
Viruses and Cajal Bodies: A Critical Cellular Target in Virus Infection?



Abstract
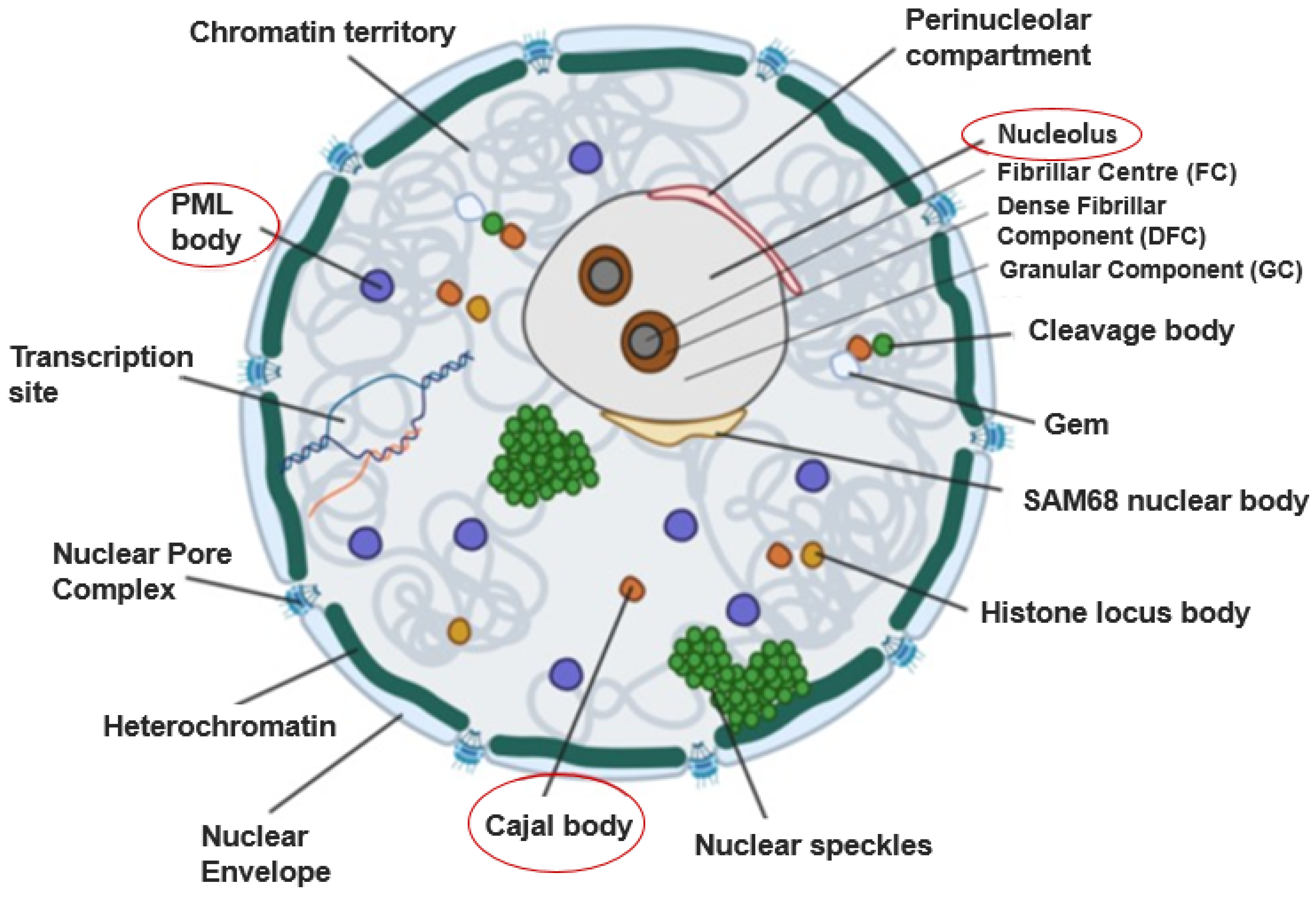
:1. Introduction
1.1. Cajal Bodies
1.2. The Nucleolus
1.3. Promyelocytic Leukaemia (PML) Bodies
1.4. Components of Cajal Bodies
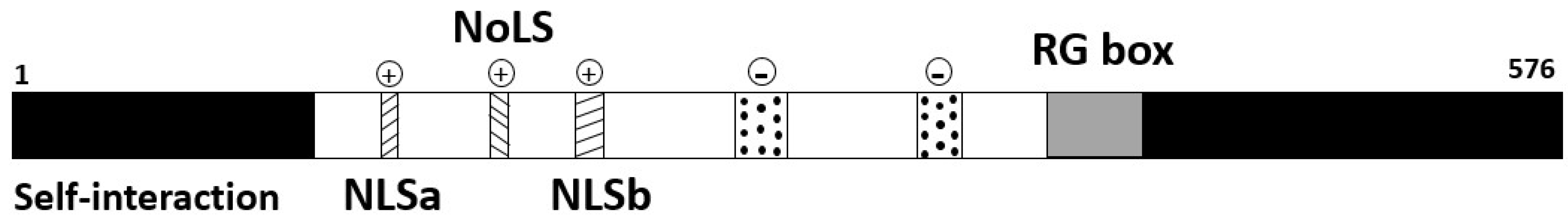
1.4.1. p80-coilin
1.4.2. SMN
1.4.3. TOE1
1.4.4. WRAP53
1.4.5. Nopp140 and Fibrillarin
1.4.6. snRNPs, snoRNPs and scaRNPs
2. Viruses That Interact with Nuclear Bodies
2.1. Animal Viruses Interacting with Nuclear Bodies
2.2. Animal Viruses That Target Cajal Bodies
2.2.1. Minute Virus of Mice
2.2.2. African Swine Fever Virus
2.3. Human Viruses Interacting with Cajal Body Components to Induce Antivirus Defence
Human Immunodeficiency Virus
2.4. Human Viruses Interacting with Cajal Body Components
2.4.1. Influenza A Virus
2.4.2. Zika Virus
2.4.3. Adenoviruses
2.4.4. Herpesviruses
2.5. Plant Viruses Interacting with Nuclear Bodies
2.5.1. Virus Interactions with the Cajal Body Component, Fibrillarin
Potato Virus A
Citrus Tristeza Virus
Groundnut Rosette Virus
2.5.2. Virus Interaction with the Cajal Body Protein Coilin, Resulting in Increased Pathogenicity
Poa Semilatent Virus
2.5.3. Virus Interaction with Cajal Body Marker Protein Coilin, Resulting in Decreased Pathogenicity
Tobacco Rattle Virus
Barley Stripe Mosaic Virus
Rice Stripe Virus
Grapevine Red Blotch-Associated Virus
2.5.4. Targeting Argonaute 4
Tomato Yellow Leaf Curl Virus
Cucumber Mosaic Virus
Pelargonium Line Pattern Virus
3. Concluding Remarks: Common Threads and Future Perspectives
3.1. Nuclear Architecture Can Be Remodelled in Similar Ways by Diverse Viruses
3.2. Involvement of Nuclear Bodies in Antivirus Defence
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sharp, P.A.; Chakraborty, A.K.; Henninger, J.E.; Young, R.A. RNA in formation and regulation of transcriptional condensates. RNA 2022, 28, 52–57. [Google Scholar] [CrossRef] [PubMed]
- Collier, S.; Pendle, A.; Boudonck, K.; van Rij, T.; Dolan, L.; Shaw, P. A distant coilin homologue is required for the formation of Cajal bodies in Arabidopsis. Mol. Biol. Cell 2006, 17, 2942–2951. [Google Scholar] [CrossRef] [PubMed]
- Strzelecka, M.; Trowitzsch, S.; Weber, G.; Lührmann, R.; Oates, A.C.; Neugebauer, K.M. Coilin-dependent snRNP assembly is essential for zebrafish embryogenesis. Nat. Struct. Mol. Biol. 2010, 17, 403–409. [Google Scholar] [CrossRef] [PubMed]
- Tucker, K.E.; Berciano, M.T.; Jacobs, E.Y.; LePage, D.F.; Shpargel, K.B.; Rossire, J.J.; Matera, A.G. Residual Cajal bodies in coilin knockout mice fail to recruit Sm snRNPs and SMN, the spinal muscular atrophy gene product. J. Cell Biol. 2001, 154, 293–308. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Dreyfuss, G. A novel nuclear structure containing the survival of motor neurons protein. EMBO J. 1996, 15, 3555–3565. [Google Scholar] [CrossRef]
- Fong, K.W.; Li, Y.; Wang, W.; Ma, W.; Li, K.; Qi, R.Z.; Chen, J. Whole-genome screening identifies proteins localized to distinct nuclear bodies. J. Cell Biol. 2013, 203, 149–164. [Google Scholar] [CrossRef]
- Mahmoudi, S.; Henriksson, S.; Weibrecht, I.; Smith, S.; Söderberg, O.; Strömblad, S.; Wiman, K.G.; Farnebo, M. WRAP53 is essential for Cajal body formation and for targeting the survival of motor neuron complex to Cajal bodies. PLoS Biol. 2010, 8, e1000521. [Google Scholar] [CrossRef]
- Isaac, C.; Yang, Y.; Thomas, M.U. Nopp140 functions as a molecular link between the nucleolus and the coiled bodies. J. Cell Biol. 1998, 142, 319–329. [Google Scholar] [CrossRef]
- Jones, K.; Gorzynski, K.; Hales, C.; Fischer, U.; Badbanchi, F.; Terns, R.; Terns, M. Direct interaction of the spinal muscular atrophy disease protein SMN with the small nucleolar RNA-associated protein fibrillarin. J. Cell Biol. 2001, 276, 38645–38651. [Google Scholar] [CrossRef]
- Meier, U.T.; Blobel, G. Nopp 140 shuttles on tracks between nucleolus and cytoplasm. Cell 1992, 70, 127–138. [Google Scholar] [CrossRef]
- Pellizzoni, L.; Baccon, J.; Charroux, B.; Dreyfuss, G.; Pellizzoni, L. The survival of motor neurons (SMN) protein interacts with the snoRNP proteins fibrillarin and GAR1. Curr. Biol. 2001, 11, 1079–1088. [Google Scholar] [CrossRef] [PubMed]
- Rampersad, S.; Tennant, P. Replication and expression strategies of viruses. In Viruses. Molecular Biology, Host Interactions and Applications to Biotechnology; Tennant, P., Fermin, B., Foster, J.E., Eds.; Academic Press: Cambridge, MA, USA, 2018; pp. 55–82. [Google Scholar]
- Wang, P.; Benhenda, S.; Wu, H.; Lallemand-Breitenbach, V.; Zhen, T.; Jollivet, F.; Peres, L.; Li, Y.; Chen, S.; Chen, Z.; et al. RING tetramerization is required for nuclear body biogenesis and PML sumoylation. Nat. Commun. 2018, 9, 1277. [Google Scholar] [CrossRef] [PubMed]
- Cajal, S.R. A simple method of selective staining of the protoplasmic reticulum and its effects on the various nervous organs of vertebrates and invertebrates. Trab. Lab. Invest. Biol. 1903, 2, 129–221. [Google Scholar]
- Lafarga, M.; Herv’s, J.; Santa-Cruz, M.; Villegas, J.; Crespo, D. The “Accessory body” of Cajal in the neuronal nucleus. Anat. Embryol. 1983, 166, 19–30. [Google Scholar] [CrossRef] [PubMed]
- Cioce, M.; Lamond, A.I. Cajal bodies: A long history of discovery. Annu. Rev. Cell Dev. Biol. 2005, 21, 105–131. [Google Scholar] [CrossRef] [PubMed]
- Young, P.; Le, T.; Dunckley, M.; Thi Man, N.; Burghes, A.; Morris, G. Nuclear gems and Cajal (coiled) bodies in fetal tissues: Nucleolar distribution of the spinal muscular atrophy protein, SMN. Exp. Cell Res. 2001, 265, 252–261. [Google Scholar] [CrossRef]
