
Sendai Virus-Vectored Vaccines That Express Envelope Glycoproteins of Respiratory Viruses


Abstract
:1. The Clinical Need for Vaccines against Human Respiratory Syncytial Virus (HRSV), Human Metapneumovirus (HMPV) and Human Parainfluenza Virus (HPIV)
2. Virology of the Paramyxoviruses and Pneumoviruses
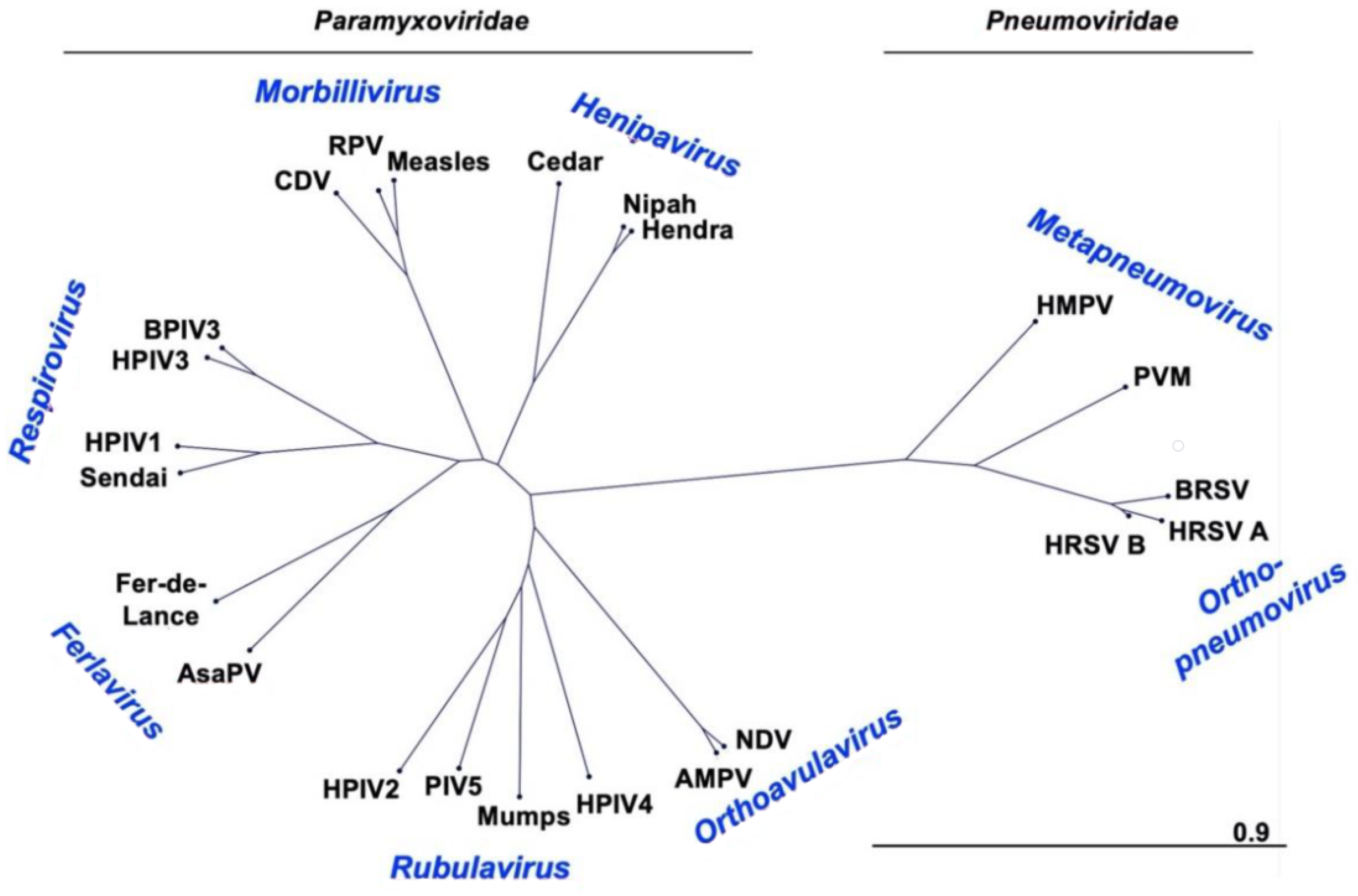
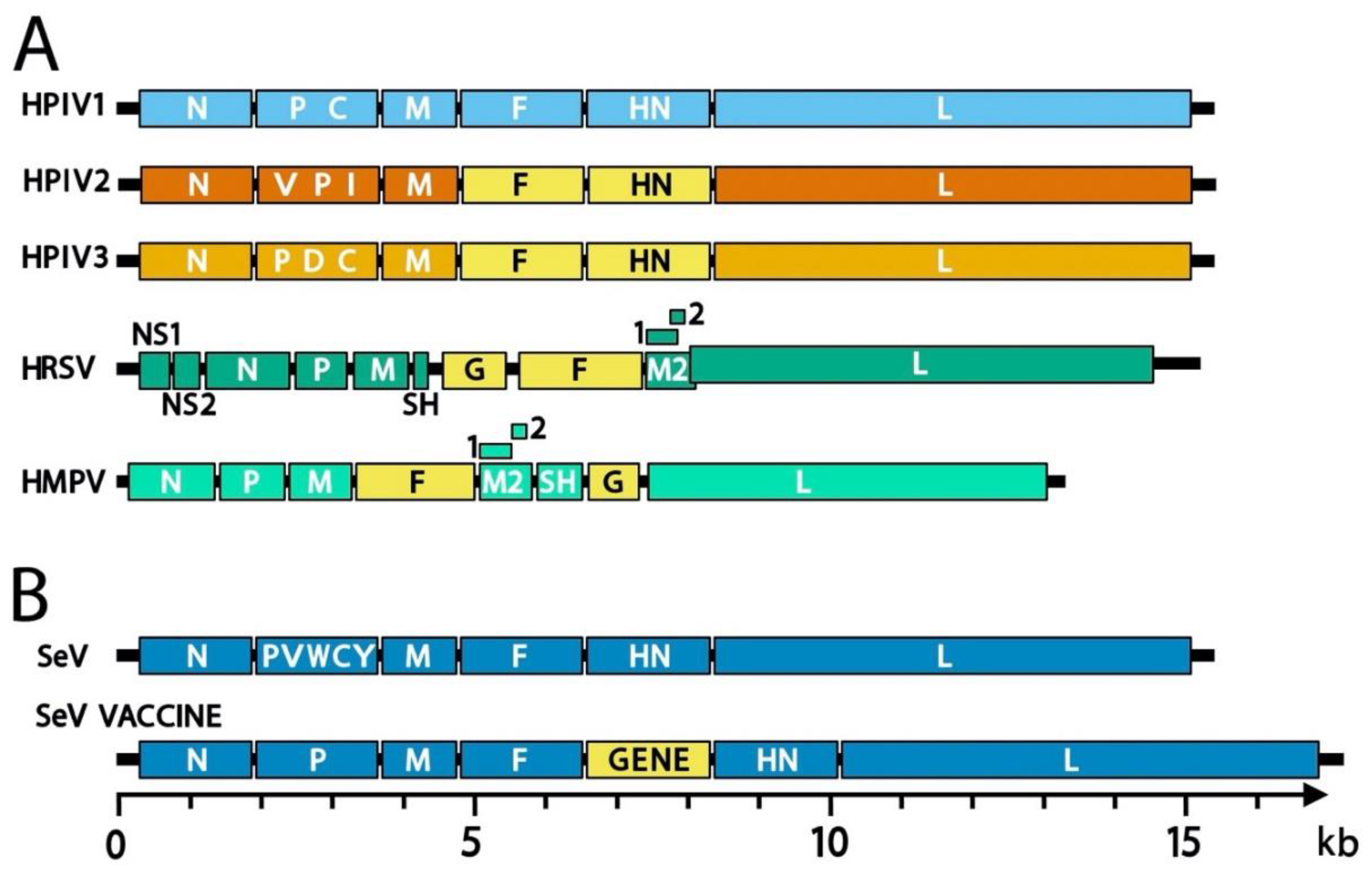
2.1. Taxonomy and Phylogeny
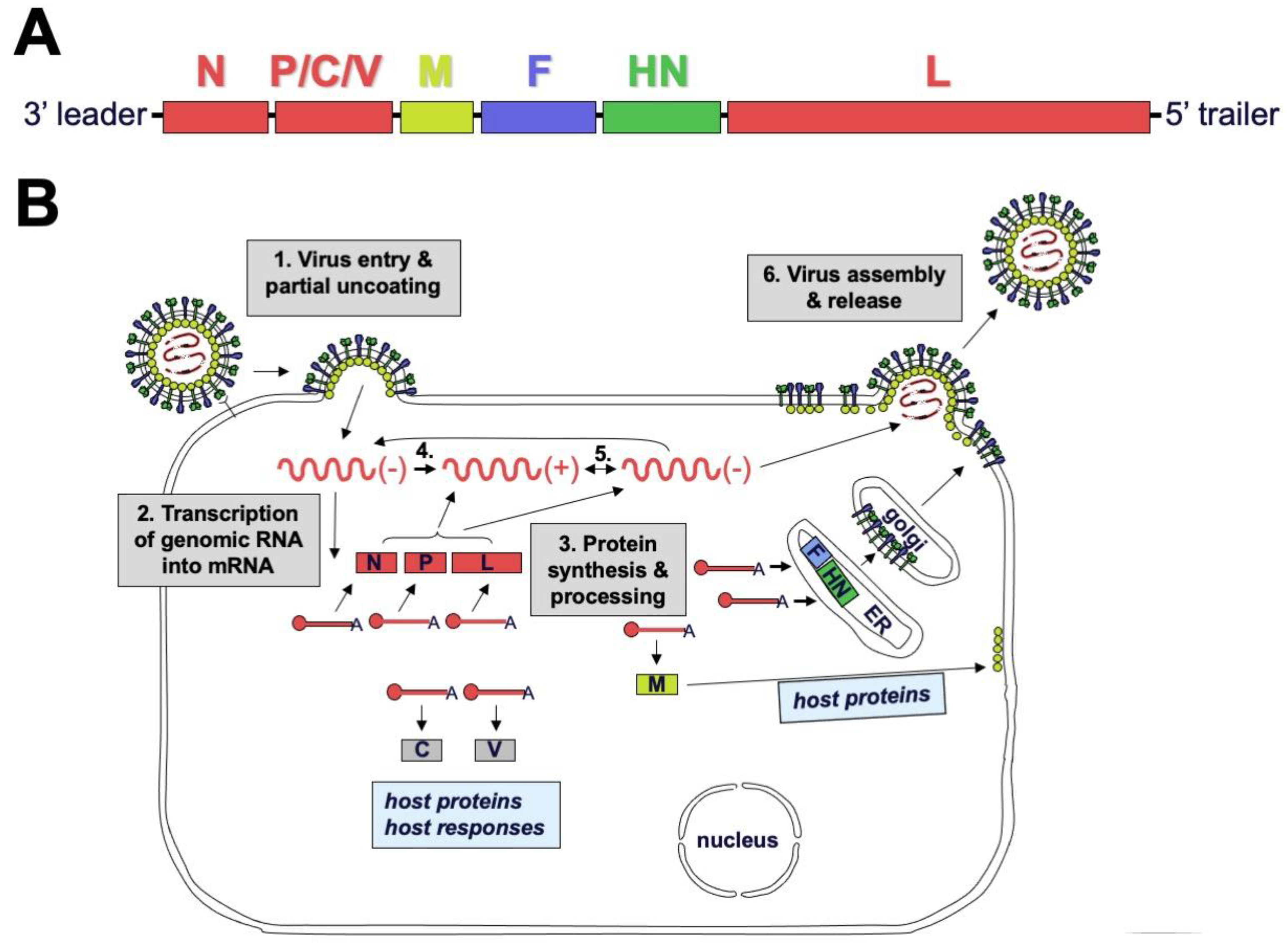
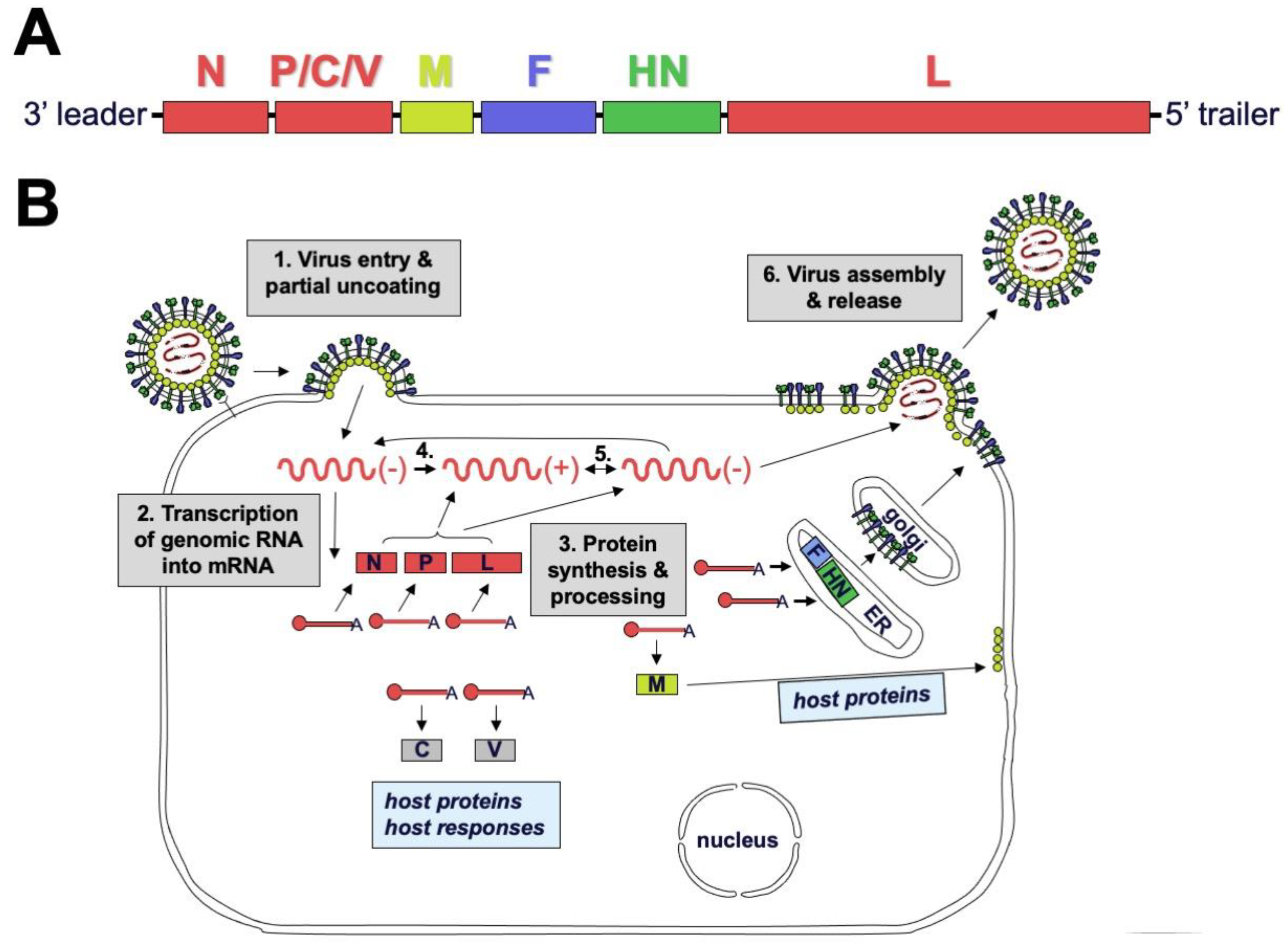
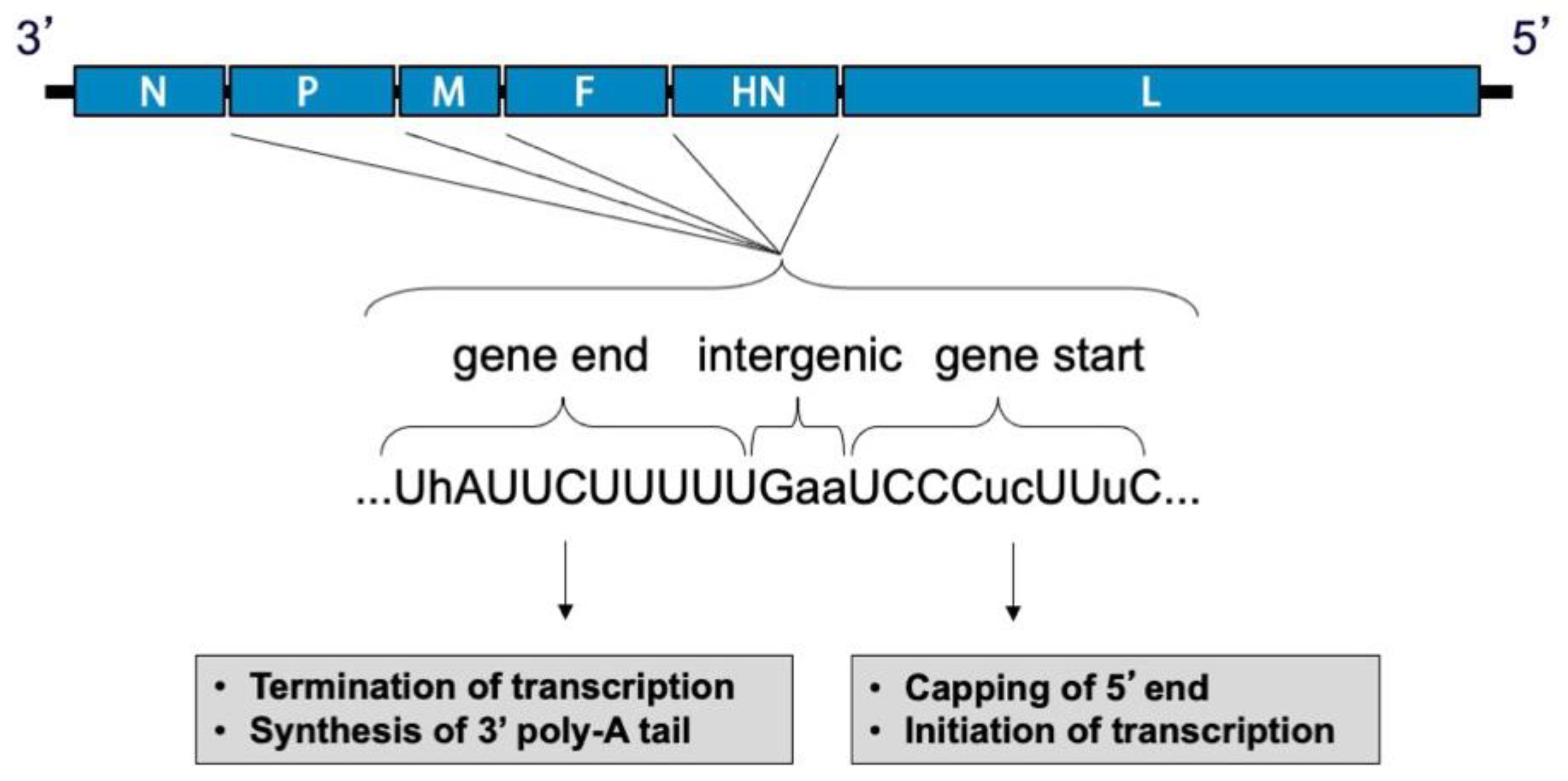
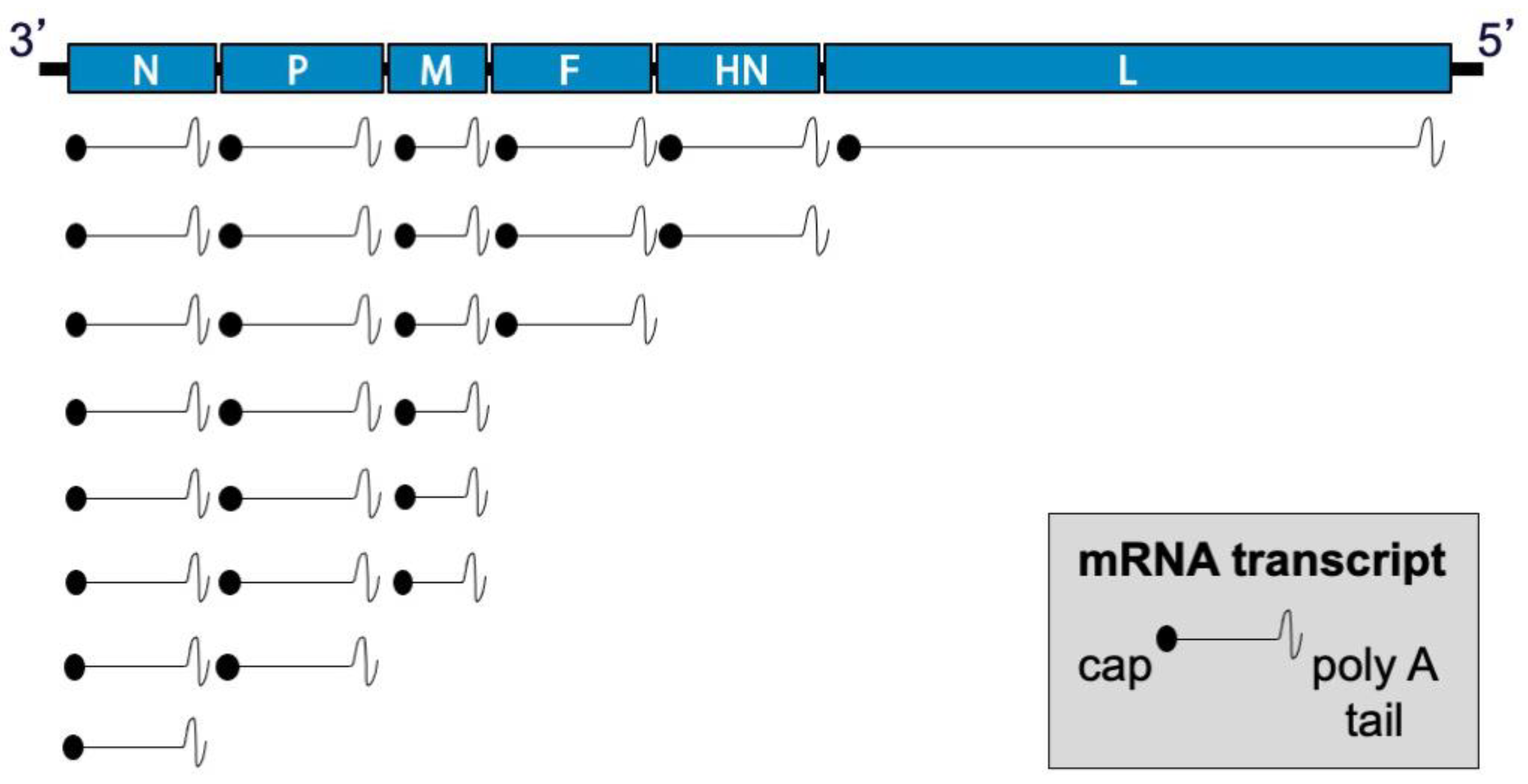
2.2. Genome and Virion Structure
2.3. Replication Cycle
2.4. Fusion (F) Glycoprotein Structures and Structural Intermediates
2.5. Attachment Protein Structures
3. Past and Current Vaccine Candidates for HRSV, HMPV, and the HPIVs
4. Sendai Virus
5. Sendai Virus as a Vaccine Platform
5.1. Sendai Virus as a Jennerian Vaccine against HPIV1
5.2. Sendai Virus-Vectored HRSV Vaccine
5.3. Sendai Virus-Vectored HPIV3 Vaccine
5.4. Sendai Virus-Vectored HPIV2 Vaccine and SeV-Vectored Vaccine Cocktails
5.5. Sendai Virus-Vectored HMPV Vaccine
6. Immunocompetence and the Vaccinated Host
7. Outlook
Funding
Conflicts of Interest
References
- Forum of international respiratory societies. In The Global Impact of Respiratory Disease, 2nd ed.; European Respiratory Society: Sheffield, UK, 2017; Available online: https://www.who.int (accessed on 3 March 2021).
- Johns Hopkins University and Medicine Coronavirus Resource Center. Available online: https://coronavirus.jhu.edu (accessed on 6 March 2021).
- Influenza 1918 Pandemic. Available online: https://www.cdc.gov (accessed on 1 May 2021).
- Nair, H.; Brooks, W.A.; Katz, M.; Roca, A.; Berkley, J.A.; Madhi, S.A.; Simmerman, J.M.; Gordon, A.; Sato, M.; Howie, S.; et al. Global burden of respiratory infections due to seasonal influenza in young children: A systematic review and meta-analysis. Lancet 2011, 378, 1917–1930. [Google Scholar] [CrossRef] [Green Version]
- Iuliano, A.D.; Roguski, K.M.; Chang, H.H.; Muscatello, D.J.; Palekar, R.; Tempia, S.; Cohen, C.; Gran, J.M.; Schanzer, D.; Cowling, B.J.; et al. Global Seasonal Influenza-associated Mortality Collaborator, N., Estimates of global seasonal influenza-associated respiratory mortality: A modelling study. Lancet 2018, 391, 1285–1300. [Google Scholar] [CrossRef]
- Gaunt, E.R.; Harvala, H.; McIntyre, C.; Templeton, K.E.; Simmonds, P. Disease burden of the most commonly detected respiratory viruses in hospitalized patients calculated using the disability adjusted life year (DALY) model. J. Clin. Virol. 2011, 52, 215–221. [Google Scholar] [CrossRef]
- Edwards, K.M.; Zhu, Y.; Griffin, M.R.; Weinberg, G.A.; Hall, C.B.; Szilagyi, P.G.; Staat, M.A.; Iwane, M.; Prill, M.M.; Williams, J.V.; et al. Burden of human metapneumovirus infection in young children. N. Engl. J. Med. 2013, 368, 633–643. [Google Scholar] [CrossRef] [Green Version]
- Shi, T.; McAllister, D.A.; O’Brien, K.L.; Simoes, E.A.F.; Madhi, S.A.; Gessner, B.D.; Polack, F.P.; Balsells, E.; Acacio, S.; Aguayo, C.; et al. Global, regional, and national disease burden estimates of acute lower respiratory infections due to respiratory syncy-tial virus in young children in 2015: A systematic review and modelling study. Lancet 2017, 390, 946–958. [Google Scholar] [CrossRef] [Green Version]
- Respiratory Syncytial Virus (RSV). Available online: https://www.niaid.nih.gov (accessed on 3 March 2021).
- Nair, H.; Nokes, D.J.; Gessner, B.D.; Dherani, M.; Madhi, S.A.; Singleton, R.J.; O’Brien, K.L.; Roca, A.; Wright, P.F.; Bruce, N.; et al. Global burden of acute lower respiratory infections due to respiratory syncytial virus in young children: A systematic review and meta-analysis. Lancet 2010, 375, 1545–1555. [Google Scholar] [CrossRef] [Green Version]
- Scheltema, N.M.; Gentile, A.; Lucion, F.; Nokes, D.J.; Munywoki, P.K.; Madhi, S.A.; Groome, M.J.; Cohen, C.; Moyes, J.; Thorburn, K.; et al. Global respiratory syncytial virus-associated mortality in young children (RSV GOLD): A retrospective case series. Lancet Glob. Health 2017, 5, e984–e991. [Google Scholar] [CrossRef] [Green Version]
- Lozano, R.; Naghavi, M.; Foreman, K.; Lim, S.; Shibuya, K.; Aboyans, V.; Abraham, J.; Adair, T.; Aggarwal, R.; Ahn, S.Y.; et al. Global and regional mortality from 235 causes of death for 20 age groups in 1990 and 2010: A systematic analysis for the Global Burden of Disease Study 2010. Lancet 2012, 380, 2095–2128. [Google Scholar] [CrossRef]
- Byington, C.L.; Wilkes, J.; Korgenski, K.; Sheng, X. Respiratory syncytial virus-associated mortality in hospitalized infants and young children. Pediatrics 2015, 135, e24–e31. [Google Scholar] [CrossRef] [Green Version]
- Branche, A.R.; Falsey, A.R. Parainfluenza Virus Infection. Semin. Respir. Crit Care Med. 2016, 37, 538–554. [Google Scholar] [CrossRef] [PubMed]
- Williams, J.V.; Edwards, K.M.; Weinberg, G.A.; Griffin, M.R.; Hall, C.B.; Zhu, Y.; Szilagyi, P.G.; Wang, C.K.; Yang, C.F.; Silva, D.; et al. Population-based incidence of human metapneumovirus infection among hospitalized children. J. Infect. Dis. 2010, 201, 1890–1898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Griffiths, C.; Drews, S.J.; Marchant, D.J. Respiratory Syncytial Virus: Infection, Detection, and New Options for Prevention and Treatment. Clin. Microbiol. Rev. 2017, 30, 277–319. [Google Scholar] [CrossRef] [Green Version]
- Groothuis, J.R.; Simoes, E.A.; Hemming, V.G. Respiratory syncytial virus (RSV) infection in preterm infants and the protective effects of RSV immune globulin (RSVIG). Respiratory Syncytial Virus Immune Globulin Study Group. Pediatrics 1995, 95, 463–467. [Google Scholar]
