
Transcriptome Analysis Reveals the Potential Role of Long Noncoding RNAs in Regulating Fowl Adenovirus Serotype 4-Induced Apoptosis in Leghorn Male Hepatocellular Cells

Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells, Viruses and Antibodies
2.2. Virus Infection
2.3. Immunoblot Analysis
2.4. Transfection of siRNAs and Gene Silencing
2.5. TUNEL Staining
2.6. Flow Cytometry Analysis
2.7. Library Construction and Quality Control
2.8. RNA Sequencing and Data Processing
2.9. Prediction of DE lncRNA Target Genes
2.10. GO and KEGG Pathway Analyses
2.11. qRT-PCR
2.12. Statistical Analysis
3. Results
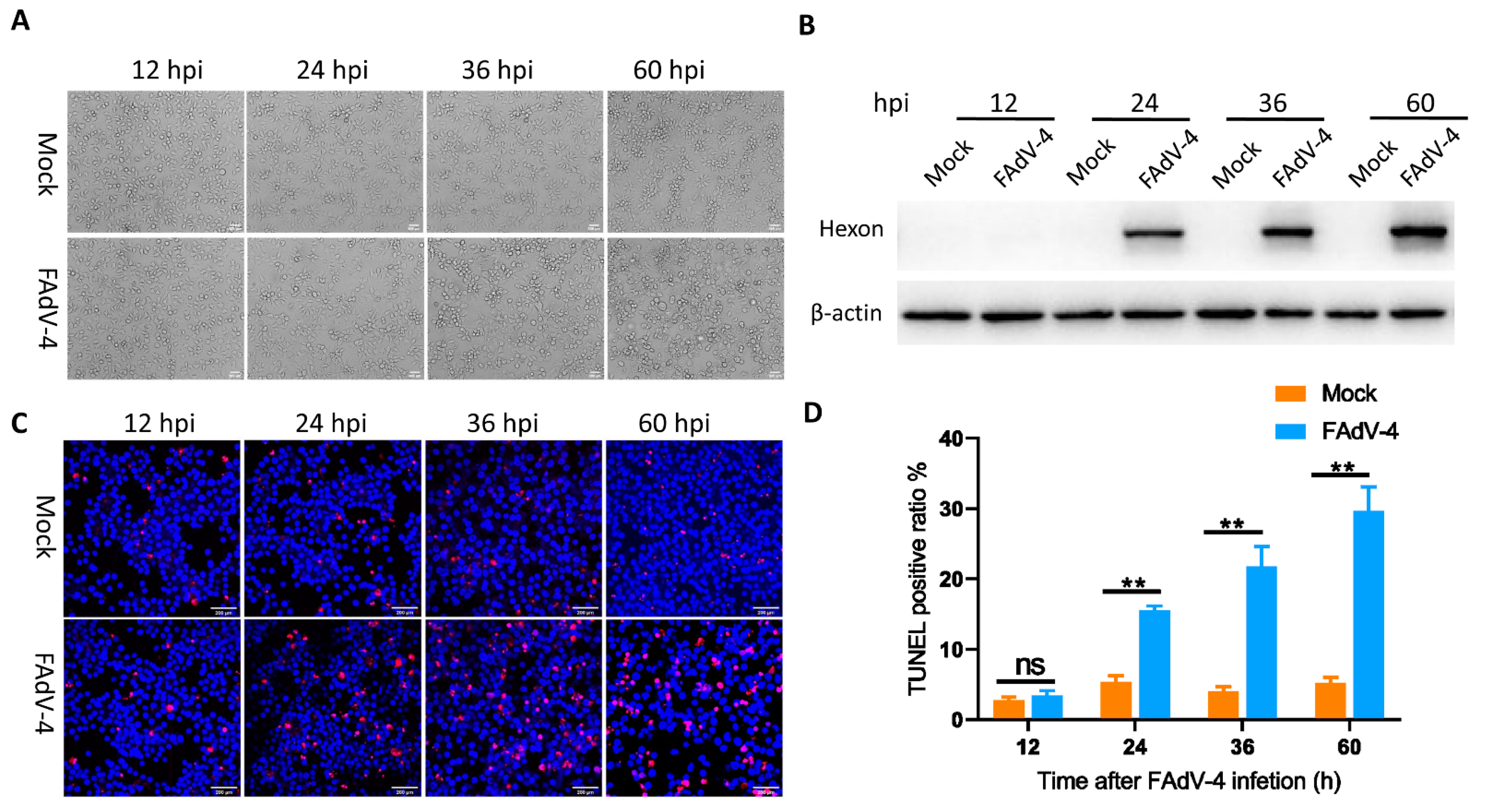
3.1. FAdV-4 Replicates and Induces Apoptosis in LMH Cells
3.2. Overview of RNA Sequencing Data
3.3. Differential Transcription Analysis of lncRNAs and mRNAs
3.4. GO and KEGG Pathway Analyses of DE mRNAs
3.5. Functional Prediction of the DE lncRNAs and Identification of lncRNAs Related to Apoptosis
3.6. Validation of the lncRNA−mRNA Pairs by qRT-PCR
3.7. LncRNA 54128 and BMP4 Are Upregulated during the Course of FAdV-4 Infection
3.8. Effect of Silencing lncRNA 54128 on FAdV-4-Induced Apoptosis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kim, J.N.; Byun, S.H.; Kim, M.J.; Kim, J.; Sung, H.W.; Mo, I.P. Outbreaks of hydropericardium syndrome and molecular characterization of Korean fowl adenoviral isolates. Avian Dis. 2008, 52, 526–530. [Google Scholar] [CrossRef] [PubMed]
- Griffin, B.D.; Nagy, E. Coding potential and transcript analysis of fowl adenovirus 4: Insight into upstream ORFs as common sequence features in adenoviral transcripts. J. Gen. Virol. 2011, 92, 1260. [Google Scholar] [CrossRef]
- Schachner, A.; Marek, A.; Jaskulska, B.; Bilic, I.; Hess, M. Recombinant FAdV-4 fiber-2 protein protects chickens against hepatitis-hydropericardium syndrome (HHS). Vaccine 2014, 32, 1086–1092. [Google Scholar] [CrossRef]
- Pan, Q.; Liu, L.; Wang, Y.; Zhang, Y.; Qi, X.; Liu, C.; Gao, Y.; Wang, X.; Cui, H. The first whole genome sequence and pathogenicity characterization of a fowl adenovirus 4 isolated from ducks associated with inclusion body hepatitis and hydropericardium syndrome. Avian Pathol. 2017, 46, 571–578. [Google Scholar] [CrossRef] [PubMed]
- Hess, M.; Prusas, C.; Vereecken, M.; Herdt, P.D. Isolation of fowl adenoviruses serotype 4 from pigeons with hepatic necrosis. Berl. Münchener Tierärztliche Wochenschr. 1998, 111, 140–142. [Google Scholar]
- Roy, P.; Vairamuthu, S.; Sakthivelan, S.M.; Purushothaman, V. Hydropericardium syndrome in Japanese quail (Coturnix coturnix japonica). Vet. Rec. 2004, 155, 273–274. [Google Scholar] [CrossRef]
