intact forest edges relative to small, retained aggregates.
4. Discussion
Overall, fewer differences between the effects of intact forest and aggregated retention were observed relative to expectations despite the small size of aggregates (0.1 ha) included in this study. Instead, distance from harvest edge (up to 5 m into harvests) and whether a plot was harvested or unharvested had more influence on structure and standing biomass of regeneration and the woody understory, regardless of whether transects were adjacent to intact forest or aggregates. Yet, even given these similarities, analysis of depth of edge influence suggests that forest influence exerted by aggregates resulted in fewer significant effects on structural variables indicative of regeneration success (i.e., large sapling density and total understory biomass). These findings have implications for managers who are interested in implementing retention practices but are also concerned about losses to productivity of the regenerating forest.
The general similarity in influence on the structure of regeneration that occurred between aggregates and intact forest is consistent with observations of community composition in vascular plant and beetle species on the same sites [
49]. These results are also consistent with comparisons of the extent of microclimate effects between aggregates and intact forest reported for cool, temperate
Eucalyptus forest in Tasmania three to four years after harvest [
43]. While forest influence may vary across a harvest with differences in aspect, we positioned transects as consistently as possible to maximize the influence of shade from mature, overstory trees on surrounding harvested areas. If a substantial difference in the mortality of overstory trees near the harvest edge does not exist between aggregates and intact forests, then neither would the resulting shade and influence on the microclimate during the middle of the day when shading would be expected to be most similar for the two patch sizes given sun position and have the greatest impact on ameliorating microclimatic conditions [
43].
We also suspect that the rapid development of regenerating harvested areas, particularly given the dominance by
P. tremuloides, a prolific and fast-growing species [
29], has lessened the contrast between mature forest and regenerating harvests (as suggested in [
44]) leading to fewer differences in the microenvironment and in resource availability between mature and regenerating forest that may have existed previously. The distance of influence we observed associated with both overstory conditions is shorter than reported for other regions over periods of up to 20 years (i.e.,