- Aliprantis, A.O.; Wolford, D.; Caro, L.; Maas, B.M.; Ma, H.; Montgomery, D.L.; Sterling, L.M.; Hunt, A.; Cox, K.S.; Vora, K.A.; et al. A Phase 1 Randomized, Double-Blind, Placebo-Controlled Trial to Assess the Safety, Tolerability, and Pharmacokinetics of a Respiratory Syncytial Virus Neutralizing Monoclonal Antibody MK-1654 in Healthy Adults. Clin. Pharm. Drug Dev. 2020, 10, 556–566. [Google Scholar] [CrossRef] [PubMed]
- Anderson, E.J.; Carosone-Link, P.; Yogev, R.; Yi, J.; Simoes, E.A.F. Effectiveness of Palivizumab in High-risk Infants and Children: A Propensity Score Weighted Regression Analysis. Pediatr. Infect. Dis. J. 2017, 36, 699–704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Griffin, M.P.; Yuan, Y.; Takas, T.; Domachowske, J.B.; Madhi, S.A.; Manzoni, P.; Simoes, E.A.F.; Esser, M.T.; Khan, A.A.; Dubovsky, F.; et al. Nirsevimab Study, G., Single-Dose Nirsevimab for Prevention of RSV in Preterm Infants. N. Engl. J. Med. 2020, 383, 415–425. [Google Scholar] [CrossRef]
- Afonso, C.L.; Amarasinghe, G.K.; Banyai, K.; Bao, Y.; Basler, C.F.; Bavari, S.; Bejerman, N.; Blasdell, K.R.; Briand, F.X.; Briese, T.; et al. Taxonomy of the order Mononegavirales: Update 2016. Arch Virol. 2016, 161, 2351–2360. [Google Scholar] [CrossRef]
- Amarasinghe, G.K.; Bao, Y.; Basler, C.F.; Bavari, S.; Beer, M.; Bejerman, N.; Blasdell, K.R.; Bochnowski, A.; Briese, T.; Bukreyev, A.; et al. Taxonomy of the order Mononegavirales: Update 2017. Arch Virol. 2017, 162, 2493–2504. [Google Scholar] [CrossRef] [PubMed]
- Phan, S.I.; Chen, Z.; Xu, P.; Li, Z.; Gao, X.; Foster, S.L.; Teng, M.N.; Tripp, R.A.; Sakamoto, K.; He, B. A respiratory syncytial virus (RSV) vaccine based on parainfluenza virus 5 (PIV5). Vaccine 2014, 32, 3050–3057. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Phan, S.; DiStefano, D.J.; Citron, M.P.; Callahan, C.L.; Indrawati, L.; Dubey, S.A.; Heidecker, G.J.; Govindarajan, D.; Liang, X.; et al. A Single-Dose Recombinant Parainfluenza Virus 5-Vectored Vaccine Expressing Respiratory Syncytial Virus (RSV) F or G Protein Protected Cotton Rats and African Green Monkeys from RSV Challenge. J. Virol. 2017, 91, e00066-17. [Google Scholar] [CrossRef] [Green Version]
- Lamb, R.A.; Parks, G.D. Paramyxoviridae: The viruses and their replication. In Fields Virology, 5th ed.; Knipe, D.M., Howley, P.M., Griffin, D.E., Martin, M.A., Lamb, R.A., Roizman, B., Straus, S.E., Eds.; Lippincott Williams and Wilkins: Philadelphia, PA, USA, 2007; Volume 1, pp. 1449–1496. [Google Scholar]
- Jeffree, C.E.; Rixon, H.W.; Brown, G.; Aitken, J.; Sugrue, R.J. Distribution of the attachment (G) glycoprotein and GM1 within the envelope of mature respiratory syncytial virus filaments revealed using field emission scanning electron microscopy. Virology 2003, 306, 254–267. [Google Scholar] [CrossRef] [Green Version]
- Roberts, S.R.; Compans, R.W.; Wertz, G.W. Respiratory syncytial virus matures at the apical surfaces of polarized epithelial cells. J. Virol. 1995, 69, 2667–2673. [Google Scholar] [CrossRef] [Green Version]
- Choppin, P.W.; Compans, R.W. Reproduction of paramyxoviruses. In Comprehensive Virology, Fraenkel-Conrat, H.; Wagner, R.R., Ed.; Plenum Press: New York, NY, USA, 1975; Volume 4, pp. 95–117. [Google Scholar]
- Lamb, R.A.; Mahy, B.W.; Choppin, P.W. The synthesis of sendai virus polypeptides in infected cells. Virology 1976, 69, 116–131. [Google Scholar] [CrossRef]
- Russell, C.J.; Luque, L.E. The structural basis of paramyxovirus invasion. Trends Microbiol. 2006, 14, 243–246. [Google Scholar] [CrossRef] [PubMed]
- Russell, C.J.; Kantor, K.L.; Jardetzky, T.S.; Lamb, R.A. A dual-functional paramyxovirus F protein regulatory switch segment: Activation and membrane fusion. J. Cell Biol. 2003, 163, 363–374. [Google Scholar] [CrossRef]
- Russell, C.J.; Jardetzky, T.S.; Lamb, R.A. Membrane fusion machines of paramyxoviruses: Capture of intermediates of fusion. EMBO J. 2001, 20, 4024–4034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kato, A.; Kiyotani, K.; Hasan, M.K.; Shioda, T.; Sakai, Y.; Yoshida, T.; Nagai, Y. Sendai virus gene start signals are not equivalent in reinitiation capacity: Moderation at the fusion protein gene. J. Virol. 1999, 73, 9237–9246. [Google Scholar] [CrossRef] [Green Version]
- Hasan, M.K.; Kato, A.; Shioda, T.; Sakai, Y.; Yu, D.; Nagai, Y. Creation of an infectious recombinant Sendai virus expressing the firefly luciferase gene from the 3’ proximal first locus. J. Gen. Virol. 1997, 78 Pt 11, 2813–2820. [Google Scholar] [CrossRef]
- Tokusumi, T.; Iida, A.; Hirata, T.; Kato, A.; Nagai, Y.; Hasegawa, M. Recombinant Sendai viruses expressing different levels of a foreign reporter gene. Virus Res. 2002, 86, 33–38. [Google Scholar] [CrossRef]
- Schmitt, A.P.; Leser, G.P.; Morita, E.; Sundquist, W.I.; Lamb, R.A. Evidence for a new viral late-domain core sequence, FPIV, necessary for budding of a paramyxovirus. J. Virol. 2005, 79, 2988–2997. [Google Scholar] [CrossRef] [Green Version]
- Schmitt, A.P.; Leser, G.P.; Waning, D.L.; Lamb, R.A. Requirements for budding of paramyxovirus simian virus 5 virus-like particles. J. Virol. 2002, 76, 3952–3964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takimoto, T.; Murti, K.G.; Bousse, T.; Scroggs, R.A.; Portner, A. Role of matrix and fusion proteins in budding of Sendai virus. J. Virol. 2001, 75, 11384–11391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fouillot-Coriou, N.; Roux, L. Structure-function analysis of the Sendai virus F and HN cytoplasmic domain: Different role for the two proteins in the production of virus particle. Virology 2000, 270, 464–475. [Google Scholar] [CrossRef] [PubMed]
