
(Re-)Emergence of Oropouche Virus (OROV) Infections: Systematic Review and Meta-Analysis of Observational Studies

 ,
,  ,
,  , ,
, ,  ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Research Concept
2.2. Study Selection and Inclusion and Exclusion Criteria
- (1)
- Availability of the full text.
- (2)
- The selection criteria for tested participants were preventively reported; for the aims of the present review, both studies on the whole of the resident population from a certain area and studies focusing on patients with potential signs and symptoms were considered.
- (3)
- OROV infection was assessed by means of (a) detection of OROV and/or its RNA in blood, saliva, and/or cerebrospinal fluid (CSF) or central nervous system (CNS) tissue; (b) detection of OROV antigens by immunohistochemical analysis with virus-specific monoclonal antibodies; (c) detection of OROV-reactive IgG and/or IgM in a serum or CSF sample with and without confirmation assay detecting antibodies against individual OROV antigens.
- (1)
- Derivative studies (i.e., systematic reviews and meta-analyses), letters, editorial comments, and case reports;
- (2)
- Studies on animals (including non-human primates);
- (3)
- The full text was not available either through online repositories or through inter-library loans, or its main text was written in a language different from English, Italian, German, French, Spanish, or Portuguese;
- (4)
- Lack of details about geographical setting and corresponding timeframe.
2.3. Data Extraction
- (a)
- Settings of the cases (year, month, or season and geographic region);
- (b)
- Recruitment strategy, summarized as a dichotomous variable: asymptomatic vs. symptomatic subjects;