- Young, P.; Le, T.; Thi Man, N.; Burghes, A.; Morris, G. The relationship between SMN, the spinal muscular atrophy protein, and nuclear coiled bodies in differentiated tissues and cultured cells. Exp. Cell Res. 2000, 256, 365–374. [Google Scholar] [CrossRef] [PubMed]
- Boudonck, K.; Dolan, L.; Shaw, P. The movement of coiled bodies visualized in living plant cells by the green fluorescent protein. Mol. Biol. Cell 1999, 10, 2297–2307. [Google Scholar] [CrossRef]
- Platani, M.; Goldberg, I.; Swedlow, J.; Lamond, A. In vivo analysis of Cajal body movement, separation, and joining in live human cells. J. Cell Biol. 2000, 151, 1561–1574. [Google Scholar] [CrossRef]
- Andrade, L.E.; Tan, E.M.; Chan, E.K. Immunocytochemical analysis of the coiled body in the cell cycle and during cell proliferation. Proc. Natl. Acad. Sci. USA 1993, 90, 1947–1951. [Google Scholar] [CrossRef]
- Cantarero, L.; Sanz-García, M.; Vinograd-Byk, H.; Renbaum, P.; Levy-Lahad, E.; Lazo, P. VRK1 regulates Cajal body dynamics and protects coilin from proteasomal degradation in cell cycle. Sci. Rep. 2015, 5, 10543. [Google Scholar] [CrossRef] [PubMed]
- Cioce, M.; Boulon, S.; Matera, A.G.; Lamond, A.I. UV-induced fragmentation of Cajal bodies. J. Cell Biol. 2006, 175, 401–413. [Google Scholar] [CrossRef] [PubMed]
- Satyanarayana, A.; Manns, M.; Rudolph, K. Telomeres, telomerase and cancer: An endless search to target the ends. Cell Cycle 2004, 3, 1136–1148. [Google Scholar] [CrossRef]
- Love, A.; Yu, C.; Petukhova, N.; Kalinina, N.; Chen, J.; Taliansky, M. Cajal bodies and their role in plant stress and disease responses. RNA Biol. 2016, 14, 779–790. [Google Scholar] [CrossRef] [PubMed]
- Kroiss, M.; Schultz, J.; Wiesner, J.; Chari, A.; Sickmann, A.; Fischer, U. Evolution of an RNP assembly system: A minimal SMN complex facilitates formation of UsnRNPs in Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 2008, 105, 10045–10050. [Google Scholar] [CrossRef]
- Barneche, F.; Steinmetz, F.; Echeverrıa, M. Fibrillarin genes encode both a conserved nucleolar protein and a novel small nucleolar RNA involved in ribosomal RNA methylation in Arabidopsis thaliana. J. Biol. Chem. 2000, 275, 27212–27220. [Google Scholar] [CrossRef]
- Beven, A.F.; Simpson, G.G.; Brown, J.W.; Shaw, P.J. The organization of spliceosomal components in the nuclei of higher plants. J. Cell Sci. 1995, 108, 509–518. [Google Scholar] [CrossRef]
- Bassett, C.L. Cajal bodies and plant RNA metabolism. Crit. Rev. Plant Sci. 2012, 31, 258–270. [Google Scholar] [CrossRef]
- Kim, S.; Spensley, M.; Choi, S.; Calixto, C.; Pendle, A.; Koroleva, O.; Shaw, P.; Brown, J. Plant U13 orthologues and orphan snoRNAs identified by RNomics of RNA from Arabidopsis nucleoli. Nucleic Acids Res. 2010, 38, 3054–3067. [Google Scholar] [CrossRef]
- Li, C.F.; Pontes, O.; El-Shami, M.; Henderson, I.R.; Bernatavichute, Y.V.; Chan, S.W.L.; Lagrange, T.; Pikaard, C.S.; Jacobsen, S.E. An ARGONAUTE4-containing nuclear processing center colocalized with Cajal bodies in Arabidopsis thaliana. Cell 2006, 126, 93–106. [Google Scholar] [CrossRef]
- Taliansky, M.E.; Love, A.J.; Kołowerzo-Lubnau, A.; Smoliński, D.J. Cajal bodies: Evolutionarily conserved nuclear biomolecular condensates with properties unique to plants. Plant Cell 2023, 35, 3214–3235. [Google Scholar] [CrossRef]
- Scheer, U.; Hock, R. Structure and function of the nucleolus. Curr. Opin. Cell Biol. 1999, 11, 385–390. [Google Scholar] [CrossRef] [PubMed]
- Iarovaia, O.V.; Minina, E.P.; Sheval, E.V.; Onichtchouk, D.; Dokudovskaya, S.; Razin, S.V.; Vassetzky, Y.S. Nucleolus: A central hub for nuclear functions. Trends Cell Biol. 2019, 29, 647–659. [Google Scholar] [CrossRef] [PubMed]
- Boulon, S.; Westman, B.J.; Hutten, S.; Boisvert, F.M.; Lamond, A.I. The nucleolus under stress. Mol. Cell 2010, 40, 216–227. [Google Scholar] [CrossRef] [PubMed]
- Guldner, H.H.; Szostecki, C.; Vosberg, H.P.; Lakomek, H.J.; Penner, E.; Bautz, F.A. Scl 70 autoantibodies from scleroderma patients recognize a 95 kDa protein identified as DNA topoisomerase I. Chromosoma 1986, 94, 132–138. [Google Scholar] [CrossRef] [PubMed]
- Reimer, G.; Rose, K.M.; Scheer, U.; Tan, E.M. Autoantibody to RNA polymerase I in scleroderma sera. J. Clin. Investig. 1987, 79, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Masson, C.; Andre, C.; Arnoult, J.; Geraud, G.; Hernandez-Verdun, D.A. 116000 Mr nucleolar antigen specific for the dense fibrillar component of the nucleoli. J. Cell Sci. 1990, 95, 371–381. [Google Scholar] [CrossRef]
- Padeken, J.; Heun, P. Nucleolus and nuclear periphery: Velcro for heterochromatin. Curr. Opin. Cell Biol. 2014, 28, 54–60. [Google Scholar] [CrossRef]
- Guarente, L. Link between aging and the nucleolus. Genes Dev. 1997, 11, 2449–2455. [Google Scholar] [CrossRef]
- Visintin, R.; Hwang, E.S.; Amon, A. Cfi1 prevents premature exit from mitosis by anchoring Cdc14 phosphatase in the nucleolus. Nature 1999, 398, 818–823. [Google Scholar] [CrossRef]
- Weber, J.D.; Taylor, L.J.; Roussel, M.F.; Sherr, C.J.; Bar-Sagi, D. Nucleolar Arf sequesters Mdm2 and activates p53. Nat. Cell Biol. 1999, 1, 20–26. [Google Scholar]
- Daniely, Y.; Borowiec, J.A. Formation of a complex between nucleolin and replication protein A after cell stress prevents initiation of DNA replication. J. Cell Biol. 2000, 149, 799–810. [Google Scholar] [CrossRef] [PubMed]
- Higashiura, M.; Shimizu, Y.; Tanimoto, M.; Morita, T.; Yagura, T. Immunolocalization of Ku-proteins (p80/p70): Localization of p70 to nucleoli and periphery of both interphase nuclei and metaphase chromosomes. Exp. Cell Res. 1992, 201, 444–451. [Google Scholar] [CrossRef] [PubMed]