- Lamb, R.A.; Jardetzky, T.S. Structural basis of viral invasion: Lessons from paramyxovirus F. Curr. Opin. Struct. Biol. 2007, 17, 427–436. [Google Scholar] [CrossRef] [Green Version]
- Yin, H.S.; Wen, X.; Paterson, R.G.; Lamb, R.A.; Jardetzky, T.S. Structure of the parainfluenza virus 5 F protein in its metastable, prefusion conformation. Nature 2006, 439, 38–44. [Google Scholar] [CrossRef]
- McLellan, J.S.; Chen, M.; Leung, S.; Graepel, K.W.; Du, X.; Yang, Y.; Zhou, T.; Baxa, U.; Yasuda, E.; Beaumont, T.; et al. Structure of RSV fusion glycoprotein trimer bound to a prefusion-specific neutralizing antibody. Science 2013, 340, 1113–1117. [Google Scholar] [CrossRef] [Green Version]
- Wen, X.; Krause, J.C.; Leser, G.P.; Cox, R.G.; Lamb, R.A.; Williams, J.V.; Crowe, J.E., Jr.; Jardetzky, T.S. Structure of the human metapneumovirus fusion protein with neutralizing antibody identifies a pneumovirus antigenic site. Nat. Struct. Mol. Biol. 2012, 19, 461–463. [Google Scholar] [CrossRef] [PubMed]
- Battles, M.B.; Mas, V.; Olmedillas, E.; Cano, O.; Vazquez, M.; Rodriguez, L.; Melero, J.A.; McLellan, J.S. Structure and immunogenicity of pre-fusion-stabilized human metapneumovirus F glycoprotein. Nat. Commun. 2017, 8, 1528. [Google Scholar] [CrossRef]
- Wong, J.J.; Paterson, R.G.; Lamb, R.A.; Jardetzky, T.S. Structure and stabilization of the Hendra virus F glycoprotein in its prefusion form. Proc. Natl. Acad. Sci. USA 2016, 113, 1056–1061. [Google Scholar] [CrossRef] [Green Version]
- Yin, H.S.; Paterson, R.G.; Wen, X.; Lamb, R.A.; Jardetzky, T.S. Structure of the uncleaved ectodomain of the paramyxovirus (hPIV3) fusion protein. Proc. Natl. Acad. Sci. USA 2005, 102, 9288–9293. [Google Scholar] [CrossRef] [Green Version]
- Paterson, R.G.; Russell, C.J.; Lamb, R.A. Fusion protein of the paramyxovirus SV5: Destabilizing and stabilizing mutants of fusion activation. Virology 2000, 270, 17–30. [Google Scholar] [CrossRef] [Green Version]
- Bose, S.; Jardetzky, T.S.; Lamb, R.A. Timing is everything: Fine-tuned molecular machines orchestrate paramyxovirus entry. Virology 2015, 479, 518–531. [Google Scholar] [CrossRef] [Green Version]
- Baker, K.A.; Dutch, R.E.; Lamb, R.A.; Jardetzky, T.S. Structural basis for paramyxovirus-mediated membrane fusion. Mol. Cell 1999, 3, 309–319. [Google Scholar] [CrossRef]
- Russell, C.J.; Jardetzky, T.S.; Lamb, R.A. Conserved glycine residues in the fusion peptide of the paramyxovirus fusion protein regulate activation of the native state. J. Virol. 2004, 78, 13727–13742. [Google Scholar] [CrossRef] [Green Version]
- Swanson, K.; Wen, X.; Leser, G.P.; Paterson, R.G.; Lamb, R.A.; Jardetzky, T.S. Structure of the Newcastle disease virus F protein in the post-fusion conformation. Virology 2010, 402, 372–379. [Google Scholar] [CrossRef]
- Chen, L.; Gorman, J.J.; McKimm-Breschkin, J.; Lawrence, L.J.; Tulloch, P.A.; Smith, B.J.; Colman, P.M.; Lawrence, M.C. The structure of the fusion glycoprotein of Newcastle disease virus suggests a novel paradigm for the molecular mechanism of membrane fusion. Structure 2001, 9, 255–266. [Google Scholar] [CrossRef] [Green Version]
- McLellan, J.S.; Yang, Y.; Graham, B.S.; Kwong, P.D. Structure of respiratory syncytial virus fusion glycoprotein in the postfusion conformation reveals preservation of neutralizing epitopes. J. Virol. 2011, 85, 7788–7796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swanson, K.A.; Settembre, E.C.; Shaw, C.A.; Dey, A.K.; Rappuoli, R.; Mandl, C.W.; Dormitzer, P.R.; Carfi, A. Structural basis for immunization with postfusion respiratory syncytial virus fusion F glycoprotein (RSV F) to elicit high neutralizing antibody titers. Proc. Natl. Acad. Sci. USA 2011, 108, 9619–9624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crennell, S.; Takimoto, T.; Portner, A.; Taylor, G. Crystal structure of the multifunctional paramyxovirus hemagglutinin-neuraminidase. Nat. Struct. Biol. 2000, 7, 1068–1074. [Google Scholar] [PubMed]
- Yuan, P.; Paterson, R.G.; Leser, G.P.; Lamb, R.A.; Jardetzky, T.S. Structure of the ulster strain newcastle disease virus hemagglutinin-neuraminidase reveals auto-inhibitory interactions associated with low virulence. PLoS Pathog. 2012, 8, e1002855. [Google Scholar] [CrossRef]
- Yuan, P.; Swanson, K.A.; Leser, G.P.; Paterson, R.G.; Lamb, R.A.; Jardetzky, T.S. Structure of the Newcastle disease virus hemagglutinin-neuraminidase (HN) ectodomain reveals a four-helix bundle stalk. Proc. Natl. Acad. Sci. USA 2011, 108, 14920–14925. [Google Scholar] [CrossRef] [Green Version]
- Lawrence, M.C.; Borg, N.A.; Streltsov, V.A.; Pilling, P.A.; Epa, V.C.; Varghese, J.N.; McKimm-Breschkin, J.L.; Colman, P.M. Structure of the haemagglutinin-neuraminidase from human parainfluenza virus type III. J. Mol. Biol. 2004, 335, 1343–1357. [Google Scholar] [CrossRef] [PubMed]
- Colf, L.A.; Juo, Z.S.; Garcia, K.C. Structure of the measles virus hemagglutinin. Nat. Struct. Mol. Biol. 2007, 14, 1227–1228. [Google Scholar] [CrossRef] [PubMed]
- Hashiguchi, T.; Kajikawa, M.; Maita, N.; Takeda, M.; Kuroki, K.; Sasaki, K.; Kohda, D.; Yanagi, Y.; Maenaka, K. Crystal structure of measles virus hemagglutinin provides insight into effective vaccines. Proc. Natl. Acad. Sci. USA 2007, 104, 19535–19540. [Google Scholar] [CrossRef] [Green Version]
- Hashiguchi, T.; Ose, T.; Kubota, M.; Maita, N.; Kamishikiryo, J.; Maenaka, K.; Yanagi, Y. Structure of the measles virus hemagglutinin bound to its cellular receptor SLAM. Nat. Struct. Mol. Biol. 2011, 18, 135–141. [Google Scholar] [CrossRef]
