1. Introduction
Eucalypts are one of the most important planted hardwoods worldwide [
1]. The remarkable characteristics of this species, including their rapid growth, straight form, valuable wood properties, and wide adaptability, have driven them to be planted over 20 million hectares [
2,
3], having provided key renewable resources for the production of pulp, paper, biomaterials, and bioenergy in more than 100 countries across six continents [
4,
5,
6,
7]. In addition, they also have high concentrations of diverse essential oils for medicinal and industrial uses [
8,
9]. As a result of many years of study, great achievements have been made on the genetic improvement of this species. However, there is still room for improvement, especially in regard to the creation of new germplasm in this tree species.
Polyploid plants are widely distributed in nature [
10,
11,
12]. Previous research has shown that polyploid varieties with high productivity, high product quality, and strong resistance to stress play an important role in crop and horticultural plant production [
13]. Similar observations have been made in the field of forestry, as polyploid breeding, particularly triploids induced by sexual polyploidization, has great potential in improving vegetative growth and economic traits. In recent years, triploid breeding has achieved great success in genetic improvement in
Populus. Compared to their diploid counterparts, triploid
Populus showed marked advantages, such as a faster growth rate, larger leaf organ, higher photosynthetic rate, better timber quality, and higher stress resistance [
14,
15,
16,
17,
18,
19,
20]. For similar general purposes, triploid breeding plans should immediately become a part of the eucalypt genetic improvement program.
Triploids can be obtained through hybridization using diploid and natural polyploids [
14] or by pollinating with natural [
21] or artificial 2n pollen [
22,
23,
24]; however, this method would be inefficient due to the weak development and competition of 2n pollen compared to normal pollen [
25]. Recent developments in forestry breeding research demonstrated that female flower buds that underwent a colchicine treatment or were exposed to high temperature to induce megaspore or embryo sac chromosome doubling were a more effectively way to produce triploids, and the yield of triploids reached 16.67%–66.67% [
26,
27,
28,
29,
30,
31,
32,
33]. This study provides some guidelines for triploid breeding induced by gamete chromosome doubling in other similar tree species.
In
Eucalyptus, there are no reports that indicate the existence of natural polyploids [
34]. While tetraploid plants have been artificially induced in vitro via colchicine treatment [
35,
36,
37], their development and application are still insufficient due to the difficulty of chimaera separation and the poor performance of the autotetraploid. We previously reported a method to obtain 2n pollen via colchicine treatment during microsporogenesis in
Eucalyptus urophylla S.T. Blake with an expectation to produce triploids [
38], but major problems, such as low yield and the purity of 2n pollen, limited its uses. Fortunately, these attempts uncovered some basic technical requirements, such as potential suitable periods for gamete chromosome doubling, which laid some foundation and gave some hope for further developments in eucalypt triploid breeding.
This study aims to explore a technique to produce triploids through female gamete chromosome doubling in Eucalyptus. Cytological observation on microsporogenesis and megasporogenesis were carried out to guide megaspore chromosome doubling to investigate the possibility of triploid induction by colchicine treatment using two different methods of application.
2. Materials and Methods
2.1. Plant Materials
Floral branches used in this study were selected from a Eucalyptus urophylla clone (2n = 2x = 22), which contains 10 individual trees. This clone was planted in a clonal test plantation at the Guangxi Dongmen Forest Farm (Guangxi Zhuang Autonomous Region, China), which was built in 2008.
2.2. Determination of the Developmental Process of the Megasporogenesis
Eucalyptus flowers were observed in a cluster in an inflorescence born in the axil of a leaf, and the unit inflorescence was generally a simple umble with 5–7 single flower buds [
2]. According to previous studies [
39,
40], the development of flower buds at different locations on a floral branch was generally asynchronous, and the relationship between microsporogenesis and flower development could be used to estimate the period of meiosis [
38]. In this study, flower buds were sampled every 12 h when the flower bud grew to 3.0 mm in diameter in the selected floral branches until fertilization from June to August in 2016. Flower bud samples were fixed in FAA (formalin–acetic acid–alcohol) fixative (70% ethanol/acetic acid/40% formaldehyde, 90:5:5) at 4 °C for 24 h. The anthers containing cells undergoing meiosis from each flower bud were extracted and stained with 2% acetocarmine for microsporogenesis observation. The ovaries from the same flower bud were embedded with paraffin, then sectioned at 8–10 μm and stained with iron hematoxylin for megasporogenesis observation. After cytological observations, the determination of the developmental process of the megasporogenesis was based on their relationship, which could be used to guide the colchicine treatment.