- Boisvert, F.M.; Van Koningsbruggen, S.; Navascués, J.; Lamond, A.I. The multifunctional nucleolus. Nat. Rev. Mol. Cell Biol. 2007, 8, 574–585. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, L.M.; Baserga, S.J. Crosstalk between the nucleolus and the DNA damage response. Mol. Biosyst. 2017, 13, 443–455. [Google Scholar] [CrossRef] [PubMed]
- Govoni, M.; Farabegoli, F.; Pession, A.; Novello, F. Inhibition of Topoisomerase II activity and its effect on nucleolar structure and function. Exp. Cell Res. 1994, 211, 36–41. [Google Scholar] [CrossRef]
- Al-Baker, E.A.; Boyle, J.; Harry, R.; Kill, I.R. A p53-independent pathway regulates nucleolar segregation and antigen translocation in response to DNA damage induced by UV irradiation. Exp. Cell Res. 2004, 292, 179–186. [Google Scholar] [CrossRef]
- Miller, M.L.; Andringa, A.; Dixon, K.; Carty, M.P. Insights into UV-induced apoptosis: Ultrastructure, trichrome stain and spectral imaging. Micron 2002, 33, 157–166. [Google Scholar] [CrossRef]
- Hernandez-Verdun, D. Nucleolus: From structure to dynamics. Histochem. Cell Biol. 2005, 125, 127–137. [Google Scholar] [CrossRef]
- Lallemand-Breitenbach, V.; de Thé, H. PML nuclear bodies: From architecture to function. Curr. Opin. Cell Biol. 2018, 52, 154–161. [Google Scholar] [CrossRef]
- Zhong, S.; Müller, S.; Ronchetti, S.; Freemont, P.; Dejean, A.; Pandolfi, P. Role of SUMO-1–modified PML in nuclear body formation. Blood 2000, 95, 2748–2752. [Google Scholar] [CrossRef]
- Borden, K.; Lally, J.; Martin, S.; O’Reilly, N.; Solomon, E.; Freemont, P. In vivo and in vitro characterization of the B1 and B2 zinc-binding domains from the acute promyelocytic leukemia protooncoprotein PML. Proc. Natl. Acad. Sci. USA 1996, 93, 1601–1606. [Google Scholar] [CrossRef]
- Sahin, U.; Ferhi, O.; Jeanne, M.; Benhenda, S.; Berthier, C.; Jollivet, F.; Niwa-Kawakita, M.; Faklaris, O.; Setterblad, N.; de Thé, H.; et al. Oxidative stress–induced assembly of PML nuclear bodies controls sumoylation of partner proteins. J. Cell Biol. 2014, 204, 931–945. [Google Scholar] [CrossRef]
- Wang, Z.; Ruggero, D.; Ronchetti, S.; Zhong, S.; Gaboli, M.; Rivi, R.; Pandolfi, P. PML is essential for multiple apoptotic pathways. Nat. Genet. 1998, 20, 266–272. [Google Scholar] [CrossRef] [PubMed]
- Regad, T.; Chelbi-Alix, M.K. Role and fate of PML nuclear bodies in response to interferon and viral infections. Oncogene 2001, 20, 7274–7286. [Google Scholar] [CrossRef]
- Raška, I.; Andrade, L.; Ochs, R.; Chan, E.; Chang, C.; Roos, G.; Tan, E. Immunological and ultrastructural studies of the nuclear coiled body with autoimmune antibodies. Exp. Cell Res. 1991, 195, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Andrade, L.E.; Chan, E.K.; Raska, I.; Peebles, C.L.; Roos, G.; Tan, E.M. Human autoantibody to a novel protein of the nuclear coiled body: Immunological characterization and cDNA cloning of p80-coilin. J. Exp. Med. 1991, 173, 1407–1419. [Google Scholar] [CrossRef] [PubMed]
- Walker, M.P.; Tian, L.; Matera, A.G. Reduced viability, fertility and fecundity in mice lacking the Cajal body marker protein, coilin. PLoS ONE 2009, 4, e6171. [Google Scholar] [CrossRef]
- Carmo-Fonseca, M.; Ferreira, J.; Lamond, A. Assembly of snRNP-containing coiled bodies is regulated in interphase and mitosis evidence that the coiled body is a kinetic nuclear structure. J. Cell Biol. 1993, 120, 841–852. [Google Scholar] [CrossRef]
- Bohmann, K.; Ferreira, J.; Lamond, A. Mutational analysis of p80 coilin indicates a functional interaction between coiled bodies and the nucleolus. J. Cell Biol. 1995, 131, 817–831. [Google Scholar] [CrossRef]
- Hebert, M.D.; Matera, A.G. Self-association of coilin reveals a common theme in nuclear body localization. Mol. Biol. Cell 2000, 11, 4159–4171. [Google Scholar] [CrossRef] [PubMed]
- Hebert, M.; Szymczyk, P.; Shpargel, K.; Matera, A. Coilin forms the bridge between Cajal bodies and SMN, the spinal muscular atrophy protein. Genes Dev. 2001, 15, 2720–2729. [Google Scholar] [CrossRef]
- Meister, G.; Bühler, D.; Laggerbauer, B.; Zobawa, M.; Lottspeich, F.; Fischer, U. Characterization of a nuclear 20S complex containing the survival of motor neurons (SMN) protein and a specific subset of spliceosomal Sm proteins. Hum. Mol. Genet. 2000, 9, 1977–1986. [Google Scholar] [CrossRef] [PubMed]
- Gubitz, A.; Feng, W.; Dreyfuss, G. The SMN complex. Exp. Cell Res. 2004, 296, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Pellizzoni, L.; Kataoka, N.; Charroux, B.; Dreyfuss, G. A novel function for SMN, the spinal muscular atrophy disease gene product, in pre-mRNA splicing. Cell 1998, 95, 615–624. [Google Scholar] [CrossRef] [PubMed]
- Sleeman, J.E.; Trinkle-Mulcahy, L.; Prescott, A.R.; Ogg, S.C.; Lamond, A.I. Cajal body proteins SMN and coilin show differential dynamic behaviour in vivo. J. Cell Sci. 2003, 116, 2039–2050. [Google Scholar] [CrossRef] [PubMed]
- Lanfranco, M.; Vassallo, N.; Cauchi, R.J. Spinal Muscular Atrophy: From defective chaperoning of snRNP assembly to neuromuscular dysfunction. Front. Mol. Biosci. 2017, 4, 41. [Google Scholar] [CrossRef]
- Young, P.J.; Day, P.M.; Zhou, J.; Androphy, E.J.; Morris, G.E.; Lorson, C.L. A direct interaction between the survival motor neuron protein and p53 and its relationship to spinal muscular atrophy. J. Biol. Chem. 2002, 277, 2852–2859. [Google Scholar] [CrossRef]
- De Belle, I.; Wu, J.X.; Sperandio, S.; Mercola, D.; Adamson, E.D. In vivo cloning and characterization of a new growth suppressor protein TOE1 as a direct target gene of Egr1. J. Biol. Chem. 2003, 278, 14306–14312. [Google Scholar] [CrossRef]