- Changjing, L.; Haiying, L.; Dongdong, W.; Jingjing, W.; Youming, W.; Shouchun, W.; Jida, L.; Ping, L.; Jianlin, W.; Shouzhen, X. Characterization of fowl adenoviruses isolated between 2007 and 2014 in China. Vet. Microbiol. 2016, 197, 62–67. [Google Scholar] [CrossRef]
- Abdul-Aziz, T.A.; Al-Attar, M.A. New syndrome in Iraqi chicks. Vet. Rec. 1991, 129, 272. [Google Scholar] [CrossRef]
- Mittal, D.; Jindal, N.; Tiwari, A.K.; Khokhar, R.S. Characterization of fowl adenoviruses associated with hydropericardium syndrome and inclusion body hepatitis in broiler chickens. Virusdisease 2014, 25, 114–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mase, M.; Nakamura, K.; Minami, F. Fowl adenoviruses isolated from chickens with inclusion body hepatitis in Japan, 2009–2010. Avian Pathol. 2012, 74, 1087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toro, H.; Prusas, C.; Raue, R.; Cerda, L.; Geisse, C.; Gonzalez, C.; Hess, M. Characterization of fowl adenoviruses from outbreaks of inclusion body hepatitis/hydropericardium syndrome in Chile. Avian Dis. 1999, 43, 262–270. [Google Scholar] [CrossRef]
- Niu, Y.J.; Xiao, Y.H.; Zhang, G.H.; Sun, H.L.; Qu, Y.J. Hydropericardium syndrome outbreak caused by fowl adenovirus serotype 4 in China in 2015. J. Gen. Virol. 2016, 97, 2684. [Google Scholar] [CrossRef]
- Ye, J.; Liang, G.; Zhang, J.; Wang, W.; Song, N.; Wang, P.; Zheng, W.; Xie, Q.; Shao, H.; Wan, Z.; et al. Outbreaks of serotype 4 fowl adenovirus with novel genotype, China. Emerg. Microbes Infect. 2016, 5, e50. [Google Scholar] [CrossRef]
- Wang, Z.; Zhao, J. Pathogenesis of Hypervirulent Fowl Adenovirus Serotype 4: The Contributions of Viral and Host Factors. Viruses 2019, 11, 741. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.; Jin, Q.; Ding, P.; Wang, Y.; Chai, Y.; Li, Y.; Liu, X.; Luo, J.; Zhang, G. Molecular epidemiology of hydropericardium syndrome outbreak-associated serotype 4 fowl adenovirus isolates in central China. Virol. J. 2016, 13, 188. [Google Scholar] [CrossRef] [Green Version]
- Chandra, R.; Shukla, S.K.; Kumar, M. The hydropericardium syndrome and inclusion body hepatitis in domestic fowl. Trop. Anim. Health Prod. 2000, 32, 99–111. [Google Scholar] [CrossRef]
- Niu, Y.; Sun, Q.; Zhang, G.; Sun, W.; Liu, X.; Xiao, Y.; Shang, Y.; Liu, S. Epidemiological investigation of outbreaks of fowl adenovirus infections in commercial chickens in China. Transbound. Emerg. Dis. 2018, 65, e121–e126. [Google Scholar] [CrossRef]
- Li, P.H.; Zheng, P.P.; Zhang, T.F.; Wen, G.Y.; Luo, Q.P. Fowl adenovirus serotype 4: Epidemiology, pathogenesis, diagnostic detection, and vaccine strategies. Poult. Sci. 2017, 96, 2630–2640. [Google Scholar] [CrossRef]