- Yuan, P.; Thompson, T.B.; Wurzburg, B.A.; Paterson, R.G.; Lamb, R.A.; Jardetzky, T.S. Structural studies of the parainfluenza virus 5 hemagglutinin-neuraminidase tetramer in complex with its receptor, sialyllactose. Structure 2005, 13, 803–815. [Google Scholar] [CrossRef] [Green Version]
- Xu, K.; Rajashankar, K.R.; Chan, Y.P.; Himanen, J.P.; Broder, C.C.; Nikolov, D.B. Host cell recognition by the henipaviruses: Crystal structures of the Nipah G attachment glycoprotein and its complex with ephrin-B3. Proc. Natl. Acad. Sci. USA 2008, 105, 9953–9958. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bowden, T.A.; Aricescu, A.R.; Gilbert, R.J.; Grimes, J.M.; Jones, E.Y.; Stuart, D.I. Structural basis of Nipah and Hendra virus attachment to their cell-surface receptor ephrin-B2. Nat. Struct. Mol. Biol. 2008, 15, 567–572. [Google Scholar] [CrossRef]
- Bowden, T.A.; Crispin, M.; Harvey, D.J.; Jones, E.Y.; Stuart, D.I. Dimeric architecture of the Hendra virus attachment glycoprotein: Evidence for a conserved mode of assembly. J. Virol. 2010, 84, 6208–6217. [Google Scholar] [CrossRef] [Green Version]
- Fedechkin, S.O.; George, N.L.; Wolff, J.T.; Kauvar, L.M.; DuBois, R.M. Structures of respiratory syncytial virus G antigen bound to broadly neutralizing antibodies. Sci. Immunol. 2018, 3. [Google Scholar] [CrossRef] [Green Version]
- Fedechkin, S.O.; George, N.L.; Nunez Castrejon, A.M.; Dillen, J.R.; Kauvar, L.M.; DuBois, R.M. Conformational Flexibility in Respiratory Syncytial Virus G Neutralizing Epitopes. J. Virol. 2020, 94, e01879-19. [Google Scholar] [CrossRef] [Green Version]
- Murphy, B.R.; Walsh, E.E. Formalin-inactivated respiratory syncytial virus vaccine induces antibodies to the fusion glycoprotein that are deficient in fusion-inhibiting activity. J. Clin. Microbiol. 1988, 26, 1595–1597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murphy, B.R.; Prince, G.A.; Walsh, E.E.; Kim, H.W.; Parrott, R.H.; Hemming, V.G.; Rodriguez, W.J.; Chanock, R.M. Dissociation between serum neutralizing and glycoprotein antibody responses of infants and children who received inactivated respiratory syncytial virus vaccine. J. Clin. Microbiol. 1986, 24, 197–202. [Google Scholar] [CrossRef] [Green Version]
- Fulginiti, V.A.; Eller, J.J.; Sieber, O.F.; Joyner, J.W.; Minamitani, M.; Meiklejohn, G. Respiratory virus immunization I. A field trial of two inactivated respiratory virus vaccines; an aqueous trivalent parainfluenza virus vaccine and an alum-precipitated respiratory syncytial virus vaccine. Am. J. Epidemiol. 1969, 89, 435–448. [Google Scholar] [CrossRef]
- Biagi, C.; Dondi, A.; Scarpini, S.; Rocca, A.; Vandini, S.; Poletti, G.; Lanari, M. Current State and Challenges in Developing Respiratory Syncytial Virus Vaccines. Vaccines 2020, 8, 672. [Google Scholar] [CrossRef] [PubMed]
- Killikelly, A.; Tunis, M.; House, A.; Quach, C.; Vaudry, W.; Moore, D. Overview of the respiratory syncytial virus vaccine candidate pipeline in Canada. Can. Commun. Dis. Rep. 2020, 46, 56–61. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.I.; DeVincenzo, J.P.; Jones, B.G.; Rudraraju, R.; Harrison, L.; Meyers, R.; Cehelsky, J.; Alvarez, R.; Hurwitz, J.L. Respiratory syncytial virus human experimental infection model: Provenance, production, and sequence of low-passaged memphis-37 challenge virus. PLoS ONE 2014, 9, e113100. [Google Scholar] [CrossRef] [Green Version]
- RSV Clinical Trial Tracker. Available online: https://www.path.org/resources/rsv-and-mab-trial-tracker (accessed on 1 May 2021).
- Schickli, J.H.; Kaur, J.; Tang, R.S. Nonclinical phenotypic and genotypic analyses of a Phase 1 pediatric respiratory syncytial virus vaccine candidate MEDI-559 (rA2cp248/404/1030DeltaSH) at permissive and non-permissive temperatures. Virus Res. 2012, 169, 38–47. [Google Scholar] [CrossRef] [PubMed]
- Mok, H.; Tollefson, S.J.; Podsiad, A.B.; Shepherd, B.E.; Polosukhin, V.V.; Johnston, R.E.; Williams, J.V.; Crowe, J.E., Jr. An alphavirus replicon-based human metapneumovirus vaccine is immunogenic and protective in mice and cotton rats. J. Virol. 2008, 82, 11410–11418. [Google Scholar] [CrossRef] [Green Version]
- Skiadopoulos, M.H.; Schmidt, A.C.; Riggs, J.M.; Surman, S.R.; Elkins, W.R.; St Claire, M.; Collins, P.L.; Murphy, B.R. Determinants of the host range restriction of replication of bovine parainfluenza virus type 3 in rhesus monkeys are polygenic. J. Virol. 2003, 77, 1141–1148. [Google Scholar] [CrossRef] [Green Version]
- Karron, R.A.; Thumar, B.; Schappell, E.; Surman, S.; Murphy, B.R.; Collins, P.L.; Schmidt, A.C. Evaluation of two chimeric bovine-human parainfluenza virus type 3 vaccines in infants and young children. Vaccine 2012, 30, 3975–3981. [Google Scholar] [CrossRef]
- Crank, M.C.; Ruckwardt, T.J.; Chen, M.; Morabito, K.M.; Phung, E.; Costner, P.J.; Holman, L.A.; Hickman, S.P.; Berkowitz, N.M.; Gordon, I.J.; et al. A proof of concept for structure-based vaccine design targeting RSV in humans. Science 2019, 365, 505–509. [Google Scholar] [CrossRef]
- Kim, K.H.; Lee, Y.T.; Hwang, H.S.; Kwon, Y.M.; Kim, M.C.; Ko, E.J.; Lee, J.S.; Lee, Y.; Kang, S.M. Virus-Like Particle Vaccine Containing the F Protein of Respiratory Syncytial Virus Confers Protection without Pulmonary Disease by Modulating Specific Subsets of Dendritic Cells and Effector T Cells. J. Virol. 2015, 89, 11692–11705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madhi, S.A.; Polack, F.P.; Piedra, P.A.; Munoz, F.M.; Trenholme, A.A.; Simoes, E.A.F.; Swamy, G.K.; Agrawal, S.; Ahmed, K.; August, A.; et al. Respiratory Syncytial Virus Vaccination during Pregnancy and Effects in Infants. N. Engl. J. Med. 2020, 383, 426–439. [Google Scholar] [CrossRef]