- Mahmoudi, S.; Henriksson, S.; Corcoran, M.; Méndez-Vidal, C.; Wiman, K.; Farnebo, M. Wrap53, a natural p53 antisense transcript required for p53 induction upon DNA damage. Mol. Cell 2009, 33, 462–471. [Google Scholar] [CrossRef]
- Henriksson, S.; Farnebo, M. On the road with WRAP53β: Guardian of Cajal bodies and genome integrity. Front. Genet. 2015, 6, 91. [Google Scholar] [CrossRef] [PubMed]
- Venteicher, A.; Abreu, E.; Meng, Z.; McCann, K.; Terns, R.; Veenstra, T.; Terns, M.; Artandi, S. A human telomerase holoenzyme protein required for Cajal body localization and telomere synthesis. Science 2009, 323, 644–648. [Google Scholar] [CrossRef] [PubMed]
- Machyna, M.; Kehr, S.; Straube, K.; Kappei, D.; Buchholz, F.; Butter, F.; Ule, J.; Hertel, J.; Stadler, P.; Neugebauer, K. The coilin interactome identifies hundreds of small noncoding RNAs that traffic through Cajal bodies. Mol. Cell 2014, 56, 389–399. [Google Scholar] [CrossRef] [PubMed]
- Staněk, D. Cajal bodies and snRNPs-friends with benefits. RNA Biol. 2017, 14, 671–679. [Google Scholar] [CrossRef]
- Carmo-Fonseca, M.; Pepperkok, R.; Sproat, B.; Ansorge, W.; Swanson, M.; Lamond, A. In vivo detection of snRNP-rich organelles in the nuclei of mammalian cells. EMBO J. 1991, 10, 1863–1873. [Google Scholar] [CrossRef] [PubMed]
- Matera, A.G.; Ward, D.C. Nucleoplasmic organization of small nuclear ribonucleoproteins in cultured human cells. J. Cell Biol. 1993, 121, 715–727. [Google Scholar] [CrossRef] [PubMed]
- Burke, M.; Logan, M.; Hebert, M. Identification of additional regulatory RNPs that impact rRNA and U6 snRNA methylation. Biol. Open 2018, 7, bio036095. [Google Scholar] [CrossRef] [PubMed]
- Darzacq, X.; Jády, B.E.; Verheggen, C.; Kiss, A.M.; Bertrand, E.; Kiss, T. Cajal body-specific small nuclear RNAs: A novel class of 2′-O-methylation and pseudouridylation guide RNAs. EMBO J. 2002, 21, 2746–2756. [Google Scholar] [CrossRef]
- Jády, B.E.; Kiss, T. A small nucleolar guide RNA functions both in 2′-O-ribose methylation and pseudouridylation of the U5 spliceosomal RNA. EMBO J. 2001, 20, 541–551. [Google Scholar] [CrossRef]
- Kiss, A.M.; Jády, B.E.; Darzacq, X.; Verheggen, C.; Bertrand, E.; Kiss, T. A Cajal body-specific pseudouridylation guide RNA is composed of two box H/ACA snoRNA-like domains. Nucleic Acids Res. 2002, 30, 4643–4649. [Google Scholar] [CrossRef]
- Tycowski, K.; You, Z.; Graham, P.; Steitz, J. Modification of U6 spliceosomal RNA is guided by other small RNAs. Mol. Cell 1998, 2, 629–638. [Google Scholar] [CrossRef] [PubMed]
- Ganot, P.; Jády, B.; Bortolin, M.; Darzacq, X.; Kiss, T. Nucleolar factors direct the 2′-O-Ribose methylation and pseudouridylation of U6 spliceosomal RNA. Mol. Cell. Biol. 1999, 19, 6906–6917. [Google Scholar] [CrossRef]
- Richard, P.; Darzacq, X.; Bertrand, E.; Jady, B.; Verheggen, C.; Kiss, T. A common sequence motif determines the Cajal body-specific localization of box H/ACA scaRNAs. EMBO J. 2003, 22, 4283–4293. [Google Scholar] [CrossRef] [PubMed]
- Tycowski, K.; Shu, M.; Kukoyi, A.; Steitz, J. A conserved WD40 protein binds the Cajal body localization signal of scaRNP particles. Mol. Cell 2009, 34, 47–57. [Google Scholar] [CrossRef] [PubMed]
- Fischer, U.; Englbrecht, C.; Chari, A. Biogenesis of spliceosomal small nuclear ribonucleoproteins. Wiley Interdiscip. Rev. RNA 2011, 2, 718–731. [Google Scholar] [CrossRef]
- Skaar, J.; Ferris, A.; Wu, X.; Saraf, A.; Khanna, K.; Florens, L.; Washburn, M.; Hughes, S.; Pagano, M. The integrator complex controls the termination of transcription at diverse classes of gene targets. Cell Res. 2015, 25, 288–305. [Google Scholar] [CrossRef]
- Takata, H.; Nishijima, H.; Maeshima, K.; Shibahara, K. The integrator complex is required for integrity of Cajal bodies. J. Cell Sci. 2012, 125, 166–175. [Google Scholar] [CrossRef]
- Matera, A.G.; Wang, Z. A day in the life of the spliceosome. Nat. Rev. Mol. Cell Biol. 2014, 15, 108–121. [Google Scholar] [CrossRef]
- Jady, B.; Darzacq, X.; Tucker, K.; Matera, A.; Bertrand, E.; Kiss, T. Modification of Sm small nuclear RNAs occurs in the nucleoplasmic Cajal body following import from the cytoplasm. EMBO J. 2003, 22, 1878–1888. [Google Scholar] [CrossRef]
- Tsukaya, H.; Byrne, M.E.; Horiguchi, G.; Sugiyama, M.; Van Lijsebettens, M.; Lenhard, M. How do ‘housekeeping’ genes control organogenesis?—Unexpected new findings on the role of housekeeping genes in cell and organ differentiation. J. Plant Res. 2013, 126, 3–15. [Google Scholar] [CrossRef]
- Anobile, J.M.; Arumugaswami, V.; Downs, D.; Czymmek, K.; Parcells, M.; Schmidt, C.J. Nuclear localization and dynamic properties of the Marek’s Disease virus oncogene products Meq and Meq/vIL8. Virol. J. 2006, 80, 1160–1166. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Lee, L.; Ye, Y.; Qian, Z.; Kung, H. Nucleolar and nuclear localization properties of a herpesvirus bZIP oncoprotein MEQ. Virol. J. 1997, 71, 3188–3196. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, T.; Seeler, J.; Ohman, K.; Jordan, P.; Pettersson, U.; Akusjärvi, G.; Carmo-Fonseca, M.; Dejean, A. Targeting of adenovirus E1A and E4-ORF3 proteins to nuclear matrix-associated PML bodies. J. Cell Biol. 1995, 131, 45–56. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, S.H.; Silva, N.P.; Delício, L.R.; Granato, C.; Andrade, L.E. The behaviour of the coiled body in cells infected with adenovirus in vitro. Mol. Biol. Rep. 1996, 23, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Gedge, L.; Morrison, E.; Blair, G.; Walker, J. Nuclear actin is partially associated with Cajal bodies in human cells in culture and relocates to the nuclear periphery after infection of cells by adenovirus 5. Exp. Cell Res. 2005, 303, 229–239. [Google Scholar] [CrossRef]