2.3. Colchicine Treatment
Floral branches, which were selected based on the determination of the developmental process of the megasporogenesis, were treated with 0.25% or 0.50% colchicine solution for 6 h from July to August in 2016. Colchicine treatments were performed in two different ways (
Figure 1). In the first way, colchicine solution was slowly aspirated from a centrifuge tube (abbreviated as ACT,
Figure 1a) into the appropriate location (a cut near the floral shoot) on a floral branch by capillary action, according to Yang et al. [
38]. In the other way, colchicine solution was slowly injected using an infusion apparatus (abbreviated as IIA,
Figure 1b) into the cut at the base of the floral branch. In addition, several untreated flower buds served as the control group. Considering the potential toxicity of colchicine, the operator should wear protective gloves and goggles while configuring the reagent or performing treatment using colchicine. When the treatment ends, the centrifuge tube or infusion apparatus with residual colchicine solution should be removed from the floral branches immediately and treated harmlessly after extensive dilution.
Matured seeds both from the treated group and control group were collected and germinated in yellow soil in June, 2017. When the seedlings grew to approximately 5 cm in height, they were transplanted to containers with nutritious soil to promote growth for further ploidy level detection.
2.4. Detection of Ploidy Level in Progeny
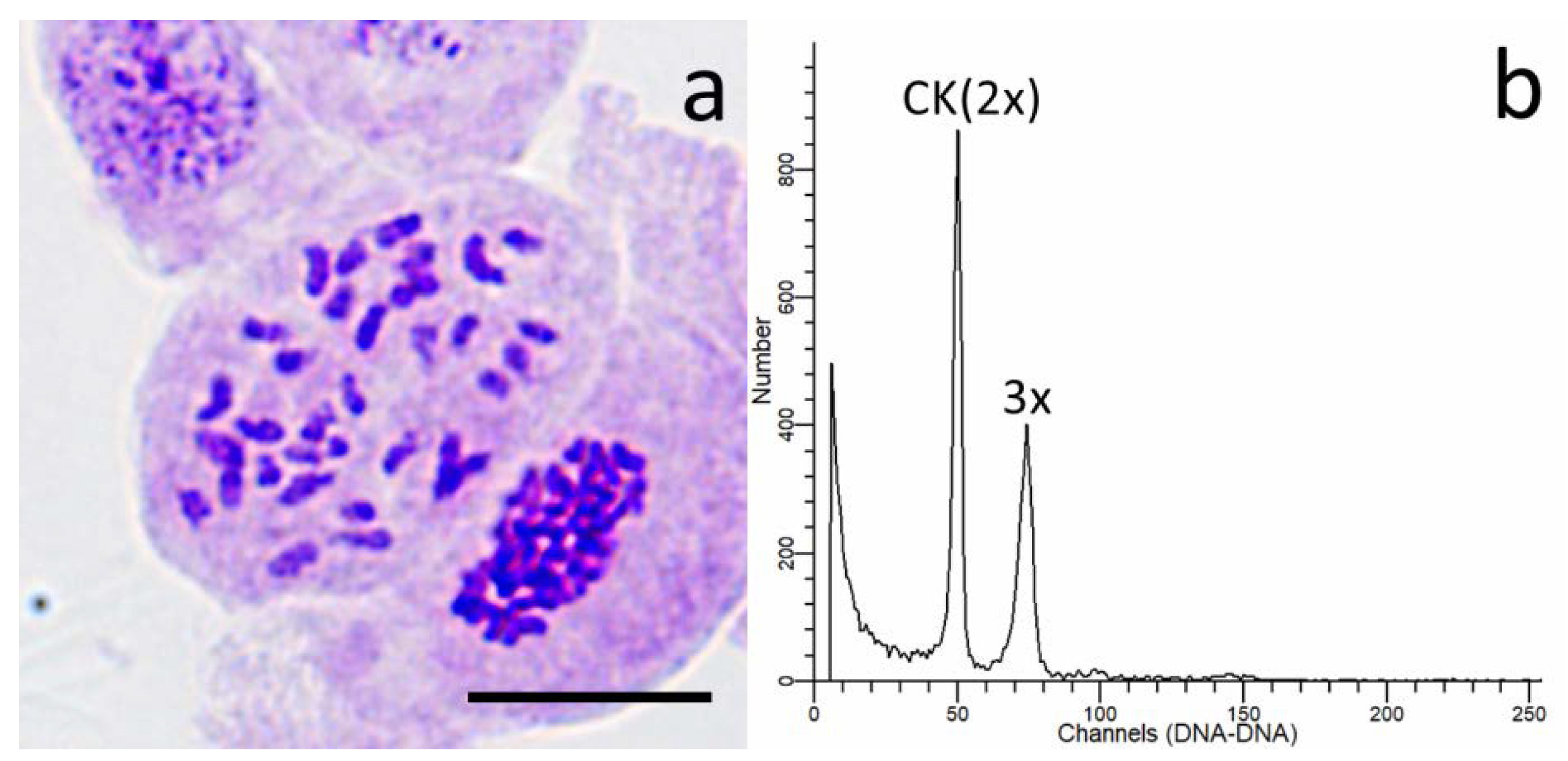
When the seedlings grew to approximately 20 cm, both flow cytometry and somatic chromosome counting were used to detect the ploidy level of offspring. Flow cytometric analysis was conducted according to Pinto et al. [
41]. In brief, nuclei were released from the cells by chopping approximately 0.5 g of young leaf material with a razor blade in Marie’s isolation buffer [
42] (50 mM glucose, 15 mM NaCl, 15 mM KCl, 5 mM EDTA Na
2, 50 mM sodium citrate, 0.5% Tween 20, 50 mM HEPES, pH 7.2). Subsequently, the nuclear suspension was filtered through a 50-μm nylon mesh to remove large debris, and then 10 μg mL
−1 of 4’,6-diamidino-2-phenylindole (DAPI) were added to stain the DNA for 5 min. Samples were analyzed by a flow cytometer (The CyFlow
® Ploidy Analyzer, Sysmex Partec GmbH, Görlitz, Germany). A known diploid plant of
E. urophylla (2n = 2x = 22) was used as an external standard.