- Polack, F.P.; Thomas, S.J.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Perez, J.L.; Perez Marc, G.; Moreira, E.D.; Zerbini, C.; et al. Safety and Efficacy of the BNT162b2 mRNA Covid-19 Vaccine. N. Engl. J. Med. 2020, 383, 2603–2615. [Google Scholar] [CrossRef]
- Widge, A.T.; Rouphael, N.G.; Jackson, L.A.; Anderson, E.J.; Roberts, P.C.; Makhene, M.; Chappell, J.D.; Denison, M.R.; Stevens, L.J.; Pruijssers, A.J.; et al. Durability of Responses after SARS-CoV-2 mRNA-1273 Vaccination. N. Engl. J. Med. 2021, 384, 80–82. [Google Scholar] [CrossRef] [PubMed]
- Kuroya, M.; Ishida, N. Newborn virus pneumonitis (type Sendai). II. The isolation of a new virus possessing hemagglutinin activity. Yokohama Med. Bull. 1953, 4, 217–233. [Google Scholar] [PubMed]
- Fukumi, H.; Nishikawa, F.; Kitayama, T. A pneumotropic virus from mice causing hemagglutination. Jpn. J. Med. Sci. Biol. 1954, 7, 345–363. [Google Scholar] [CrossRef] [Green Version]
- Ishida, N.; Homma, M. Sendai virus. Adv. Virus Res. 1978, 23, 349–383. [Google Scholar] [PubMed]
- Karron, R.A.; Collins, P.L. Parainfluenza viruses. In Fields Virology, 5th ed.; Knipe, D.M., Howley, P.M., Griffin, D.E., Martin, M.A., Lamb, R.A., Roizman, B., Straus, S.E., Eds.; Lippincott Williams and Wilkins: Philadelphia, PA, USA, 2007; pp. 1497–1526. [Google Scholar]
- Bhatt, P.N.; Jonas, A.M. An epizootic of Sendai infection with mortality in a barrier-maintained mouse colony. Am. J. Epidemiol. 1974, 100, 222–229. [Google Scholar] [CrossRef] [PubMed]
- Parker, J.C.; Tennant, R.W.; Ward, T.G.; Rowe, W.P. Enzootic Sendai Virus Infections in Mouse Breeder Colonies within the United States. Science 1964, 146, 936–938. [Google Scholar] [CrossRef] [PubMed]
- Profeta, M.L.; Lief, F.S.; Plotkin, S.A. Enzootic sendai infection in laboratory hamsters. Am. J. Epidemiol. 1969, 89, 316–324. [Google Scholar] [CrossRef]
- Faisca, P.; Anh, D.B.; Desmecht, D.J. Sendai virus-induced alterations in lung structure/function correlate with viral loads and reveal a wide resistance/susceptibility spectrum among mouse strains. Am. J. Physiol. Lung Cell Mol. Physiol. 2005, 289, L777–L787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burke, C.W.; Bridges, O.; Brown, S.; Rahija, R.; Russell, C.J. Mode of parainfluenza virus transmission determines the dynamics of primary infection and protection from reinfection. PLoS Pathog. 2013, 9, e1003786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burke, C.W.; Li, M.; Hurwitz, J.L.; Vogel, P.; Russell, C.J. Relationships among dissemination of primary parainfluenza virus infection in the respiratory tract, mucosal and peripheral immune responses, and protection from reinfection: A noninvasive bioluminescence-imaging study. J. Virol. 2015, 89, 3568–3583. [Google Scholar] [CrossRef] [Green Version]
- Burke, C.W.; Mason, J.N.; Surman, S.L.; Jones, B.G.; Dalloneau, E.; Hurwitz, J.L.; Russell, C.J. Illumination of parainfluenza virus infection and transmission in living animals reveals a tissue-specific dichotomy. PLoS Pathog. 2011, 7, e1002134. [Google Scholar] [CrossRef] [Green Version]
- Crotty, S.; Felgner, P.; Davies, H.; Glidewell, J.; Villarreal, L.; Ahmed, R. Cutting edge: Long-term B cell memory in humans after smallpox vaccination. J. Immunol. 2003, 171, 4969–4973. [Google Scholar] [CrossRef] [Green Version]
- Amanna, I.J.; Slifka, M.K.; Crotty, S. Immunity and immunological memory following smallpox vaccination. Immunol. Rev. 2006, 211, 320–337. [Google Scholar] [CrossRef] [PubMed]
- Gorman, W.L.; Gill, D.S.; Scroggs, R.A.; Portner, A. The hemagglutinin-neuraminidase glycoproteins of human parainfluenza virus type 1 and Sendai virus have high structure-function similarity with limited antigenic cross-reactivity. Virology 1990, 175, 211–221. [Google Scholar] [CrossRef]
- Lyn, D.; Gill, D.S.; Scroggs, R.A.; Portner, A. The nucleoproteins of human parainfluenza virus type 1 and Sendai virus share amino acid sequences and antigenic and structural determinants. J. Gen. Virol. 1991, 72, 983–987. [Google Scholar] [CrossRef]
- Sealy, R.; Jones, B.G.; Surman, S.L.; Hurwitz, J.L. Robust IgA and IgG-producing antibody forming cells in the diffuse-NALT and lungs of Sendai virus-vaccinated cotton rats associate with rapid protection against human parainfluenza virus-type 1. Vaccine 2010, 28, 6749–6756. [Google Scholar] [CrossRef] [Green Version]
- Hurwitz, J.L.; Soike, K.F.; Sangster, M.Y.; Portner, A.; Sealy, R.E.; Dawson, D.H.; Coleclough, C. Intranasal Sendai virus vaccine protects African green monkeys from infection with human parainfluenza virus-type one. Vaccine 1997, 15, 533–540. [Google Scholar] [CrossRef]
- Skiadopoulos, M.H.; Surman, S.R.; Riggs, J.M.; Elkins, W.R.; St Claire, M.; Nishio, M.; Garcin, D.; Kolakofsky, D.; Collins, P.L.; Murphy, B.R. Sendai virus, a murine parainfluenza virus type 1, replicates to a level similar to human PIV1 in the upper and lower respiratory tract of African green monkeys and chimpanzees. Virology 2002, 297, 153–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slobod, K.S.; Shenep, J.L.; Lujan-Zilbermann, J.; Allison, K.; Brown, B.; Scroggs, R.A.; Portner, A.; Coleclough, C.; Hurwitz, J.L. Safety and immunogenicity of intranasal murine parainfluenza virus type 1 (Sendai virus) in healthy human adults. Vaccine 2004, 22, 3182–3186. [Google Scholar] [CrossRef] [PubMed]