- James, N.; Howell, G.; Walker, J.; Blair, G. The role of Cajal bodies in the expression of late phase adenovirus proteins. Virology 2010, 399, 299–311. [Google Scholar] [CrossRef] [PubMed]
- White, L.; Erbay, B.; Blair, G.E. The Cajal Body protein p80-coilin forms a complex with the adenovirus L4-22K protein and facilitates the nuclear export of adenovirus mRNA. mBio 2023, 14, e01459–e01482. [Google Scholar] [CrossRef]
- Coyaud, E.; Ranadheera, C.; Cheng, D.; Gonçalves, J.; Dyakov, B.; Laurent, E.; St-Germain, J.; Pelletier, L.; Gingras, A.; Brumell, J.; et al. Global interactomics uncovers extensive organellar targeting by Zika virus. Mol. Cell. Proteom. 2018, 17, 2242–2255. [Google Scholar] [CrossRef]
- Young, P.; Jensen, K.; Burger, L.; Pintel, D.; Lorson, C. Minute Virus of Mice NS1 interacts with the SMN protein, and they colocalize in novel nuclear bodies induced by Parvovirus infection. Virol. J. 2002, 76, 3892–3904. [Google Scholar] [CrossRef]
- Salsman, J.; Zimmerman, N.; Chen, T.; Domagala, M.; Frappier, L. Genome-wide screen of three Herpesviruses for protein subcellular localization and alteration of PML nuclear bodies. PLoS Pathog. 2008, 4, e1000100. [Google Scholar] [CrossRef]
- Davey, J.; Colman, A.; Dimmock, N. Location of Influenza Virus M, NP and NS1 proteins in microinjected cells. J. Gen. Virol. 1985, 66, 2319–2334. [Google Scholar] [CrossRef] [PubMed]
- Fortes, P.; Lamond, A.; Ortin, J. Influenza virus NS1 protein alters the subnuclear localization of cellular splicing components. J. Gen. Virol. 1995, 76, 1001–1007. [Google Scholar] [CrossRef] [PubMed]
- Höfer, C.; Jolmes, F.; Haralampiev, I.; Veit, M.; Herrmann, A. Influenza A virus nucleoprotein targets subnuclear structures. Cell. Microbiol. 2016, 19, e12679. [Google Scholar] [CrossRef] [PubMed]
- Murayama, R.; Harada, Y.; Shibata, T.; Kuroda, K.; Hayakawa, S.; Shimizu, K.; Tanaka, T. Influenza A virus non-structural protein 1 (NS1) interacts with cellular multifunctional protein nucleolin during infection. Biochem. Biophys. Res. Commun. 2007, 362, 880–885. [Google Scholar] [CrossRef] [PubMed]
- Ozawa, M.; Fujii, K.; Muramoto, Y.; Yamada, S.; Yamayoshi, S.; Takada, A.; Goto, H.; Horimoto, T.; Kawaoka, Y. Contributions of two nuclear localization signals of Influenza A virus nucleoprotein to viral replication. Virol. J. 2007, 81, 30–41. [Google Scholar] [CrossRef] [PubMed]
- Sato, Y.; Yoshioka, K.; Suzuki, C.; Awashima, S.; Hosaka, Y.; Yewdell, J.; Kuroda, K. Localization of influenza virus proteins to nuclear dot 10 structures in influenza virus infected cells. Virology 2003, 310, 29–40. [Google Scholar] [CrossRef] [PubMed]
- Simões, M.; Rino, J.; Pinheiro, I.; Martins, C.; Ferreira, F. Alterations of nuclear architecture and epigenetic signatures during African Swine Fever Virus infection. Viruses 2015, 7, 4978–4996. [Google Scholar] [CrossRef]
- Dixon, L.; Chapman, D.; Netherton, C.; Upton, C. African swine fever virus replication and genomics. Virus Res. 2013, 173, 3–14. [Google Scholar] [CrossRef]
- Sánchez, E.; Quintas, A.; Nogal, M.; Castelló, A.; Revilla, Y. African swine fever virus controls the host transcription and cellular machinery of protein synthesis. Virus Res. 2013, 173, 58–75. [Google Scholar] [CrossRef]
- Sperandio, S.; Barat, C.; Cabrita, M.; Gargaun, A.; Berezovski, M.; Tremblay, M.; de Belle, I. TOE1 is an inhibitor of HIV-1 replication with cell-penetrating capability. Proc. Natl. Acad. Sci. USA 2015, 112, E3392–E3401. [Google Scholar] [CrossRef]
- Fanales-Belasio, E.; Raimondo, M.; Suligoi, B.; Buttò, S. HIV virology and pathogenetic mechanisms of infection: A brief overview. Ann. Ist. Super. Sanita. 2010, 46, 5–14. [Google Scholar] [CrossRef]
- Rensen, E.; Mueller, F.; Scoca, V.; Parmar, J.J.; Souque, P.; Zimmer, C.; Di Nunzio, F. Clustering and reverse transcription of HIV-1 genomes in nuclear niches of macrophages. EMBO J. 2021, 40, e105247. [Google Scholar] [CrossRef]
- Cotmore, S.; Sturzenbecker, L.; Tattersall, P. The autonomous parvovirus MVM encodes two nonstructural proteins in addition to its capsid polypeptides. Virology 1983, 129, 333–343. [Google Scholar] [CrossRef]
- Cotmore, S.F.; Tattersall, P. Organization of nonstructural genes of the autonomous Parvovirus Minute Virus of mice. Virol. J. 1986, 58, 724–732. [Google Scholar] [CrossRef]
- Cziepluch, C.; Lampel, S.; Grewenig, A.; Grund, C.; Lichter, P.; Rommelaere, J. H-1 Parvovirus-associated replication bodies: A distinct virus-induced nuclear structure. Virol. J. 2000, 74, 4807–4815. [Google Scholar] [CrossRef]
- Bashir, T.; Rommelaere, J.; Cziepluch, C. In Vivo accumulation of Cyclin A and cellular replication factors in autonomous Parvovirus Minute Virus of mice-associated replication bodies. Virol. J. 2001, 75, 4394–4398. [Google Scholar] [CrossRef] [PubMed]
- Webster, R.G.; Bean, W.J.; Gorman, O.T.; Chambers, T.M.; Kawaoka, Y. Evolution and ecology of Influenza A viruses. Microbiol. Rev. 1992, 56, 152–179. [Google Scholar] [CrossRef] [PubMed]
- Josset, L.; Frobert, E.; Rosa-Calatrava, M. Influenza A replication and host nuclear compartments: Many changes and many questions. J. Clin. Virol. 2008, 43, 381–390. [Google Scholar] [CrossRef] [PubMed]
- Engelhardt, O.G.; Fodor, E. Functional association between viral and cellular transcription during influenza virus infection. Rev. Med. Virol. 2006, 16, 329–345. [Google Scholar] [CrossRef] [PubMed]
- Chelbi-Alix, M.; Quignon, F.; Pelicano, L.; Koken, M.; de Thé, H. Resistance to virus infection conferred by the interferon-induced promyelocytic leukemia protein. Virol. J. 1998, 72, 1043–1051. [Google Scholar] [CrossRef]