The ploidy level of all putative triploid plants was finally confirmed through somatic chromosome counting. Stem tips were removed from the seedlings and pretreated in a saturated solution of paradichlorobenzene for 4 h at room temperature, then washed once and fixed in fresh Carnoy’s fixative (ethanol/acetic acid, 3:1) for at least 24 h at 4 °C. After that, samples were hydrolyzed in 38% HCl/ethanol (1:1) for 10 min at room temperature. After washing in distilled water three times for 15min, samples were squashed and stained with Carbol fuchsin, and then observed using a microscope (BX51, Olympus, Tokyo, Japan). Photomicrographs were acquired using the Olympus DP70 Camera System (DP70, Olympus, Tokyo, Japan).
2.5. Measurement of Phenotypic Traits of Triploid and Diploid Eucalypts
Seven diploid and seven triploid plants were used for stomata observation. Five leaves from each plant were selected for stomata observation. Mature leaves from each plant were peeled off using nail polish and then placed on a glass slide for observation, and photomicrography was performed using a microscope. The length and width of 20 stomata per leaf were measured randomly using the ImageJ software (version 1.51, NIH, Bethesda, MD, USA). Ten microscopic field areas per leaf were randomly selected to measure the stomata density. Significant differences were evaluated using t test at the 0.05 level of probability.
In the meantime, triploids and their diploid full-sib families were undergoing clonal propagation from the end of 2017. One triploid (genotype: T) and its diploid full-sib family (three genotypes: D1, D2, and D3) with 4 biological replications per genotype, which had been successfully propagated, were used for phenotypic traits measurement. In order to compare the differences between triploid and diploid plants, a total of ten phenotypic traits data were recorded when all the plants had grown up to 6 months old in the greenhouse. Height (H) and ground diameter (GD) were measured using the flexible rule. Mature leaves from the top branch of each plant were used to measure their photosynthetic parameters. The net photosynthetic rate (Pn), stomatal conductance (Gs), intercellular carbon dioxide concentration (Ci), and transpiration rate (Tr) were measured using an LI-6400-02B portable photosynthesis system (Li-Cor Inc., Lincoln, NE, USA) at 8 to 10 o’clock on a sunny day in July 2018. All photosynthetic parameters were obtained under a photosynthetic photon flux density of 1400 μmol
−2∙s
−1, at 60% relative humidity, and with the CO
2 concentration held at 400 μmol∙mol
−1 using a CO
2 injecting system [
43]. Then, the chlorophyll content index (CCI) and leaf area (LA) were recorded using a CCM-200 Plus chlorophyll content meter (OPTI-sciences Ins., Hudson, NH, USA) and CI-203 handheld laser leaf area meter (Li-Cor Inc., Lincoln, NE, USA) on the same day. The instantaneous water use efficiency (WUE
i) was calculated as Pn divided by Tr, and the photosynthetic efficiency of the whole leaf (PE
w) was calculated as Pn multiplied by LA [
19].