- Adderson, E.; Branum, K.; Sealy, R.E.; Jones, B.G.; Surman, S.L.; Penkert, R.; Freiden, P.; Slobod, K.S.; Gaur, A.H.; Hayden, R.T.; et al. Safety and immunogenicity of an intranasal Sendai virus-based human parainfluenza virus type 1 vaccine in 3- to 6-year-old children. Clin. Vaccine Immunol. 2015, 22, 298–303. [Google Scholar] [CrossRef]
- Takimoto, T.; Hurwitz, J.L.; Coleclough, C.; Prouser, C.; Krishnamurthy, S.; Zhan, X.; Boyd, K.; Scroggs, R.A.; Brown, B.; Nagai, Y.; et al. Recombinant Sendai virus expressing the G glycoprotein of respiratory syncytial virus (RSV) elicits immune protection against RSV. J. Virol. 2004, 78, 6043–6047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhan, X.; Hurwitz, J.L.; Krishnamurthy, S.; Takimoto, T.; Boyd, K.; Scroggs, R.A.; Surman, S.; Portner, A.; Slobod, K.S. Respiratory syncytial virus (RSV) fusion protein expressed by recombinant Sendai virus elicits B-cell and T-cell responses in cotton rats and confers protection against RSV subtypes A and B. Vaccine 2007, 25, 8782–8793. [Google Scholar] [CrossRef] [Green Version]
- Jones, B.G.; Sealy, R.E.; Surman, S.L.; Portner, A.; Russell, C.J.; Slobod, K.S.; Dormitzer, P.R.; DeVincenzo, J.; Hurwitz, J.L. Sendai virus-based RSV vaccine protects against RSV challenge in an in vivo maternal antibody model. Vaccine 2014, 32, 3264–3273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhan, X.; Slobod, K.S.; Krishnamurthy, S.; Luque, L.E.; Takimoto, T.; Jones, B.; Surman, S.; Russell, C.J.; Portner, A.; Hurwitz, J.L. Sendai virus recombinant vaccine expressing hPIV-3 HN or F elicits protective immunity and combines with a second recombinant to prevent hPIV-1, hPIV-3 and RSV infections. Vaccine 2008, 26, 3480–3488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, B.; Zhan, X.; Mishin, V.; Slobod, K.S.; Surman, S.; Russell, C.J.; Portner, A.; Hurwitz, J.L. Human PIV-2 recombinant Sendai virus (rSeV) elicits durable immunity and combines with two additional rSeVs to protect against hPIV-1, hPIV-2, hPIV-3, and RSV. Vaccine 2009, 27, 1848–1857. [Google Scholar] [CrossRef] [Green Version]
- Jones, B.G.; Sealy, R.E.; Rudraraju, R.; Traina-Dorge, V.L.; Finneyfrock, B.; Cook, A.; Takimoto, T.; Portner, A.; Hurwitz, J.L. Sendai virus-based RSV vaccine protects African green monkeys from RSV infection. Vaccine 2012, 30, 959–968. [Google Scholar] [CrossRef] [Green Version]
- Zhan, X.; Slobod, K.S.; Jones, B.G.; Sealy, R.E.; Takimoto, T.; Boyd, K.; Surman, S.; Russell, C.J.; Portner, A.; Hurwitz, J.L. Sendai virus recombinant vaccine expressing a secreted, unconstrained respiratory syncytial virus fusion protein protects against RSV in cotton rats. Int. Immunol. 2015, 27, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Mason, J.N.; Elbahesh, H.; Russell, C.J. Influence of antigen insertion site and vector dose on immunogenicity and protective capacity in Sendai virus-based human parainfluenza virus type 3 vaccines. J. Virol. 2013, 87, 5959–5969. [Google Scholar] [CrossRef] [Green Version]
- Russell, C.J.; Jones, B.G.; Sealy, R.E.; Surman, S.L.; Mason, J.N.; Hayden, R.T.; Tripp, R.A.; Takimoto, T.; Hurwitz, J.L. A Sendai virus recombinant vaccine expressing a gene for truncated human metapneumovirus (hMPV) fusion protein protects cotton rats from hMPV challenge. Virology 2017, 509, 60–66. [Google Scholar] [CrossRef]
- Dave, V.P.; Allan, J.E.; Slobod, K.S.; Smith, F.S.; Ryan, K.W.; Takimoto, T.; Power, U.F.; Portner, A.; Hurwitz, J.L. Viral cross-reactivity and antigenic determinants recognized by human parainfluenza virus type 1-specific cytotoxic T-cells. Virology 1994, 199, 376–383. [Google Scholar] [CrossRef]
- Smith, F.S.; Portner, A.; Leggiadro, R.J.; Turner, E.V.; Hurwitz, J.L. Age-related development of human memory T-helper and B-cell responses toward parainfluenza virus type-1. Virology 1994, 205, 453–461. [Google Scholar] [CrossRef]
- Sangster, M.; Smith, F.S.; Coleclough, C.; Hurwitz, J.L. Human parainfluenza virus type 1 immunization of infant mice protects from subsequent Sendai virus infection. Virology 1995, 212, 13–19. [Google Scholar] [CrossRef] [Green Version]
- Rudraraju, R.; Surman, S.; Jones, B.; Sealy, R.; Woodland, D.L.; Hurwitz, J.L. Phenotypes and functions of persistent Sendai virus-induced antibody forming cells and CD8+ T cells in diffuse nasal-associated lymphoid tissue typify lymphocyte responses of the gut. Virology 2011, 410, 429–436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sealy, R.E.; Surman, S.L.; Vogel, P.; Hurwitz, J.L. Antibody-secreting cells in respiratory tract tissues in the absence of eosinophils as supportive partners. Int. Immunol. 2016, 28, 559–564. [Google Scholar] [CrossRef] [Green Version]
- Sealy, R.; Webby, R.J.; Crumpton, J.C.; Hurwitz, J.L. Differential localization and function of antibody-forming cells responsive to inactivated or live-attenuated influenza virus vaccines. Int. Immunol. 2013, 25, 183–195. [Google Scholar] [CrossRef] [PubMed]
- Hyland, L.; Sangster, M.; Sealy, R.; Coleclough, C. Respiratory virus infection of mice provokes a permanent humoral immune response. J. Virol. 1994, 68, 6083–6086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcin, D.; Pelet, T.; Calain, P.; Roux, L.; Curran, J.; Kolakofsky, D. A highly recombinogenic system for the recovery of infectious Sendai paramyxovirus from cDNA: Generation of a novel copy-back nondefective interfering virus. EMBO J. 1995, 14, 6087–6094. [Google Scholar] [CrossRef]