- Compans, R.W.; Dimmock, N.J. An electron microscopic study of single-cycle infection of chick embryo fibroblasts by influenza virus. Virology 1969, 39, 499–515. [Google Scholar] [CrossRef] [PubMed]
- Terrier, O.; Moules, V.; Carron, C.; Cartet, G.; Frobert, E.; Yver, M.; Traversier, A.; Wolff, T.; Riteau, B.; Naffakh, N.; et al. The influenza fingerprints: NS1 and M1 proteins contribute to specific host cell ultrastructure signatures upon infection by different influenza A viruses. Virology 2012, 432, 204–218. [Google Scholar] [CrossRef]
- Shankar, A.; Patil, A.; Skariyachan, S. Recent perspectives on genome, transmission, clinical manifestation, diagnosis, therapeutic strategies, vaccine developments, and challenges of Zika Virus research. Front. Microbiol. 2017, 8, 1761. [Google Scholar] [CrossRef] [PubMed]
- Hearing, P. Adenoviridae: The viruses and their replication. In Fields Virology: DNA Viruses, 7th ed.; Howley, P.M., Knipe, D.M., Eds.; Wolters Kluwer Health: Philadelphia, PA, USA, 2021; Volume 2, pp. 97–127. [Google Scholar]
- Guimet, D.; Hearing, P. The adenovirus L4-22K protein has distinct functions in the posttranscriptional regulation of gene expression and encapsidation of the viral genome. Virol. J. 2013, 87, 7688–7699. [Google Scholar] [CrossRef] [PubMed]
- Biasiotto, R.; Akusjärvi, G. Regulation of human adenovirus alternative RNA splicing by the adenoviral L4-33K and L4-22K proteins. Int. J. Mol. Sci. 2015, 16, 2893–2912. [Google Scholar] [CrossRef] [PubMed]
- Ullman, A.J.; Hearing, P. Cellular proteins PML and Daxx mediate an innate antiviral defense antagonized by the adenovirus E4 ORF3 protein. Virol. J. 2008, 82, 7325–7335. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Ruiz, S.; Soler, N.; Sánchez-Navarro, J.; Fagoaga, C.; Lopez, C.; Navarro, L.; Moreno, P.; Peña, L.; Flores, R. Citrus tristeza virus p23: Determinants for nucleolar localization and their influence on suppression of RNA silencing and pathogenesis. Mol. Plant-Microbe Interact. 2013, 26, 306–318. [Google Scholar] [CrossRef]
- Pérez-Cañamás, M.; Hevia, E.; Hernández, C. Epigenetic changes in host ribosomal DNA promoter induced by an asymptomatic plant virus infection. Biology 2020, 9, 91. [Google Scholar] [CrossRef]
- Pérez-Cañamás, M.; Taliansky, M.; Hernández, C. A viral suppressor of RNA silencing may be targeting a plant defence pathway involving fibrillarin. Plants 2022, 11, 1903. [Google Scholar] [CrossRef]
- Rajamaki, M.L.; Valkonen, J.P. Control of nuclear and nucleolar localization of nuclear inclusion protein a of Picorna-Like Potato Virus A in nicotiana species. Plant Cell 2009, 21, 2485–2502. [Google Scholar] [CrossRef]
- Kim, S.; Ryabov, E.; Kalinina, N.; Rakitina, D.; Gillespie, T.; MacFarlane, S.; Haupt, S.; Brown, J.; Taliansky, M. Cajal bodies and the nucleolus are required for a plant virus systemic infection. EMBO J. 2007, 26, 2169–2179. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; MacFarlane, S.; Kalinina, N.; Rakitina, D.; Ryabov, E.; Gillespie, T.; Haupt, S.; Brown, J.; Taliansky, M. Interaction of a plant virus-encoded protein with the major nucleolar protein fibrillarin is required for systemic virus infection. Proc. Natl. Acad. Sci. USA 2007, 104, 11115–11120. [Google Scholar] [CrossRef] [PubMed]
- Guo, T.; Vimalesvaran, D.; Thompson, J.; Perry, K.; Krenz, B. Subcellular localization of grapevine red blotch-associated virus ORFs V2 and V3. Virus Genes 2015, 51, 156–158. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; He, J.; Ding, Z.; Zhang, C.; Meng, R. Identification of functional domain(s) of fibrillarin interacted with p2 of Rice stripe virus. Can. J. Infect. Dis. Med. Microbiol. 2018, 2018, 8402839. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; Hong, P.; Guo, X.; Li, Y.; Xie, L. Rice stripe virus p2 colocalizes and interacts with Arabidopsis Cajal bodies and its domains in plant cells. BioMed Res. Int. 2020, 2020, 5182164. [Google Scholar] [CrossRef] [PubMed]
- Lim, H.; Bragg, J.; Ganesan, U.; Lawrence, D.; Yu, J.; Isogai, M.; Hammond, J.; Jackson, A. triple gene block protein interactions involved in movement of Barley Stripe Mosaic Virus. Virol. J. 2008, 82, 4991–5006. [Google Scholar] [CrossRef]
- Semashko, M.; Rakitina, D.; González, I.; Canto, T.; Kalinina, N.; Taliansky, M. Movement protein of Hordeivirus interacts in vitro and in vivo with coilin, a major structural protein of Cajal bodies. Dokl. Biochem. 2012, 442, 57–60. [Google Scholar] [CrossRef] [PubMed]
- Shaw, J.; Love, A.; Makarova, S.; Kalinina, N.; Harrison, B.; Taliansky, M. Coilin, the signature protein of Cajal bodies, differentially modulates the interactions of plants with viruses in widely different taxa. Nucleus 2014, 5, 85–94. [Google Scholar] [CrossRef]
- Shaw, J.; Yu, C.; Makhotenko, A.; Makarova, S.; Love, A.; Kalinina, N.; MacFarlane, S.; Chen, J.; Taliansky, M. Interaction of a plant virus protein with the signature Cajal body protein coilin facilitates salicylic acid-mediated plant defence responses. New Phytol. 2019, 224, 439–453. [Google Scholar] [CrossRef]
- Li, Z.; Zhang, Y.; Jiang, Z.; Jin, X.; Zhang, K.; Wang, X.; Han, C.; Yu, J.; Li, D. Hijacking of the nucleolar protein fibrillarin by TGB1 is required for cell-to-cell movement of Barley stripe mosaic virus. Mol. Plant Pathol. 2017, 19, 1222–1237. [Google Scholar] [CrossRef]
- Wang, L.; Ding, Y.; He, L.; Zhang, G.; Zhu, J.; Lozano-Duran, R. A virus-encoded protein suppresses methylation of the viral genome through its interaction with AGO4 in the Cajal body. eLife 2020, 9, e55542. [Google Scholar] [CrossRef] [PubMed]