2.6. Statistical Analysis
Comparisons of the phenotypic parameters were evaluated based on an ANOVA with Duncan’s multiple range tests to determine whether there were any significant differences between triploid and diploid plants. Comparing the mean difference of stomata characteristics between diploid and triploid plants was calculated by paired-samples t test. All statistical analyses were performed using SPSS software (version 19.0, SPSS Inc., Chicago, IL, USA).
4. Discussion
The discovery and utilization of polyploid in forestry breeding were very early. It was 1936 when the discovery of natural triploid poplars exhibiting excellent properties was first reported [
44,
45]. Afterward, many studies looking at the artificial induction of polyploid began [
14,
21,
46], and many excellent results were obtained in the last 10 years [
27,
28,
29,
30,
31,
32,
33]. Previous research has shown that polyploid breeding is more successful upon utilization of heterosis [
47,
48,
49]. To realize the comprehensive utilization of the dosage effect and heterosis in
Eucalyptus, a more reasonable process could be to obtain triploids by sexual polyploidization. The first study on artificial induction of 2n pollen in
Eucalyptus was performed by Yang et al. [
38]. However, compared with an anemophilous flower, such as
Populus, eucalypt produced little pollen, which could not be used to produce triploids due to the low yield and purity of the 2n pollen. In this study, we successfully obtained 7 triploid progenies with heterozygosity via megaspore chromosome doubling for the first time. The stomata characteristics of these triploids were consistent with other studies [
50,
51,
52,
53]. The increased cell size as exemplified by enlarged stomata size in triploid eucalypt may play an important role in increasing the size of the leaves, which was also observed in this study [
54]. In addition, the higher photosynthetic efficiency of these triploid eucalypts may be able to explain their significant faster growth in plant height and ground diameter [
19]. Considering the outstanding growth advantage exhibited by triploid in this study, our study provides a new perspective and method for germplasm creation in
Eucalyptus, which may have a significant impact on further varietal improvement and genetic research in this tree species.
In general, the primary means of colchicine treatment includes immersing, injection, coating, etc., and the treatment method employed should be specifically designed according to the characteristics of the plant material. However, the eucalypt flower buds were coated with wax [
55,
56], which prevented the colchicine from entering the ovary via immersing or coating. The internal partition of the flower buds was compact as well, any damage to the flower bud structures could lead to death, which made it impossible to inject the colchicine solution directly into the flower bud. To alleviate this challenge, we injected the colchicine solution using the IIA method to transport the solution into the ovaries through the transfusion tissue, which differs from the directly injected method in
Populus [
26,
29,
33]. This method could treat more flower buds on the branches all at once with less mechanical damage, but the effective rate of this treatment method was low in this study because it took more time on transporting. This outcome made measuring the specific time when the colchicine arrived in the ovaries and the final concentration of the treatment inconclusive. Previous research also provided an ACT mothed to induce 2n pollen in
Eucalyptus urophylla [
38]. We also used this method to induce 2n female gametes in this study, and successfully obtained 6 triploid progenies in total. This result indicated that the ACT method was more direct and instantaneous because the colchicine was rapidly transported into the ovaries with less dissipation and dilution. It should also be noted that after sowing and seeding, the quantity of seedlings that underwent the ACT treatment group was less than that in the IIA group, and the quantity of seedlings that underwent a higher colchicine concentration treatment was also less than the quantity of seedlings that were from the lower group. These results indicated that the toxicity of colchicine on plant materials became high when treatment intensity was increased, consistent with previous studies [
29,
33,
38]. The high yield of triploid induction is not always realized through increased colchicine treatment intensity, but instead could cause shedding of flower buds, and the optimal treatment intensity should be set based on the tolerance of the plant materials in the future studies.