Psuedotsuga menziesii-dominated forests in Washington, DC, USA; [
15]). However, it is consistent with evidence from studies in boreal and sub-boreal forests more similar to those assessed here that suggest that the response of regeneration growth and density to proximity to mature overstory trees may generally extend short distances [
44]. The lower stature of these forests (mean mature overstory height of 17.4 m across the study) may contribute to the shorter distances of influence relative to other systems such as those dominated by
P. menziesii where a moderate site index at 100 years ranges 32–35 m [
59] and in mature temperate eucalypt forests with overstory tree heights of 35–50 m [
49].
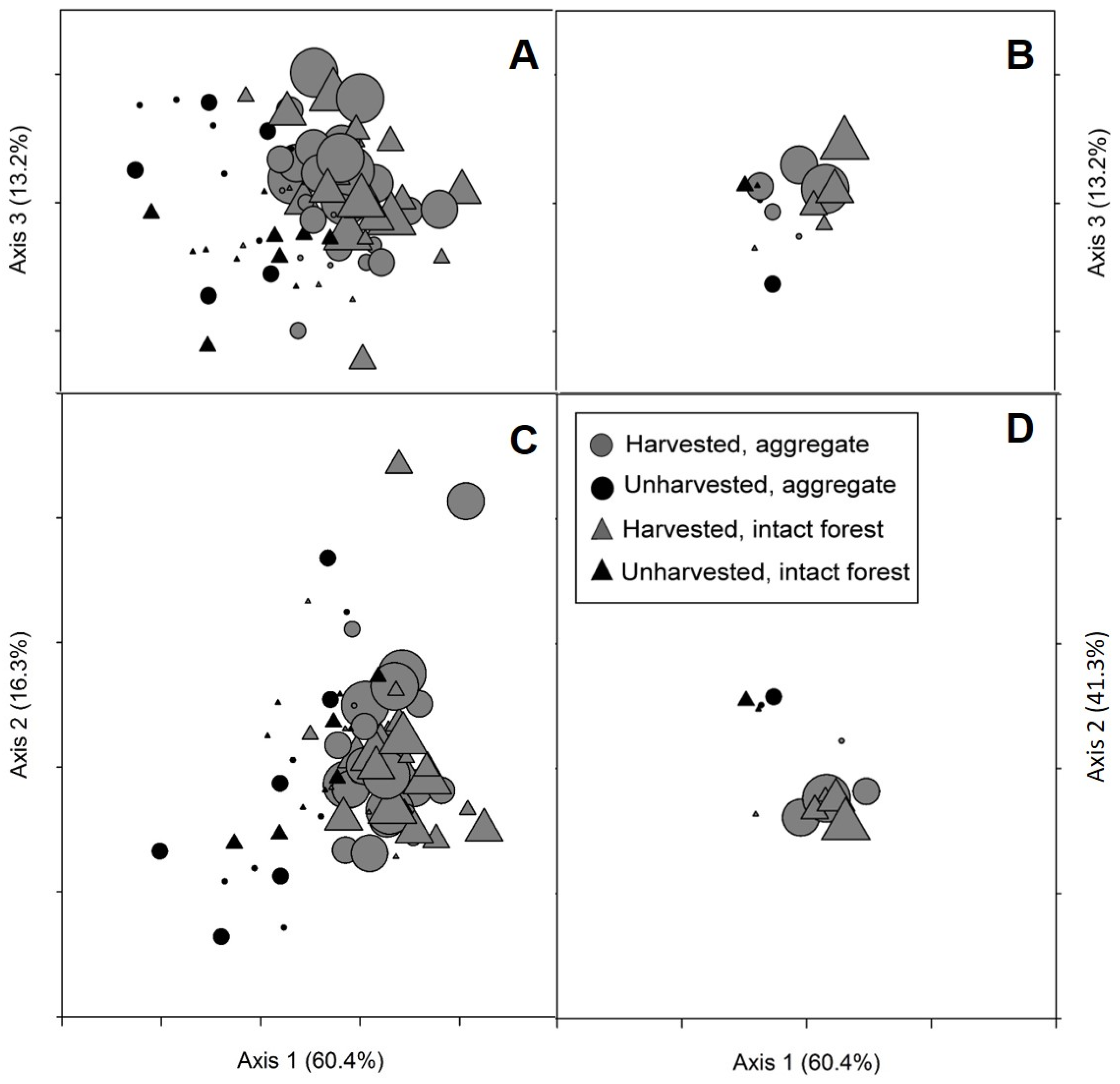
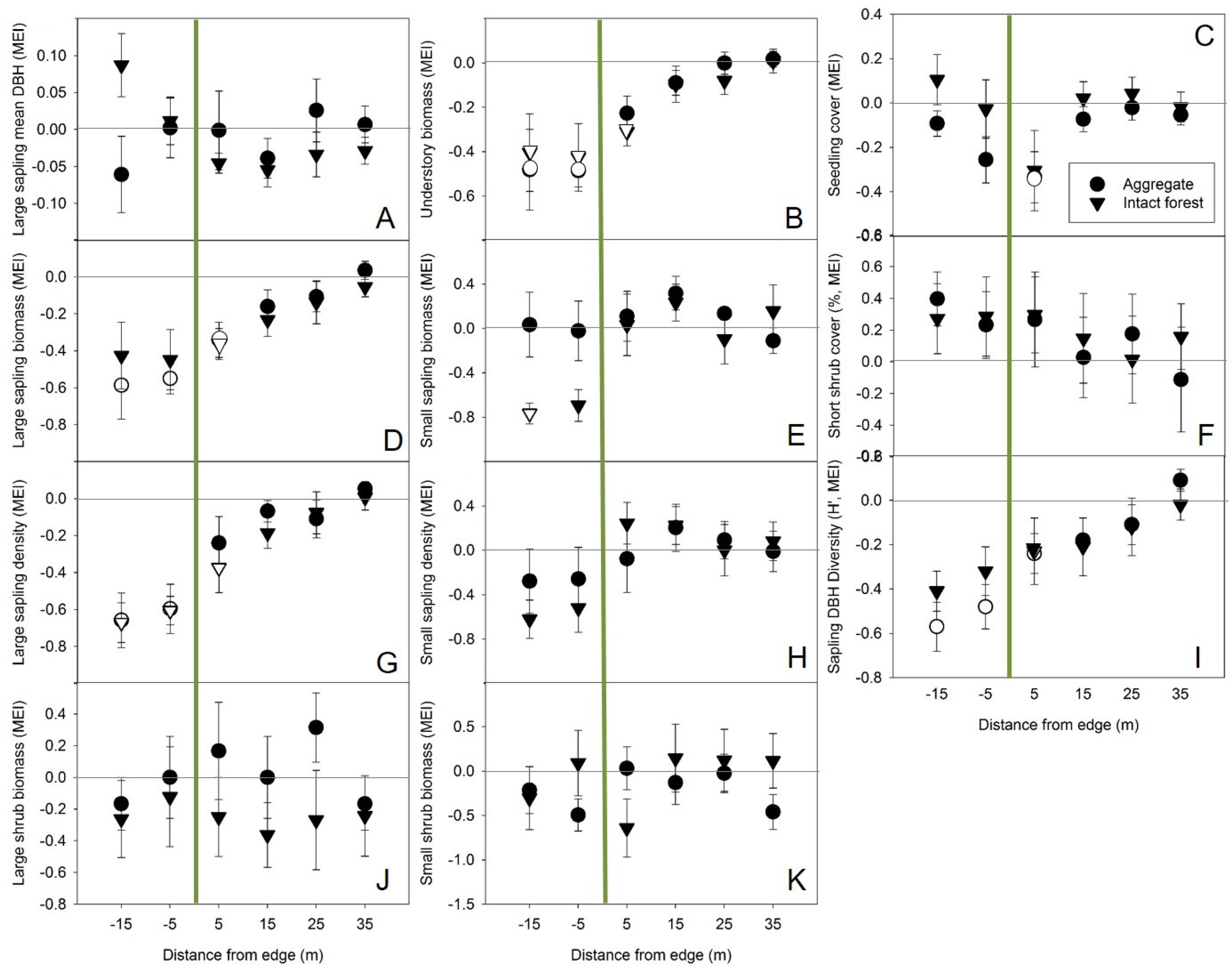
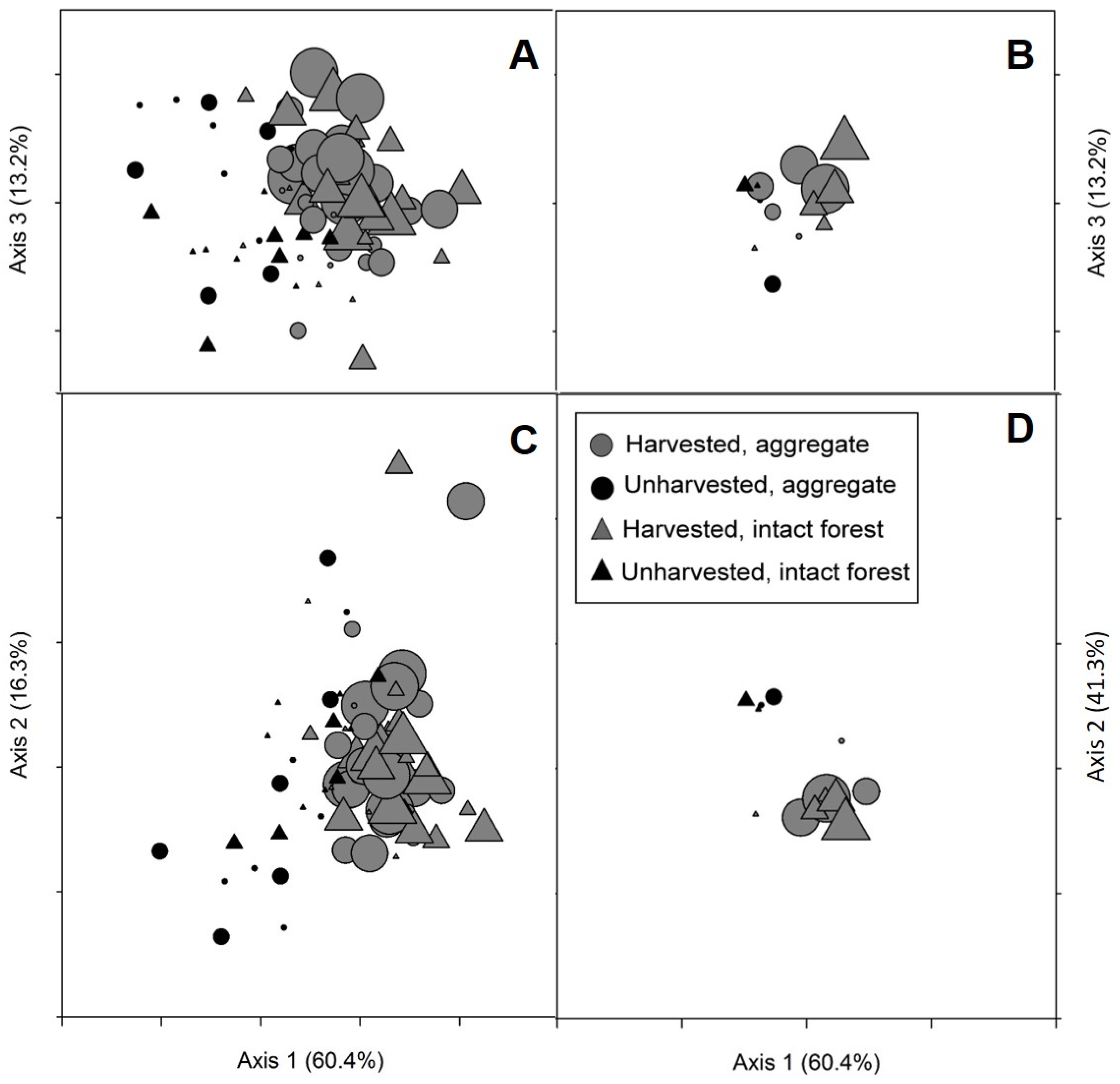
Variability within and across sites as well as other variables not included in these analyses appeared to be more important drivers of differences in sapling densities and biomass than either overstory condition of nearby retention (aggregate vs. intact forest) or distance from edge (
Figure 2,
Table A2). Although a reduction in total large sapling biomass was observed in harvested areas immediately adjacent to aggregates, the density of large saplings in total, and the density and biomass of large
P. tremuloides saplings specifically, was variable and not significantly different from open conditions at all distances from edge.
P. tremuloides abundance also showed only weak relationships with the two NMS axes associated with distance from edge and overstory condition (Axes 1 and 3, respectively). These results contrast with reductions in stem density and biomass observed in
P. tremuloides-dominated systems with 40% basal area retained [
38]. However, our results align with findings that retention level (up to 50% basal area retained) had little effect on total or specifically aspen/birch sapling densities [
22] and that stem density may be less impacted by retained overstory trees than growth rates [
27]. Comparable eucalypt densities and growth rates between harvests with aggregated retention compared to clearcuts also suggests negative impacts on crop tree regeneration may be less of a concern in some systems [
60]. The negative forest influence on sapling diameter diversity observed here, while minimal, suggests that overstory retention may retard structural development [
60].