- Cotter, T.G. Apoptosis and cancer: The genesis of a research field. Nat. Rev. Cancer 2009, 9, 501–507. [Google Scholar] [CrossRef]
- Duan, X.; Zhao, M.; Wang, Y.; Li, X.; Cao, H.; Zheng, S.J. Epigenetic Upregulation of Chicken MicroRNA-16-5p Expression in DF-1 Cells following Infection with Infectious Bursal Disease Virus (IBDV) Enhances IBDV-Induced Apoptosis and Viral Replication. J. Virol. 2020, 94, e01724-19. [Google Scholar] [CrossRef] [PubMed]
- Radke, J.R.; Grigera, F.; Ucker, D.S.; Cook, J.L. Adenovirus E1B 19-kilodalton protein modulates innate immunity through apoptotic mimicry. J. Virol. 2014, 88, 2658–2669. [Google Scholar] [CrossRef] [Green Version]
- Jiang, H.; White, E.J.; Rios-Vicil, C.I.; Xu, J.; Gomez-Manzano, C.; Fueyo, J. Human adenovirus type 5 induces cell lysis through autophagy and autophagy-triggered caspase activity. J. Virol. 2011, 85, 4720–4729. [Google Scholar] [CrossRef] [Green Version]
- Niu, Y.; Sun, Q.; Shi, Y.; Ding, Y.; Li, Z.; Sun, Y.; Li, M.; Liu, S. Immunosuppressive potential of fowl adenovirus serotype 4. Poult. Sci. 2019, 98, 3514–3522. [Google Scholar] [CrossRef] [PubMed]
- Niu, Y.; Sun, Q.; Liu, X.; Liu, S. Mechanism of fowl adenovirus serotype 4-induced heart damage and formation of pericardial effusion. Poult. Sci. 2019, 98, 1134–1145. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Duan, X.; Wang, Y.; Gao, L.; Cao, H.; Li, X.; Zheng, S.J. A Novel Role for PX, a Structural Protein of Fowl Adenovirus Serotype 4 (FAdV4), as an Apoptosis-Inducer in Leghorn Male Hepatocellular Cell. Viruses 2020, 12, 228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, J. The functional role of long non-coding RNAs and epigenetics. Biol. Proced. Online 2014, 16, 11. [Google Scholar] [CrossRef] [Green Version]
- Kapranov, P.; Cheng, J.; Dike, S.; Nix, D.A.; Duttagupta, R.; Willingham, A.T.; Stadler, P.F.; Hertel, J.; Hackermuller, J.; Hofacker, I.L.; et al. RNA maps reveal new RNA classes and a possible function for pervasive transcription. Science 2007, 316, 1484–1488. [Google Scholar] [CrossRef] [Green Version]
- Si, Y.; Bai, J.; Wu, J.; Li, Q.; Mo, Y.; Fang, R.; Lai, W. LncRNA PlncRNA1 regulates proliferation and differentiation of hair follicle stem cells through TGFbeta1mediated Wnt/betacatenin signal pathway. Mol. Med. Rep. 2018, 17, 1191–1197. [Google Scholar]
- Zhao, X.; Su, L.; He, X.; Zhao, B.; Miao, J. Long noncoding RNA CA7-4 promotes autophagy and apoptosis via sponging MIR877-3P and MIR5680 in high glucose-induced vascular endothelial cells. Autophagy 2020, 16, 70–85. [Google Scholar] [CrossRef]