- Hamera, S.; Song, X.; Su, L.; Chen, X.; Fang, R. Cucumber mosaic virus suppressor 2b binds to AGO4-related small RNAs and impairs AGO4 activities. Plant J. 2012, 69, 104–115. [Google Scholar] [CrossRef] [PubMed]
- Duan, C.G.; Fang, Y.Y.; Zhou, B.J.; Zhao, J.H.; Hou, W.N.; Zhu, H.; Ding, S.W.; Guo, H.S. Suppression of Arabidopsis ARGONAUTE1-mediated slicing, transgene-induced RNA silencing, and DNA methylation by distinct domains of the Cucumber mosaic virus 2b protein. Plant Cell 2012, 24, 259–274. [Google Scholar] [CrossRef] [PubMed]
- Schaad, M.; Haldeman-Cahill, R.; Cronin, S.; Carrington, J. Analysis of the VPg-proteinase (NIa) encoded by tobacco etch potyvirus: Effects of mutations on subcellular transport, proteolytic processing, and genome amplification. Virol. J. 1996, 70, 7039–7048. [Google Scholar] [CrossRef] [PubMed]
- Taliansky, M.E.; Robinson, D.J. Molecular biology of umbraviruses: Phantom warriors. J. Gen. Virol. 2003, 84, 1951–1960. [Google Scholar] [CrossRef] [PubMed]
- Ryabov, E.; Oparka, K.; Santa Cruz, S.; Robinson, D.; Taliansky, M. Intracellular location of two Groundnut Rosette Umbravirus proteins delivered by PVX and TMV vectors. Virology 1998, 242, 303–313. [Google Scholar] [CrossRef] [PubMed]
- Ryabov, E.; Robinson, D.; Taliansky, M. A plant virus-encoded protein facilitates long-distance movement of heterologous viral RNA. Proc. Natl. Acad. Sci. USA 1999, 96, 1212–1217. [Google Scholar] [CrossRef] [PubMed]
- Taliansky, M.; Roberts, I.; Kalinina, N.; Ryabov, E.; Raj, S.; Robinson, D.; Oparka, K. An umbraviral protein, involved in long-distance RNA movement, binds viral RNA and forms unique, protective ribonucleoprotein complexes. Virol. J. 2003, 77, 3031–3040. [Google Scholar] [CrossRef]
- Solovyev, A.; Savenkov, E.; Agranovsky, A.; Morozov, S. Comparisons of the genomiccis-elements and coding regions in RNAβ components of the Hordeiviruses Barley stripe mosaic virus, Lychnis ringspot virus, and Poa semilatent virus. Virology 1996, 219, 9–18. [Google Scholar] [CrossRef]
- Jackson, A.; Lim, H.; Bragg, J.; Ganesan, U.; Lee, M. Hordeivirus replication, movement, and pathogenesis. Annu. Rev. Phytopathol. 2009, 47, 385–422. [Google Scholar] [CrossRef]
- MacFarlane, S. Tobraviruses-plant pathogens and tools for biotechnology. Mol. Plant Pathol. 2010, 11, 577–583. [Google Scholar] [CrossRef] [PubMed]
- Spechenkova, N.; Samarskaya, V.O.; Kalinina, N.O.; Zavriev, S.K.; MacFarlane, S.; Love, A.J.; Taliansky, M. Plant Poly (ADP-ribose) polymerase 1 is a potential mediator of cross-talk between the Cajal body protein coilin and salicylic acid-mediated antiviral defence. Viruses 2023, 15, 1282. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Jackson, A.O. Expression of the Barley Stripe Mosaic Virus RNAβ “Triple gene block”. Virology 1996, 216, 367–379. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; Du, Z.; Lin, C.; Mao, Q.; Wu, K.; Wu, J.; Wei, T.; Wu, Z.; Xie, L. Rice stripe tenuivirus p2 may recruit or manipulate nucleolar functions through an interaction with fibrillarin to promote virus systemic movement. Mol. Plant Pathol. 2015, 16, 921–930. [Google Scholar] [CrossRef]
- Zhao, S.; Gong, P.; Ren, Y.; Liu, H.; Li, H.; Li, F.; Zhou, X. The novel C5 protein from tomato yellow leaf curl virus is a virulence factor and suppressor of gene silencing. Stress Biol. 2022, 2, 19. [Google Scholar] [CrossRef]
- Czosnek, H.; Eybishtz, A.; Sade, D.; Gorovits, R.; Sobol, I.; Bejarano, E.; Rosas-Díaz, T.; Lozano-Durán, R. Discovering host genes involved in the infection by the tomato yellow leaf curl virus complex and in the establishment of resistance to the virus using tobacco rattle virus-based post transcriptional gene silencing. Viruses 2013, 5, 998–1022. [Google Scholar] [CrossRef]
- Ceniceros-Ojeda, E.; Rodríguez-Negrete, E.; Rivera-Bustamante, R. Two populations of viral minichromosomes are present in a geminivirus-infected plant showing symptom remission (Recovery). Virol. J. 2016, 90, 3828–3838. [Google Scholar] [CrossRef]
- Deuschle, K.; Kepp, G.; Jeske, H. Differential methylation of the circular DNA in geminiviral minichromosomes. Virology 2016, 499, 243–258. [Google Scholar] [CrossRef]
- Raja, P.; Sanville, B.; Buchmann, R.; Bisaro, D. Viral genome methylation as an epigenetic defense against Geminiviruses. Virol. J. 2008, 82, 8997–9007. [Google Scholar] [CrossRef]
- Rodríguez-Negrete, E.; Carrillo-Tripp, J.; Rivera-Bustamante, R. RNA silencing against Geminivirus: Complementary action of posttranscriptional gene silencing and transcriptional gene silencing in host recovery. Virol. J. 2008, 83, 1332–1340. [Google Scholar] [CrossRef]
- Wang, B.; Li, F.; Huang, C.; Yang, X.; Qian, Y.; Xie, Y.; Zhou, X. V2 of tomato yellow leaf curl virus can suppress methylation-mediated transcriptional gene silencing in plants. J. Gen. Virol. 2014, 95, 225–230. [Google Scholar] [CrossRef] [PubMed]
- Zrachya, A.; Glick, E.; Levy, Y.; Arazi, T.; Citovsky, V.; Gafni, Y. Suppressor of RNA silencing encoded by tomato yellow leaf curl virus-israel. Virology 2007, 358, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Yang, X.; Wang, Y.; Xie, Y.; Zhou, X. Tomato Yellow Leaf Curl Virus V2 interacts with host histone deacetylase 6 to suppress methylation-mediated transcriptional gene silencing in plants. J. Virol. 2018, 92, 00036-18. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wu, Y.; Gong, Q.; Ismayil, A.; Yuan, Y.; Lian, B.; Jia, Q.; Han, M.; Deng, H.; Hong, Y.; et al. Geminiviral V2 protein suppresses transcriptional gene silencing through interaction with AGO4. J. Virol. 2019, 93, e01675-18. [Google Scholar] [CrossRef]