Previous research has shown that the efficiency of 2n gamete induction depended on whether the treatment period was suitable for chromosome doubling. Kang et al. [
22,
57] determined that the most suitable period for 2n pollen induction in
Populus via colchicine treatment was at pachytene during microsporogenesis, and the most suitable period for megaspore chromosome doubling was from leptotene to pachytene [
26]. Therefore, we needed to determine the accurate meiosis stage immediately. Derived from the relationship between microsporogenesis and megasporogenesis in
E. urophylla, on the fifth day after the time maker was determined, most of the flower buds’ meiotic division of the microsporocytes developed to metaphase I–telophase I, while the meiotic division of the megasporocytes developed to leptotene–pachytene. On the seventh to ninth day after the time maker was determined, most of the flower buds’ microsporocytes developed to the prophase II–tetrad phase of meiotic division, while the meiotic division of the megasporocytes developed to diplotene–diakinesis. These results indicated that the fifth day and the following few days would be the optimal treatment period for chromosome doubling using colchicine solution.
Results in this study also indicated that the meiotic stages at leptotene–diakinesis during megasporogenesis in the flower buds might be the optimal meiosis period for megaspore chromosome doubling by colchicine treatment, which was consistent with previous research on 2n pollen induction in
Eucalyptus urophylla [
38]. However, there were some differences with previous research on
Populus, which had found the optimal periods for megaspore chromosome doubling via colchicine was when the megasporocytes were at leptotene–pachytene [
26,
33]. The most likely reason for this difference is that the times for megasporogenesis are different across different plant species. As a result, in this study, colchicine treatment proceeded when the megasporocytes were at diplotene–diakinesis and could gain the greatest number of triploids, but the most suitable period for megaspore chromosome doubling is a topic for further research.
Colchicine is widely used for megaspore chromosome doubling. In this study, the triploid yield was lower than in previous research on
Populus [
26,
33]. Similarly, previous studies on 2n pollen induction via colchicine treatment demonstrated that the highest yield of 2n pollen in
Eucalyptus was 28.71% [
38], while the yields were 88% [
57] and 82.83% [
58] in
Populus. The most likely reason for this discrepancy is that the flower bud growth and megasporogenesis on the whole floral branch in
Eucalyptus is asynchronous [
39,
40]. While poplars can be cultivated in a hydroponic greenhouse to maintain relative consistency in the process of flower bud development, it is difficult to control the flower bud growth and megasporogenesis of eucalypt in a wild environment due to the longstanding flower bud developmental phase and fruit-setting period, which exacerbates the asynchronism because of the environmental effect. The relationship between flower bud growth and microsporogenesis among different flower buds on different locations in the floral branch has been studied [
39,
40], and the asynchronism among different flower buds may result in a lower yield of 2n pollen induction [
38]. The unique features of meiosis in microspore mother cells are probably similar to those of megaspore mother cells, which could also be influenced by the asynchronism among different flower buds on different locations in the floral branch. Therefore, it would be difficult to ensure that all flower buds within a treatment group were at the same developmental stages during megasporogenesis, thereby further reducing the triploid yields. A possible solution might be to remove flower buds in treatment groups that are significantly different in their developmental stage based on morphologic and cytological observations to make sure the rest of the flower buds developed more uniformly, which might increase the efficiency of colchicine treatment for a better yield of triploid induction in future studies.
From the present research that demonstrates high yield of triploid induction in
Populus, there is still room for improvement in triploid breeding research in
Eucalyptus through the optimization of techniques, including the timing and method of treatment. In addition, it has been reported that female flower buds from
Populus that were treated with colchicine solution after pollination by embryo sac chromosome doubling could also produce triploids [
29,
59]; this outcome presents a possible new way of triploid germplasm creation in
Eucalyptus. In recent years, high temperature treatment was widely used for triploid breeding with a higher efficiency compared to that of colchicine treatment due to a more direct effect on the 2n gamete production process [
27,
28,
30,
31,
32]. Compared with colchicine treatment, the temperature could increase quickly to save the time the colchicine transportation needed, which would enable us to distinguish the meiotic stages for chromosome doubling more accurately to increase the efficiency of triploid production. High temperature treatment could also avoid the toxicity of colchicine to the plant materials, thereby further raising the yield of triploid induction. This outcome suggests that both the megaspore and the embryo sac chromosome doubling induced by high temperature treatment is a potentially new direction for research for eucalypt breeders, as it would be a more efficient way for triploid germplasm creation in this tree species.

 ,
, 





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}