- Li, P.; He, J.; Yang, Z.; Ge, S.; Zhang, H.; Zhong, Q.; Fan, X. ZNNT1 long noncoding RNA induces autophagy to inhibit tumorigenesis of uveal melanoma by regulating key autophagy gene expression. Autophagy 2019, 16, 1–14. [Google Scholar]
- Tsagakis, I.; Douka, K.; Birds, I.; Aspden, J.L. Long non-coding RNAs in development and disease: Conservation to mechanisms. J. Pathol. 2020, 5, 250. [Google Scholar] [CrossRef] [Green Version]
- Du, Z.; Sun, T.; Hacisuleyman, E.; Fei, T.; Wang, X.; Brown, M.; Rinn, J.L.; Lee, M.G.; Chen, Y.; Kantoff, P.W.; et al. Integrative analyses reveal a long noncoding RNA-mediated sponge regulatory network in prostate cancer. Nat. Commun. 2016, 7, 10982. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.; Jiang, Y.; Xu, X.; Su, X.; Liu, Y.; Ma, Y.; Zhao, Y.; Shen, Z.; Huang, B.; Cao, X. Inducible degradation of lncRNA Sros1 promotes IFN-gamma-mediated activation of innate immune responses by stabilizing Stat1 mRNA. Nat. Immunol. 2019, 20, 1621–1630. [Google Scholar] [CrossRef]
- Meng, X.Y.; Luo, Y.; Anwar, M.N.; Sun, Y.; Gao, Y.; Zhang, H.; Munir, M.; Qiu, H.J. Long Non-Coding RNAs: Emerging and Versatile Regulators in Host-Virus Interactions. Front. Immunol. 2017, 8, 1663. [Google Scholar] [CrossRef]
- Wang, P.; Xu, J.; Wang, Y.; Cao, X. An interferon-independent lncRNA promotes viral replication by modulating cellular metabolism. Science 2017, 358, 1051–1055. [Google Scholar] [CrossRef] [Green Version]
- Ouyang, J.; Zhu, X.; Chen, Y.; Wei, H.; Chen, Q.; Chi, X.; Qi, B.; Zhang, L.; Zhao, Y.; Gao, G.F. NRAV, a long noncoding RNA, modulates antiviral responses through suppression of interferon-stimulated gene transcription. Cell Host Microbe 2014, 16, 616–626. [Google Scholar] [CrossRef] [Green Version]
- Maarouf, M.; Chen, B.; Chen, Y.; Wang, X.; Rai, K.R.; Zhao, Z.; Liu, S.; Li, Y.; Xiao, M.; Chen, J.L. Identification of lncRNA-155 encoded by MIR155HG as a novel regulator of innate immunity against influenza A virus infection. Cell Microbiol. 2019, 21, e13036. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zou, Z.; Huang, K.; Lin, X.; Chen, H.; Jin, M. Insights into leghorn male hepatocellular cells response to fowl adenovirus serotype 4 infection by transcriptome analysis. Vet. Microbiol. 2018, 214, 65–74. [Google Scholar] [CrossRef]
- Wu, N.; Yang, B.; Wen, B.; Wang, T.; Wang, J. Interactions Among Expressed MicroRNAs and mRNAs in the Early Stages of Fowl Adenovirus Aerotype 4-Infected Leghorn Male Hepatocellular Cells. Front. Microbiol. 2020, 11, 831. [Google Scholar] [CrossRef]