- Lin, J.; Chen, J.; Elenbaas, B.; Levine, A. Several hydrophobic amino acids in the p53 amino-terminal domain are required for transcriptional activation, binding to mdm-2 and the adenovirus 5 E1B 55-kD protein. Genes Dev. 1994, 8, 1235–1246. [Google Scholar] [CrossRef]
- Schreiner, S.; Wimmer, P.; Sirma, H.; Everett, R.; Blanchette, P.; Groitl, P.; Dobner, T. Proteasome-dependent degradation of Daxx by the viral E1B-55K Protein in human adenovirus-infected cells. J. Virol. 2010, 84, 7029–7038. [Google Scholar] [CrossRef]
- Zhao, L.; Colosimo, A.; Liu, Y.; Wan, Y.; Liao, D. Adenovirus E1B 55-kilodalton oncoprotein binds to Daxx and eliminates enhancement of p53-dependent transcription by Daxx. J. Virol. 2003, 77, 11809–11821. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Species Infected by Virus | Name of Virus | Virus Protein/Nucleic Acid Sequence Element That Interacts with Cajal Bodies | Cajal Body Protein That Interacts with Virus | Function of Interaction | Other Nuclear Bodies That Interact with Virus (Along with CBs) | References |
|---|---|---|---|---|---|---|
| Aves (Birds), principally Gallus gallus domesticus (Chicken) | Marek’s Disease Virus (MDV) (Avian alphaherpesvirus) | Meq/Meq/vIL8 | p80-Coilin | Unknown | Nucleoplasm, nucleolus | [92,93] |
| Several mammalian species, including Homo sapiens (Human) | Adenovirus Type 5 (Ad5) | L4-22K | p80-coilin | Ad5 infection disrupts CBs, redistributed into CB microfoci; L4-22K forms complex with p80-coilin. Depletion of p80-coilin reduces export of virus mRNAs from nucleus in infected cells | PML bodies (E4-ORF3 11 kDa), PML bodies (E1A) | [94,95,96,97,98] |
| Several animal species, including Homo sapiens (Human) | Zika Virus (ZIKV) | NS5 | p80-coilin | Unknown, disrupt CBs, resulting in an increased number of smaller CBs | Unknown | [99] |
| Murinae (Murine), principally Mus musculus (Mouse) | Minute Virus of Mice (MVM) | NS1 | SMN | Disrupt components of CBs, recombined into SAABs (sites of virus replication) | Unknown | [100] |
| Homo sapiens (Human) | Human Cytomegalovirus (HCMV) | UL3 and UL30 | Unknown | Unknown, expression of UL3 and UL30 decreased number of CBs in cell | PML bodies | [101] |
| Several animal species, including Homo sapiens (Human) | Influenza A virus | NP | Unknown | Unknown, expression of NP results in an increased number of smaller CBs | PML bodies, nucleolus (virus NS1) | [102,103,104,105,106,107] |
| Sus scrofa domesticus (Swine) | African Swine Fever Virus (AFSV) | Unknown | Unknown | Unknown, disrupt CBs, results in an increased number of smaller CBs | Nuclear speckles, PML bodies | [108,109,110] |
| Homo sapiens (Human) | Human Immunodeficiency Virus (HIV) | TAR (Transactivation Response RNA Element) | TOE1 | Host antivirus defence | Unknown | [111,112,113] |
| Species Infected by Virus | Name of Virus | Virus Protein That Interacts with Cajal Bodies | Cajal Body Protein That Interacts with Virus | Function of Interaction | Other Nuclear Bodies That Interact with Virus (Along with CBs) | References |
|---|---|---|---|---|---|---|
| Nicotiana benthamiana | Citrus tristeza virus (CTV) | P23 | Fibrillarin | Suppression of RNA silencing, enhancing systemic infection and virus accumulation | Nucleolus | [129] |
| Nicotiana benthamiana | Pelargonium line pattern virus (PLPV) | P37 | Fibrillarin, coilin | Suppression of RNA silencing, overcoming host defence, modulation of methylation of ribosomal DNA promoter | Nucleolus | [130,131] |
| Solanum tuberosum (Potato) | Potato Virus A (PVA) | NIa | Fibrillarin | Unknown, reduced fibrillarin resulted in reduced accumulation of PVA | Nucleolus | [132] |
| Arachis hypogaea (Groundnut) | Groundnut Rosette Virus (GRV) | ORF3 | Fibrillarin | Long-distance movement, establishing systemic infection, stabilisation of virus RNA | Nucleolus | [133,134] |
| Vitis (Grapevine) | Grapevine Red Blotch-associated Virus (GrRBaV) | V2 | Fibrillarin, coilin | Unknown | Nucleolus, inclusions in the nucleoplasm | [135] |
| Oryza sativa (Rice) | Rice Stripe Virus (RSV) | P2 | Coilin | Unknown | Nucleolus (via fibrillarin to establish systemic infection) | [136,137] |
| Poaceae (Grass) | Poa Semilatent Virus (PSLV) | TGBp1 | Coilin | Cell-to-cell movement, long distance transport, establishing a systemic infection | Nucleolus | [138,139] |
| A wide variety of species | Tobacco Rattle Virus (TRV) | 16 K | Coilin | Host antivirus defence (RNA and SA silencing pathways) | Nucleolus | [140,141] |
| Hordeum vulgare (Barley) | Barley Stripe Mosaic Virus (BSMV) | TGB1 | Fibrillarin, coilin | Cell-to-cell movement of virus during infection | Unknown | [142] |
| Solanum ycopersicum (Tomato) | Tomato Yellow Leaf Curl Virus (TYLCV) | V2 | AGO4 | Suppression of host antivirus defence (inhibit TGS and PTGS pathways) | Nucleoplasm | [143] |
| A wide variety of species | Cucumber Mosaic Virus (CMV) | 2b | AGO4 | RNA-directed DNA methylation | Nucleolus | [144,145] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lettin, L.; Erbay, B.; Blair, G.E. Viruses and Cajal Bodies: A Critical Cellular Target in Virus Infection? Viruses 2023, 15, 2311. https://doi.org/10.3390/v15122311
Lettin L, Erbay B, Blair GE. Viruses and Cajal Bodies: A Critical Cellular Target in Virus Infection? Viruses. 2023; 15(12):2311. https://doi.org/10.3390/v15122311
Chicago/Turabian StyleLettin, Lucy, Bilgi Erbay, and G. Eric Blair. 2023. "Viruses and Cajal Bodies: A Critical Cellular Target in Virus Infection?" Viruses 15, no. 12: 2311. https://doi.org/10.3390/v15122311