- Kato, A.; Sakai, Y.; Shioda, T.; Kondo, T.; Nakanishi, M.; Nagai, Y. Initiation of Sendai virus multiplication from transfected cDNA or RNA with negative or positive sense. Genes Cells 1996, 1, 569–579. [Google Scholar] [CrossRef] [PubMed]
- Nagai, Y.; Kato, A. Paramyxovirus reverse genetics is coming of age. Microbiol. Immunol. 1999, 43, 613–624. [Google Scholar] [CrossRef]
- Sakai, Y.; Kiyotani, K.; Fukumura, M.; Asakawa, M.; Kato, A.; Shioda, T.; Yoshida, T.; Tanaka, A.; Hasegawa, M.; Nagai, Y. Accommodation of foreign genes into the Sendai virus genome: Sizes of inserted genes and viral replication. FEBS Lett. 1999, 456, 221–226. [Google Scholar] [CrossRef]
- Mazur, N.I.; Higgins, D.; Nunes, M.C.; Melero, J.A.; Langedijk, A.C.; Horsley, N.; Buchholz, U.J.; Openshaw, P.J.; McLellan, J.S.; Englund, J.A.; et al. Respiratory Syncytial Virus Network, F., The respiratory syncytial virus vaccine landscape: Lessons from the graveyard and promising candidates. Lancet Infect. Dis. 2018, 18, e295–e311. [Google Scholar] [CrossRef] [Green Version]
- Pallesen, J.; Wang, N.; Corbett, K.S.; Wrapp, D.; Kirchdoerfer, R.N.; Turner, H.L.; Cottrell, C.A.; Becker, M.M.; Wang, L.; Shi, W.; et al. Immunogenicity and structures of a rationally designed prefusion MERS-CoV spike antigen. Proc. Natl. Acad. Sci. USA 2017, 114, E7348–E7357. [Google Scholar] [CrossRef] [Green Version]
- Wrapp, D.; Wang, N.; Corbett, K.S.; Goldsmith, J.A.; Hsieh, C.L.; Abiona, O.; Graham, B.S.; McLellan, J.S. Cryo-EM structure of the 2019-nCoV spike in the prefusion conformation. Science 2020, 367, 1260–1263. [Google Scholar] [CrossRef] [Green Version]
- Scaggs Huang, F.; Bernstein, D.I.; Slobod, K.S.; Portner, A.; Takimoto, T.; Russell, C.J.; Meagher, M.; Jones, B.G.; Sealy, R.E.; Coleclough, C.; et al. Safety and immunogenicity of an intranasal sendai virus-based vaccine for human parainfluenza virus type I and respiratory syncytial virus (SeVRSV) in adults. Hum. Vaccin. Immunother. 2021, 17, 554–559. [Google Scholar] [CrossRef]
- Russell, C.J.; Penkert, R.R.; Kim, S.; Hurwitz, J.L. Human Metapneumovirus: A Largely Unrecognized Threat to Human Health. Pathogens 2020, 9, 109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mui, Y.; Lee, B.Y.; Adam, A.; Kharmats, A.Y.; Budd, N.; Nau, C.; Gittelsohn, J. Healthy versus Unhealthy Suppliers in Food Desert Neighborhoods: A Network Analysis of Corner Stores’ Food Supplier Networks. Int. J. Env. Res. Public Health 2015, 12, 15058–15074. [Google Scholar] [CrossRef] [Green Version]
- Jones, B.G.; Oshansky, C.M.; Bajracharya, R.; Tang, L.; Sun, Y.; Wong, S.S.; Webby, R.; Thomas, P.G.; Hurwitz, J.L. Retinol binding protein and vitamin D associations with serum antibody isotypes, serum influenza virus-specific neutralizing activities and airway cytokine profiles. Clin. Exp. Immunol. 2016, 183, 239–247. [Google Scholar] [CrossRef] [Green Version]
- Patel, N.; Penkert, R.R.; Jones, B.G.; Sealy, R.E.; Surman, S.L.; Sun, Y.; Tang, L.; DeBeauchamp, J.; Webb, A.; Richardson, J.; et al. Baseline Serum Vitamin A and D Levels Determine Benefit of Oral Vitamin A&D Supplements to Humoral Immune Responses Following Pediatric Influenza Vaccination. Viruses 2019, 11, 907. [Google Scholar]
- Trasino, S.E.; Tang, X.H.; Jessurun, J.; Gudas, L.J. Obesity Leads to Tissue, but not Serum Vitamin A Deficiency. Sci. Rep. 2015, 5, 15893. [Google Scholar] [CrossRef] [PubMed]
- Neidich, S.D.; Green, W.D.; Rebeles, J.; Karlsson, E.A.; Schultz-Cherry, S.; Noah, T.L.; Chakladar, S.; Hudgens, M.G.; Weir, S.S.; Beck, M.A. Increased risk of influenza among vaccinated adults who are obese. Int. J. Obes. 2017, 41, 1324–1330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Penkert, R.R.; Cortez, V.; Karlsson, E.A.; Livingston, B.; Surman, S.L.; Li, Y.; Catharine Ross, A.; Schultz-Cherry, S.; Hurwitz, J.L. Vitamin A Corrects Tissue Deficits in Diet-Induced Obese Mice and Reduces Influenza Infection After Vaccination and Challenge. Obesity 2020, 28, 1631–1636. [Google Scholar] [CrossRef] [PubMed]
- Penkert, R.R.; Rowe, H.M.; Surman, S.L.; Sealy, R.E.; Rosch, J.; Hurwitz, J.L. Influences of Vitamin A on Vaccine Immunogenicity and Efficacy. Front Immunol. 2019, 10, 1576. [Google Scholar] [CrossRef] [Green Version]
- Penkert, R.R.; Surman, S.L.; Jones, B.G.; Sealy, R.E.; Vogel, P.; Neale, G.; Hurwitz, J.L. Vitamin A deficient mice exhibit increased viral antigens and enhanced cytokine/chemokine production in nasal tissues following respiratory virus infection despite the presence of FoxP3+ T cells. Int. Immunol. 2016, 28, 139–152. [Google Scholar] [CrossRef] [PubMed]
- Surman, S.L.; Jones, B.G.; Rudraraju, R.; Sealy, R.E.; Hurwitz, J.L. Intranasal administration of retinyl palmitate with a respiratory virus vaccine corrects impaired mucosal IgA response in the vitamin A-deficient host. Clin. Vaccine Immunol. 2014, 21, 598–601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Surman, S.L.; Jones, B.G.; Sealy, R.E.; Rudraraju, R.; Hurwitz, J.L. Oral retinyl palmitate or retinoic acid corrects mucosal IgA responses toward an intranasal influenza virus vaccine in vitamin A deficient mice. Vaccine 2014, 32, 2521–2524. [Google Scholar] [CrossRef] [Green Version]
- Sommer, A. Vitamin A, infectious disease, and childhood mortality: A 2 cent solution? J. Infect. Dis. 1993, 167, 1003–1007. [Google Scholar] [CrossRef]
- Keller, H.; Givel, F.; Perroud, M.; Wahli, W. Signaling cross-talk between peroxisome proliferator-activated receptor/retinoid X receptor and estrogen receptor through estrogen response elements. Mol. Endocrinol. 1995, 9, 794–804. [Google Scholar] [PubMed] [Green Version]