- Chen, Y.; Huang, R.; Qu, G.; Peng, Y.; Xu, L.; Wang, C.; Huang, C.; Wang, Q. Transcriptome Analysis Reveals New Insight of Fowl Adenovirus Serotype 4 Infection. Front. Microbiol. 2020, 11, 146. [Google Scholar] [CrossRef] [Green Version]
- Ren, G.; Wang, H.; Huang, M.; Yan, Y.; Liu, F.; Chen, R. Transcriptome analysis of fowl adenovirus serotype 4 infection in chickens. Virus Genes 2019, 55, 619–629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; van Baren, M.J.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tafer, H.; Hofacker, I.L. RNAplex: A fast tool for RNA-RNA interaction search. Bioinformatics 2008, 24, 2657–2663. [Google Scholar] [CrossRef]
- Zhao, H.; Dahlo, M.; Isaksson, A.; Syvanen, A.C.; Pettersson, U. The transcriptome of the adenovirus infected cell. Virology 2012, 424, 115–128. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.; Lin, Y.; Wu, J. Long non-coding RNA expression profiling of mouse testis during postnatal development. PLoS ONE 2013, 8, e75750. [Google Scholar] [CrossRef]
- Zhao, J.; Zhong, Q.; Zhao, Y.; Hu, Y.X.; Zhang, G.Z. Pathogenicity and Complete Genome Characterization of Fowl Adenoviruses Isolated from Chickens Associated with Inclusion Body Hepatitis and Hydropericardium Syndrome in China. PLoS ONE 2015, 10, e0133073. [Google Scholar] [CrossRef]
- Liu, W.; Ding, C. Roles of LncRNAs in Viral Infections. Front. Cell Infect. Microbiol. 2017, 7, 205. [Google Scholar] [CrossRef]
- Wang, Y.; Tian, H. miR-101 suppresses HBV replication and expression by targeting FOXO1 in hepatoma carcinoma cell lines. Biochem. Biophys. Res. Commun. 2017, 487, 167–172. [Google Scholar] [CrossRef]
- Oteiza, A.; Mechti, N. FoxO4 negatively controls Tat-mediated HIV-1 transcription through the post-transcriptional suppression of Tat encoding mRNA. J. Gen. Virol. 2017, 98, 1864–1878. [Google Scholar] [CrossRef]
- Vousden, K.H.; Prives, C. Blinded by the Light: The Growing Complexity of p53. Cell 2009, 137, 413–431. [Google Scholar] [CrossRef] [Green Version]
- Levine, A.J. The many faces of p53: Something for everyone. J. Mol. Cell Biol. 2019, 11, 524–530. [Google Scholar] [CrossRef]
- Vousden, K.H.; Lane, D.P. p53 in health and disease. Nat. Rev. Mol. Cell Biol. 2007, 8, 275–283. [Google Scholar] [CrossRef]
- Hall, A.R.; Dix, B.R.; O’Carroll, S.J.; Braithwaite, A.W. p53-dependent cell death/apoptosis is required for a productive adenovirus infection. Nat. Med. 1998, 4, 1068–1072. [Google Scholar] [CrossRef] [PubMed]