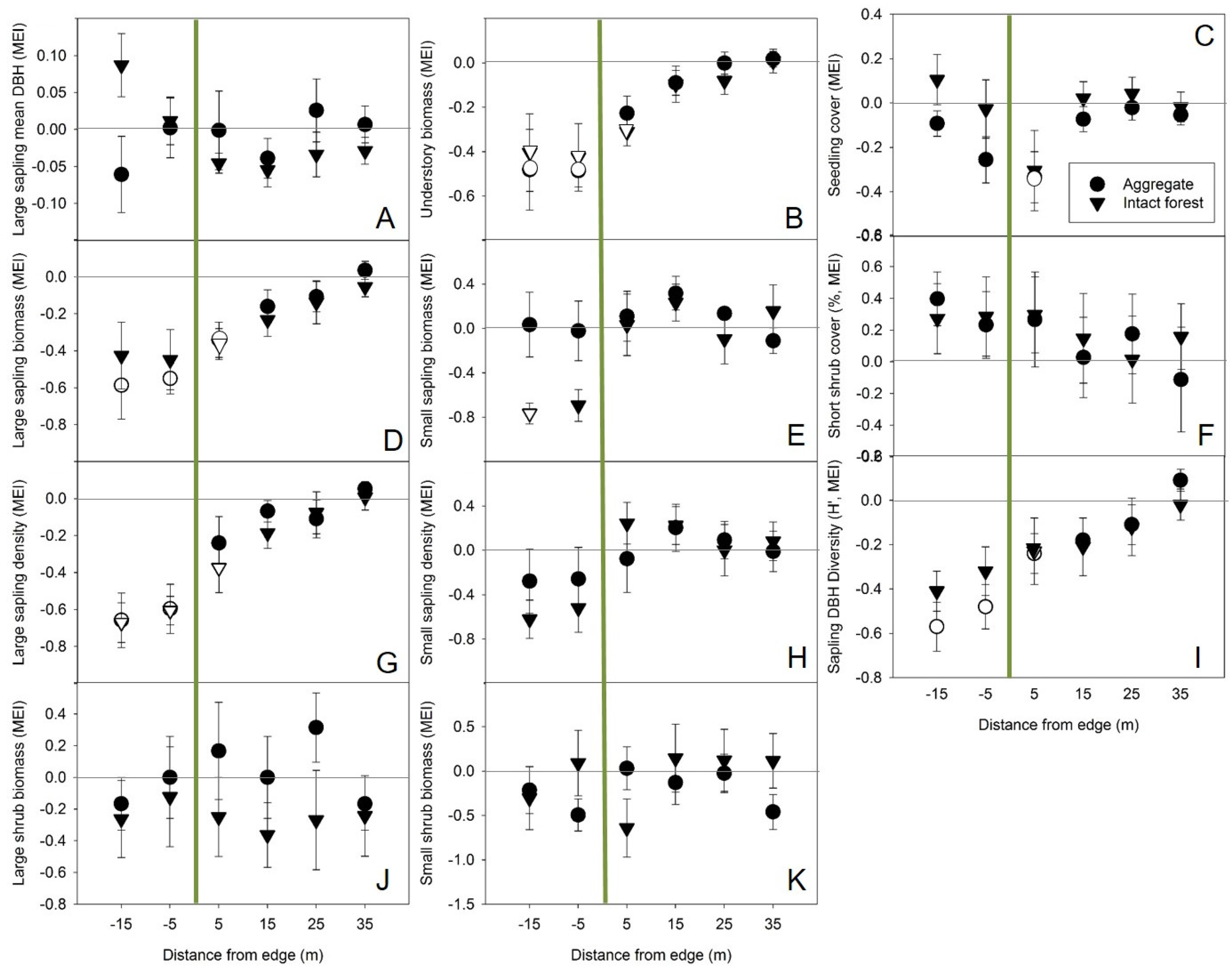
Shrub biomass immediately adjacent to aggregates, while not found to be significantly affected by forest influence, may have been high enough on average to make up for any reduction in sapling biomass contributing to the overall standing biomass pool (
Figure 3). Aggregates had a negative influence on seedling cover up to 5 m into regenerating harvest areas, but negative correlations between seedling cover and multiple measures of shrub abundance suggest this could have been an indirect effect resulting from the level of competition exerted by shrubs which were relatively high in density, biomass, and cover near the harvested side of those edges (
Figure 3). As observed in boreal aspen-mixed woods [
35], retention did not appear to reduce competition from shrubs in nearby harvested areas. In fact, interpretation of the lesser difference between observed plots and reference (open, harvested) plots near aggregates along Axis 3 (positively correlated with small shrub abundance) suggests the possibility that aggregates had a lower negative influence on shrubs than intact forest.


 ,
, 


{kind=link}
{kind=link}
{kind=link}