- Krycer, J.R.; Brown, A.J. Cross-talk between the androgen receptor and the liver X receptor: Implications for cholesterol homeostasis. J. Biol. Chem. 2011, 286, 20637–20647. [Google Scholar] [CrossRef] [Green Version]
- Evans, R.M.; Mangelsdorf, D.J. Nuclear Receptors, RXR, and the Big Bang. Cell 2014, 157, 255–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, B.G.; Sealy, R.E.; Penkert, R.R.; Surman, S.L.; Birshtein, B.K.; Xu, B.; Neale, G.; Maul, R.W.; Gearhart, P.J.; Hurwitz, J.L. From Influenza Virus Infections to Lupus: Synchronous Estrogen Receptor alpha and RNA Polymerase II Binding within the Immunoglobulin Heavy Chain Locus. Viral Immunol. 2020, 33, 307–315. [Google Scholar] [CrossRef] [PubMed]
- Jones, B.G.; Sealy, R.E.; Penkert, R.R.; Surman, S.L.; Maul, R.W.; Neale, G.; Xu, B.; Gearhart, P.J.; Hurwitz, J.L. Complex sex-biased antibody responses: Estrogen receptors bind estrogen response elements centered within immunoglobulin heavy chain gene enhancers. Int. Immunol. 2019, 31, 141–156. [Google Scholar] [CrossRef] [PubMed]
- Sealy, R.E.; Jones, B.G.; Surman, S.L.; Penkert, R.R.; Pelletier, S.; Neale, G.; Hurwitz, J.L. Will Attention by Vaccine Developers to the Host’s Nuclear Hormone Levels and Immunocompetence Improve Vaccine Success? Vaccines 2019, 7, 26. [Google Scholar] [CrossRef] [Green Version]
- Surman, S.L.; Penkert, R.R.; Sealy, R.E.; Jones, B.G.; Marion, T.N.; Vogel, P.; Hurwitz, J.L. Consequences of Vitamin A Deficiency: Immunoglobulin Dysregulation, Squamous Cell Metaplasia, Infectious Disease, and Death. Int. J. Mol. Sci. 2020, 21, 5570. [Google Scholar] [CrossRef]
- Li, K.; Li, Z.; Wohlford-Lenane, C.; Meyerholz, D.K.; Channappanavar, R.; An, D.; Perlman, S.; McCray, P.B., Jr.; He, B. Single-Dose, Intranasal Immunization with Recombinant Parainfluenza Virus 5 Expressing Middle East Respiratory Syndrome Coronavirus (MERS-CoV) Spike Protein Protects Mice from Fatal MERS-CoV Infection. mBio 2020, 11, e00554-20. [Google Scholar] [CrossRef] [Green Version]
- Lu, M.; Dravid, P.; Zhang, Y.; Trivedi, S.; Li, A.; Harder, O.; Kc, M.; Chaiwatpongsakorn, S.; Zani, A.; Kenney, A.; et al. A safe and highly efficacious measles virus-based vaccine expressing SARS-CoV-2 stabilized prefusion spike. Proc. Natl Acad Sci USA 2021, 118, 12. [Google Scholar] [CrossRef] [PubMed]
- Rossi, S.L.; Comer, J.E.; Wang, E.; Azar, S.R.; Lawrence, W.S.; Plante, J.A.; Ramsauer, K.; Schrauf, S.; Weaver, S.C. Immunogenicity and Efficacy of a Measles Virus-Vectored Chikungunya Vaccine in Nonhuman Primates. J. Infect. Dis 2019, 220, 735–742. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
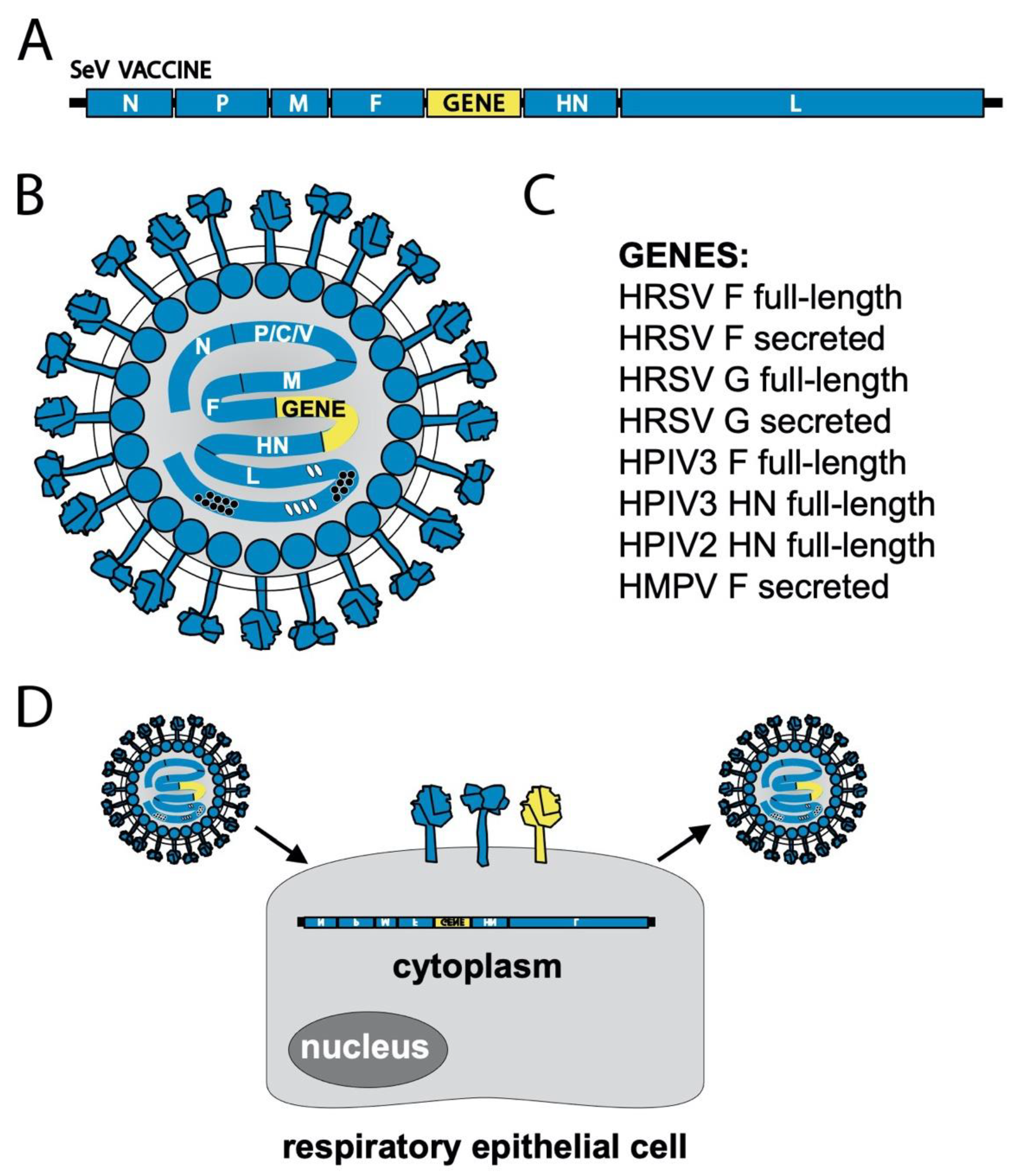{kind=link}
| Common Name | Species | Genus |
|---|---|---|
| Human parainfluenza virus 1 (HPIV1) | Human respirovirus 1 | Respirovirus |
| Human parainfluenza virus 3 (HPIV3) | Human respirovirus 3 | |
| Sendai virus (SeV) | Murine respirovirus | |
| Human parainfluenza virus 2 (HPIV2) | Human orthorubulavirus 2 | Orthorubulavirus |
| Human parainfluenza virus 4 (HPIV4) | Human orthorubulavirus 4 | |
| Parainfluenza virus 5 (PIV5) | Mammalian orthorubulavirus 5 | |
| Mumps virus (MuV) | Mumps orthorubulavirus | |
| Canine distemper virus (CDV) | Canine morbillivirus | Morbillivirus |
| Measles virus (MeV) | Measles morbillivirus | |
| Newcastle disease virus (NDV) | Avian avulavirus 1 | Orthoavulavirus |
| Hendra virus (HeV) | Hendra henipavirus | Henipavirus |
| Nipah virus (NiV) | Nipah henipavirus |
| Common Name | Species | Genus |
|---|---|---|
| Human metapneumovirus (HMPV) | Human metapneumovirus | Metapneumovirus |
| Bovine respiratory syncytial virus (BRSV) | Bovine orthopneumovirus | Orthopneumovirus |
| Human respiratory syncytial virus (HRSV) | Human orthopneumovirus |
| Vaccine Name | Inserted Antigen | Insertion Site | Host | Reference |
|---|---|---|---|---|
| Sendai virus | None | None | cotton rats | [99] |
| Sendai virus | None | None | African green monkeys (AGM) | [100] |
| Sendai virus | None | None | AGM, chimpanzees | [101] |
| Sendai virus | None | None | human (adults/3–6 y.o.) | [102,103] |
| rSV-RSV-G | RSV A2 G | F-HN | cotton rats | [104,105] |
| rSV-RSV-F (SeVRSV) | RSV A2 F | F-HN | cotton rats | [105,106,107,108] |
| rSV-RSV-F (SeVRSV) | RSV A2 F | F-HN | AGM | [109] |
| rSV-RSV-F (SeVRSV) | RSV A2 F | F-HN | human adults | [103] |
| rSV-RSV-Fs | RSV A2 F (secreted) | F-HN | cotton rats | [110] |
| rSV-HPIV3-HN | HPIV3 C243 HN | F-HN | cotton rats | [107,108] |
| rSV-HPIV3-F | HPIV3 C243 F | F-HN | cotton rats | [107,111] |
| rSV-HPIV3-F(P-M) | HPIV3 C243 F | P-M | cotton rats | [111] |
| rSV-HPIV2-HN | HPIV2 VR92 HN | F-HN | cotton rats | [108] |
| rSV-HPIV2-F | HPIV2 VR92 F | F-HN | cotton rats | [108] |
| rSV-HMPV-Fs | HMPV CAN00-16 F (secreted) | F-HN | cotton rats | [112] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Russell, C.J.; Hurwitz, J.L. Sendai Virus-Vectored Vaccines That Express Envelope Glycoproteins of Respiratory Viruses. Viruses 2021, 13, 1023. https://doi.org/10.3390/v13061023
Russell CJ, Hurwitz JL. Sendai Virus-Vectored Vaccines That Express Envelope Glycoproteins of Respiratory Viruses. Viruses. 2021; 13(6):1023. https://doi.org/10.3390/v13061023
Chicago/Turabian StyleRussell, Charles J., and Julia L. Hurwitz. 2021. "Sendai Virus-Vectored Vaccines That Express Envelope Glycoproteins of Respiratory Viruses" Viruses 13, no. 6: 1023. https://doi.org/10.3390/v13061023
APA StyleRussell, C. J., & Hurwitz, J. L. (2021). Sendai Virus-Vectored Vaccines That Express Envelope Glycoproteins of Respiratory Viruses. Viruses, 13(6), 1023. https://doi.org/10.3390/v13061023