- Mihaylova, M.M.; Shaw, R.J. The AMPK signalling pathway coordinates cell growth, autophagy and metabolism. Nat. Cell Biol. 2011, 13, 1016–1023. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.Q.; Yu, Y.; Zhang, X.H.; Floyd, E.Z.; Cefalu, W.T. Human adenovirus 36 decreases fatty acid oxidation and increases de novo lipogenesis in primary cultured human skeletal muscle cells by promoting Cidec/FSP27 expression. Int. J. Obes. 2010, 34, 1355–1364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niu, Y.; Sun, Q.; Zhang, G.; Liu, X.; Shang, Y.; Xiao, Y.; Liu, S. Fowl adenovirus serotype 4-induced apoptosis, autophagy, and a severe inflammatory response in liver. Vet. Microbiol. 2018, 223, 34–41. [Google Scholar] [CrossRef]
- Cao, S.; Reece, E.A.; Shen, W.B.; Yang, P. Restoring BMP4 expression in vascular endothelial progenitors ameliorates maternal diabetes-induced apoptosis and neural tube defects. Cell Death Dis. 2020, 11, 859. [Google Scholar] [CrossRef]
- Rossig, L.; Haendeler, J.; Hermann, C.; Malchow, P.; Urbich, C.; Zeiher, A.M.; Dimmeler, S. Nitric oxide down-regulates MKP-3 mRNA levels: Involvement in endothelial cell protection from apoptosis. J. Biol. Chem. 2000, 275, 25502–25507. [Google Scholar] [CrossRef] [Green Version]
- Lim, H.M.; Lee, J.; Nam, M.J.; Park, S.H. Acetylshikonin Induces Apoptosis in Human Colorectal Cancer HCT-15 and LoVo Cells via Nuclear Translocation of FOXO3 and ROS Level Elevation. Oxidative Med. Cell Longev. 2021, 2021, 6647107. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers | Sequences (5′–3′) |
|---|---|
| ENSGALG00000054128 | GGACACATCTCTTTCTTGCCC |
| ACGTGATTAAGGGCAGCAGAT | |
| ENSGALG00000054172 | GCAGGATGCCAGTTCAGAGT |
| TGCCTTTCTCAGTTATTCCAGT | |
| XLOC_026155 | TGCACTTAGTAGCAGTATAGCCA |
| TTCAAGGTCCTAGCCTCCCA | |
| ENSGALG00000050472 | TGCGGAGTAATAGGTCAGTGAG |
| TCCAGCCTTCTGCATTCTCC | |
| ENSGALG00000048532 | AAGCATGTGGCAACTTCGTA |
| ACTGATCAGCTGCCATTTATGA | |
| BMP4 | CGCTCCTGGTCACCTTCG |
| CAACCCACGTCGCTGAAATC | |
| SOCS2 | GTACCAGGACGGCAAGTTCA |
| TAGAGGTGGACCGTCCCATT | |
| FOXO3 | CCCATGATGTCGTTTGCTGC |
| CCGCTAAGAGGAGAGCTGTG | |
| SLC40A1 | CTAGGGTTGGCCTTTGGTCC |
| GCCTCTTTCAGATTCCGCCA | |
| GAPDH | GAGGGTAGTGAAGGCTGCTG |
| CATCAAAGGTGGAGGAATGG |
| Sample | Raw Reads | Clean Reads | Q30 of Clean Reads (%) | Total Mapped Reads (%) |
|---|---|---|---|---|
| M-1 | 93,753,712 | 93,689,766 | 94.25 | 92.545 |
| M-2 | 107,934,056 | 107,443,430 | 93.77 | 91.440 |
| M-3 | 94,306,864 | 93,891,362 | 94.17 | 90.616 |
| 24P-1 | 108,867,196 | 108,460,480 | 92.28 | 73.424 |
| 24P-2 | 102,718,926 | 102,619,998 | 94.07 | 77.610 |
| 24P-3 | 113,636,874 | 113,200,420 | 93.98 | 73.189 |
| LncRNA_Name | LncRNA_Name Fold Change | mRNA_Name | Target mRNA Fold Change | Correlation |
|---|---|---|---|---|
| XLOC_026155 | −11.64 | FOXO3 | −1.86 | 0.990 |
| ENSGALG00000037194 | 3.99 | RHOB | 1.54 | 0.998 |
| XLOC_003734 | 8.41 | MKP3 | 2.82 | 0.986 |
| ENSGALG00000052384 | 1.59 | TNFRSF18 | 2.19 | 0.986 |
| XLOC_000612 | 3.14 | MKP3 | 2.82 | 0.984 |
| ENSGALG00000037919 | 3.89 | RHOB | 1.54 | 0.983 |
| XLOC_032168 | 2.57 | FOS | 2.81 | 0.979 |
| ENSGALG00000050472 | −1.86 | FOXO3 | −1.86 | 0.978 |
| XLOC_027548 | 9.33 | ENSGALG00000031518 | 2.90 | 0.978 |
| XLOC_033831 | 2.47 | BMP4 | 3.41 | 0.977 |
| ENSGALG00000047387 | 9.09 | CRYAA | 3.06 | 0.975 |
| ENSGALG00000049522 | 4.70 | ENSGALG00000031427 | 2.75 | 0.975 |
| XLOC_034149 | 2.50 | PAX2 | 1.90 | 0.974 |
| ENSGALG00000048532 | −2.46 | SLC40A1 | −1.85 | 0.969 |
| ENSGALG00000050121 | 2.38 | TNFAIP3 | 3.89 | 0.957 |
| XLOC_014249 | 2.30 | HOXA5 | 2.20 | 0.949 |
| ENSGALG00000054128 | 1.95 | BMP4 | 3.41 | 0.935 |
| ENSGALG00000051113 | 3.73 | FASLG | 1.59 | 0.921 |
| XLOC_035991 | 1.72 | NR4A2 | 2.26 | 0.918 |
| XLOC_027850 | 3.89 | ENSGALG00000031518 | 2.90 | 0.910 |
| LncRNA_id | LncRNA_seq Fold Change | Real-Time PCR Fold Change | mRNA Symbol | mRNA_seq Fold Change | Real-Time PCR Fold Change |
|---|---|---|---|---|---|
| ENSGALG00000054128 | 1.95 | 3.56 | BMP4 | 3.41 | 13.7 |
| ENSGALG00000054172 | 2.66 | 4.45 | SOCS2 | 1.26 | 2.82 |
| XLOC_026155 | −11.64 | −2.39 | FOXO3 | −1.86 | −2.51 |
| ENSGALG00000050472 | −1.8 | −1.58 | FOXO3 | −1.86 | −2.51 |
| ENSGALG00000048532 | −2.46 | −3.7 | SLC40A1 | −1.85 | −2.44 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wen, B.; Wang, X.; Yang, L.; Wang, T.; Hou, X.; Qi, X.; Wang, J. Transcriptome Analysis Reveals the Potential Role of Long Noncoding RNAs in Regulating Fowl Adenovirus Serotype 4-Induced Apoptosis in Leghorn Male Hepatocellular Cells. Viruses 2021, 13, 1623. https://doi.org/10.3390/v13081623
Wen B, Wang X, Yang L, Wang T, Hou X, Qi X, Wang J. Transcriptome Analysis Reveals the Potential Role of Long Noncoding RNAs in Regulating Fowl Adenovirus Serotype 4-Induced Apoptosis in Leghorn Male Hepatocellular Cells. Viruses. 2021; 13(8):1623. https://doi.org/10.3390/v13081623
Chicago/Turabian StyleWen, Bo, Xueping Wang, Lulu Yang, Ting Wang, Xiaolan Hou, Xuefeng Qi, and Jingyu Wang. 2021. "Transcriptome Analysis Reveals the Potential Role of Long Noncoding RNAs in Regulating Fowl Adenovirus Serotype 4-Induced Apoptosis in Leghorn Male Hepatocellular Cells" Viruses 13, no. 8: 1623. https://doi.org/10.3390/v13081623
APA StyleWen, B., Wang, X., Yang, L., Wang, T., Hou, X., Qi, X., & Wang, J. (2021). Transcriptome Analysis Reveals the Potential Role of Long Noncoding RNAs in Regulating Fowl Adenovirus Serotype 4-Induced Apoptosis in Leghorn Male Hepatocellular Cells. Viruses, 13(8), 1623. https://doi.org/10.3390/v13081623