- (c)
- Laboratory testing strategy, summarized as virological studies (i.e., all laboratory testing leading to the isolation of OROV or its RNA through viral cultures and/or real-time quantitative polymerase chain reaction studies), antigenic studies (i.e., all studies leading to the identification of OROV antigens), and serological studies (i.e., all studies leading to the identification OROV-reactive antibodies).
- (d)
- Where available, data on the testing of other arboviruses (i.e., DENV, CHKV, and ZIKV) were similarly collected.
2.4. Qualitative Assessment
2.5. Data Analysis
2.5.1. Descriptive Analysis
2.5.2. Meta-Analysis
2.5.3. Heterogeneity
2.5.4. Sensitivity Analysis
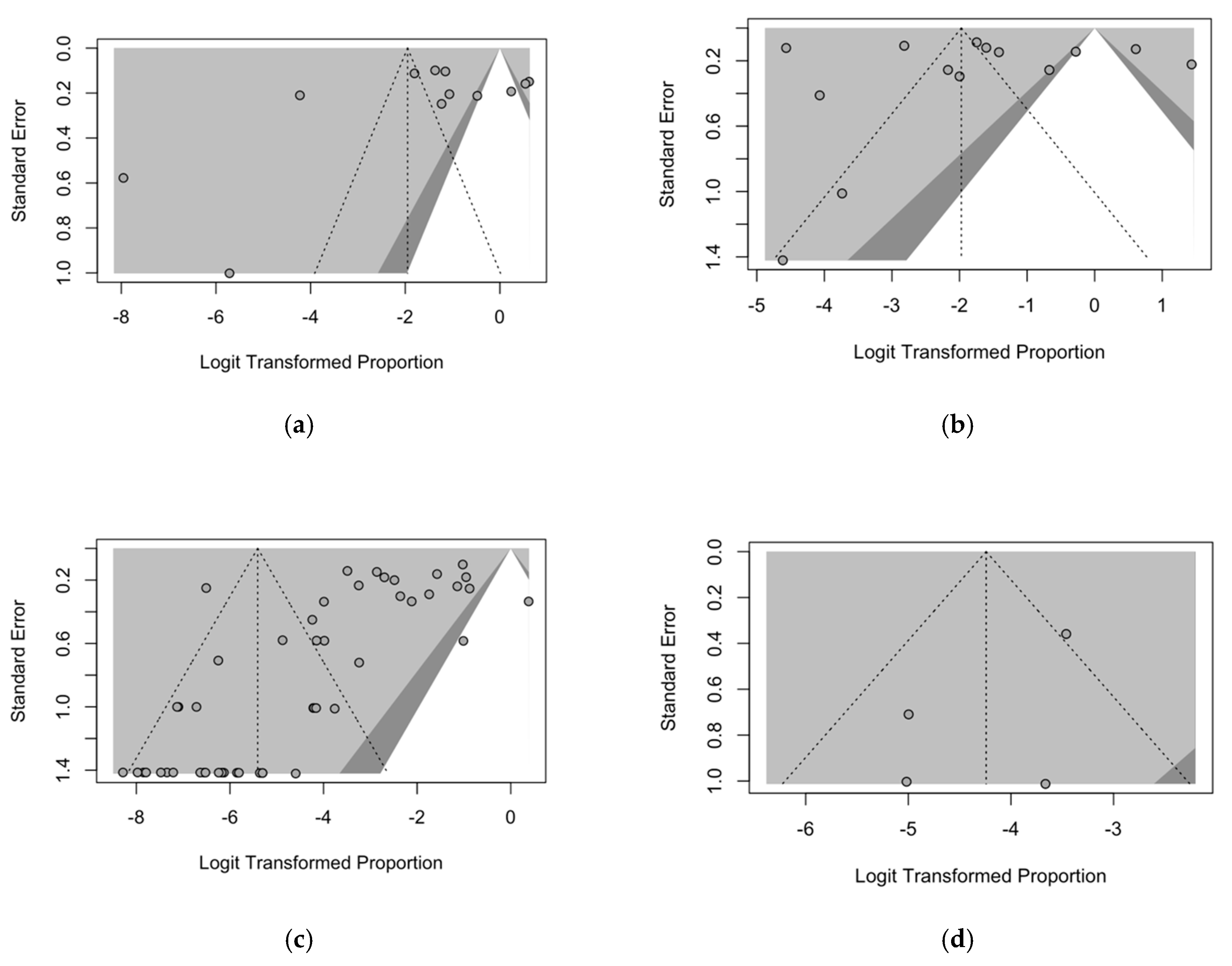
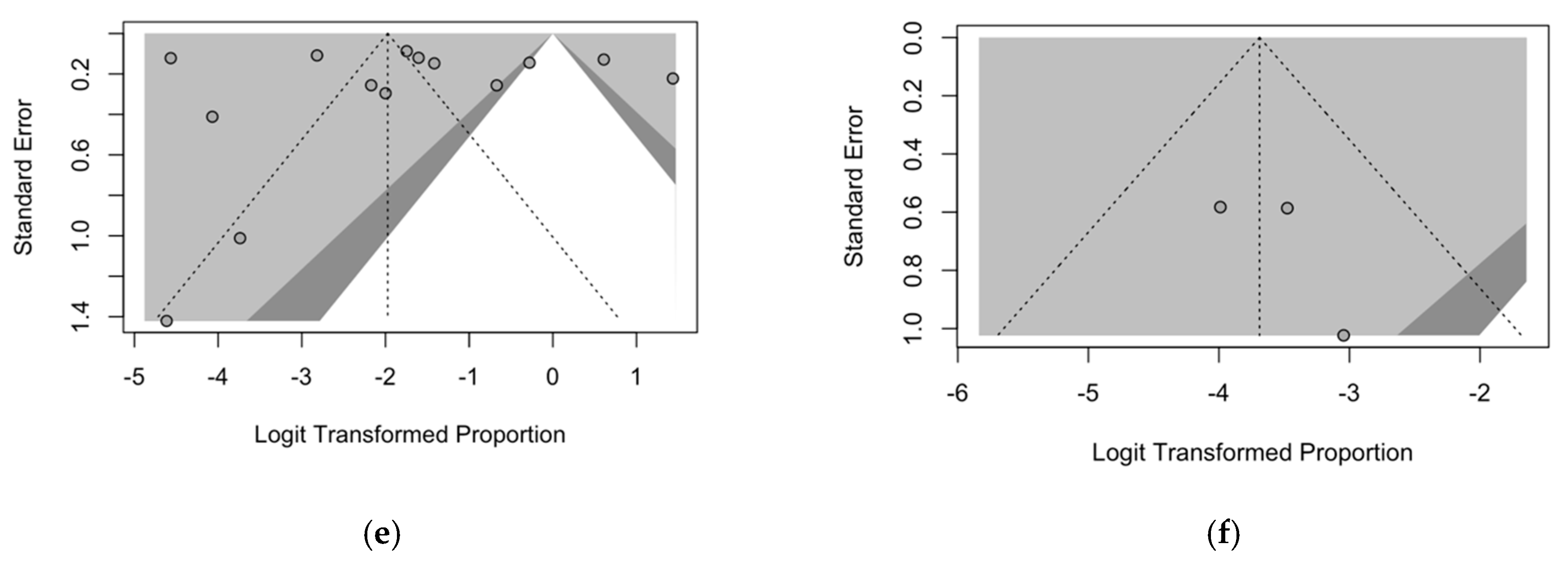
2.5.5. Publication Bias
2.5.6. Software
3. Results
3.1. Search and Selection Process
3.2. Summary of Included Studies
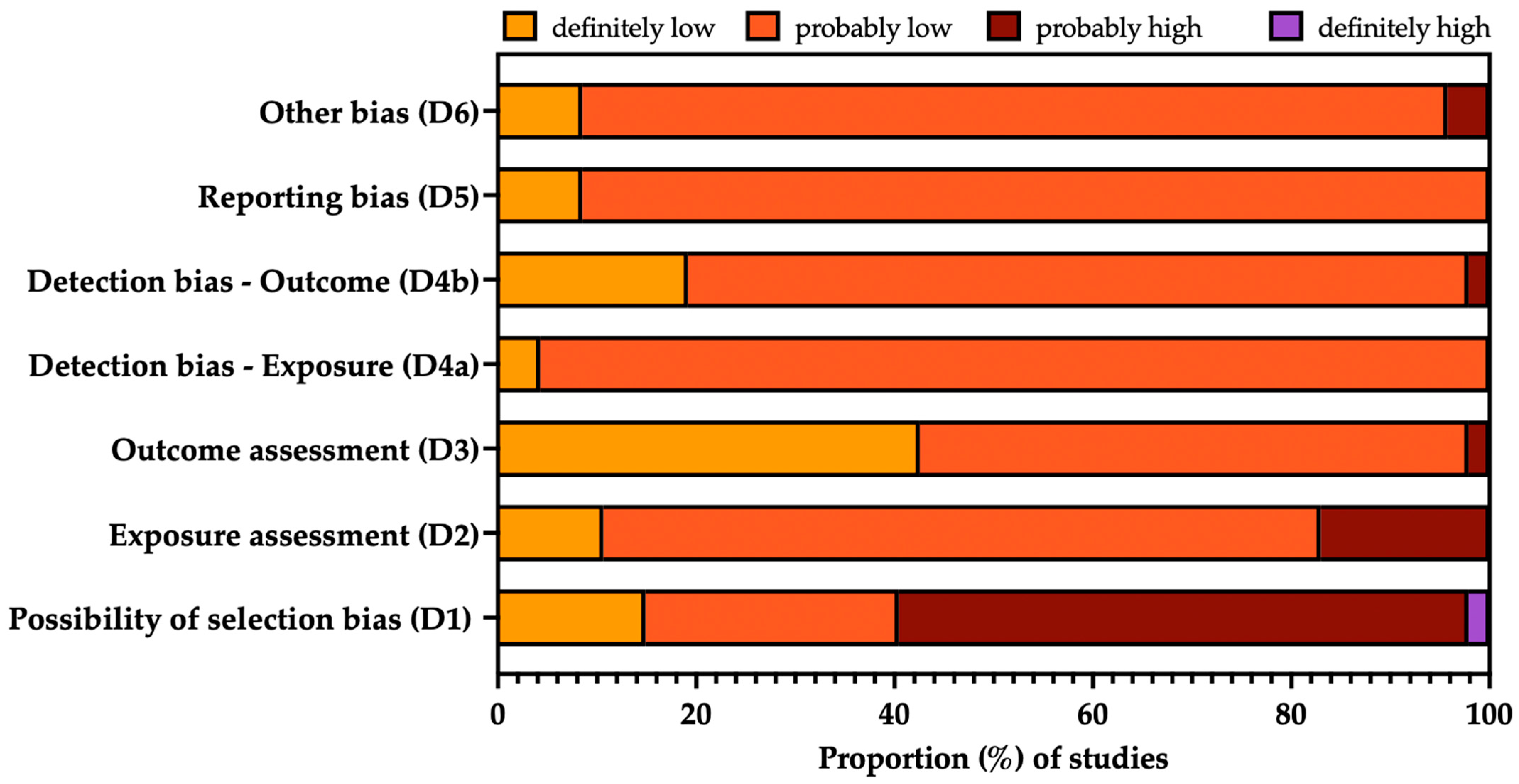
3.3. Risk of Bias Analysis
3.4. Summary of Main Results
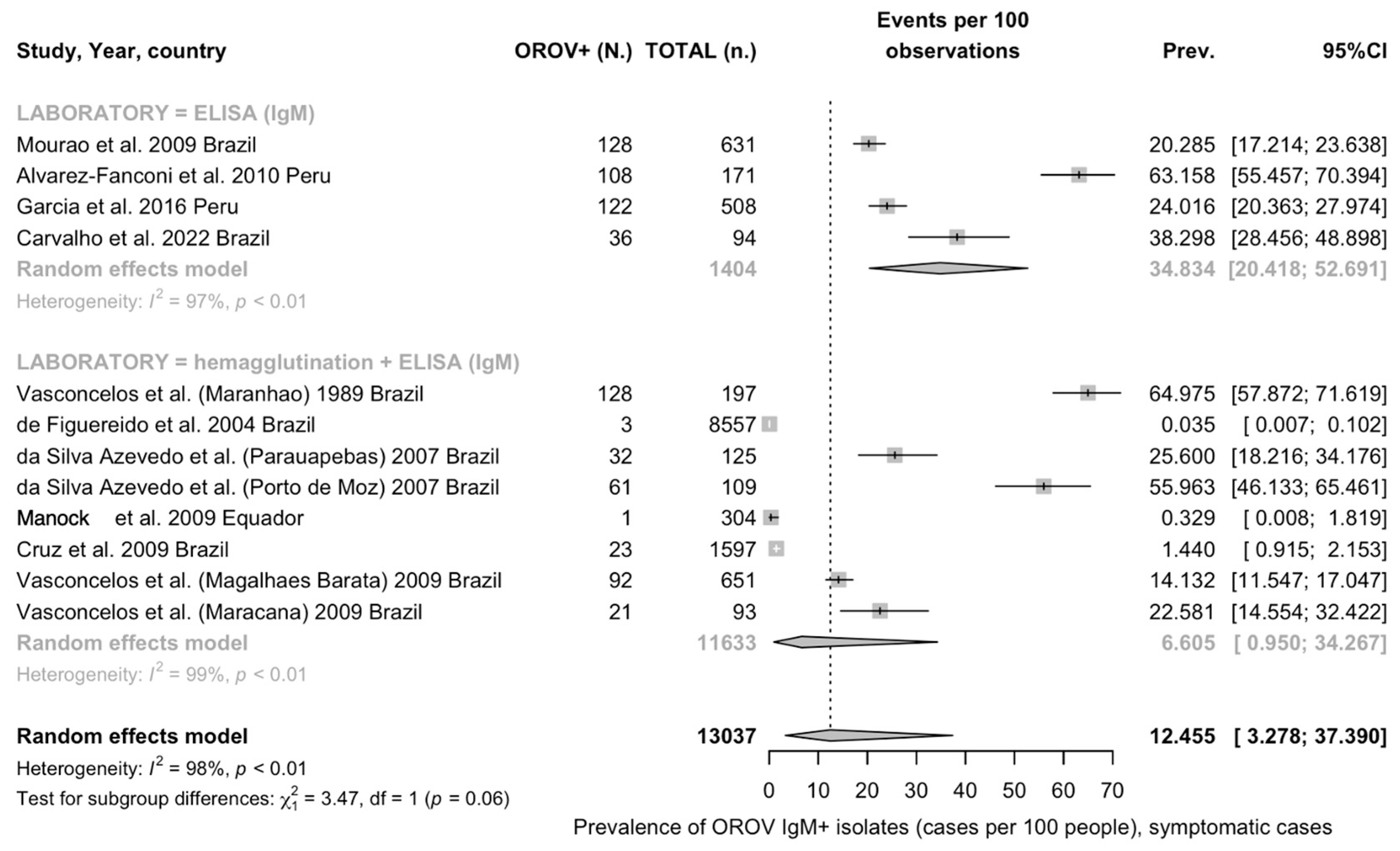
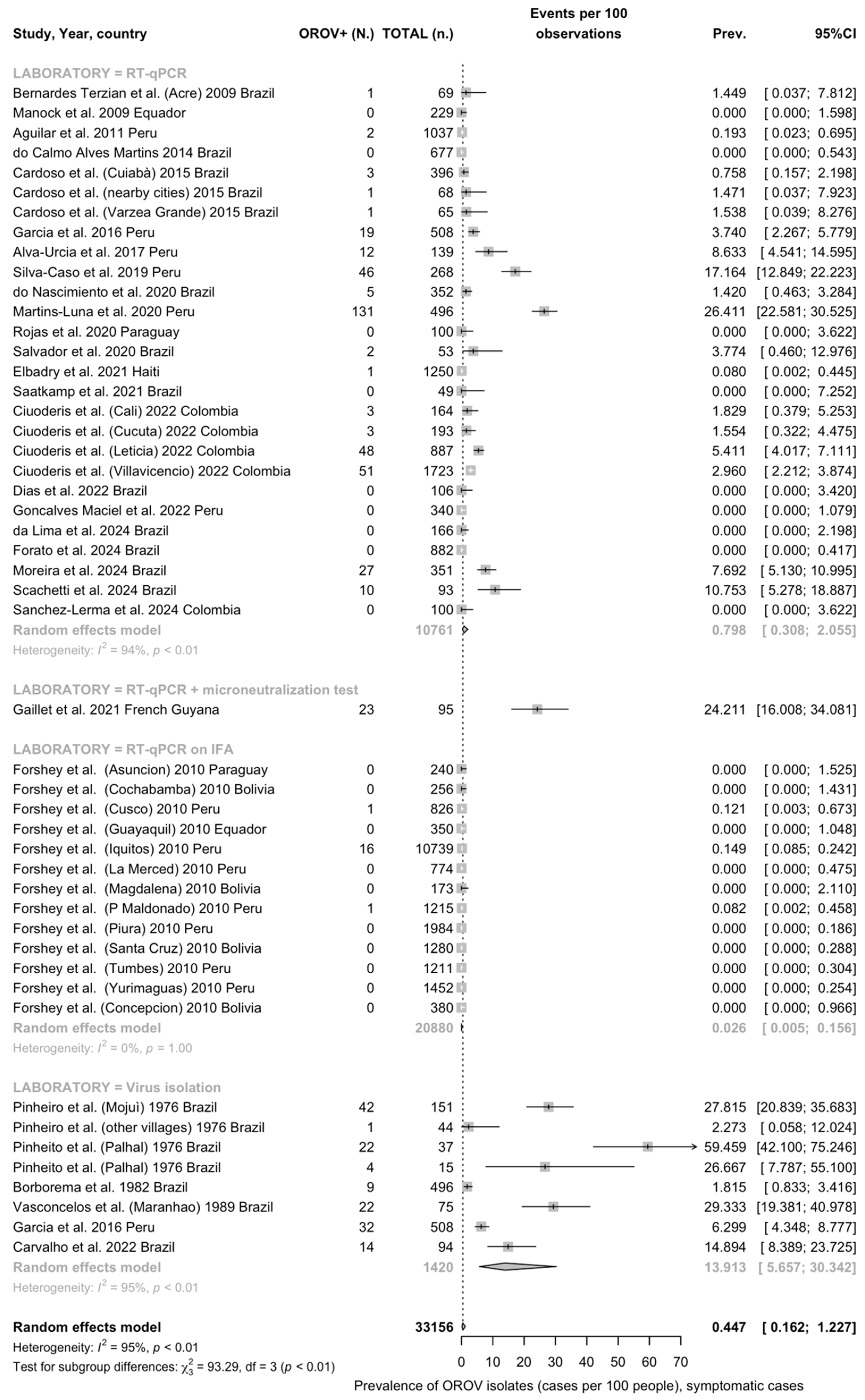
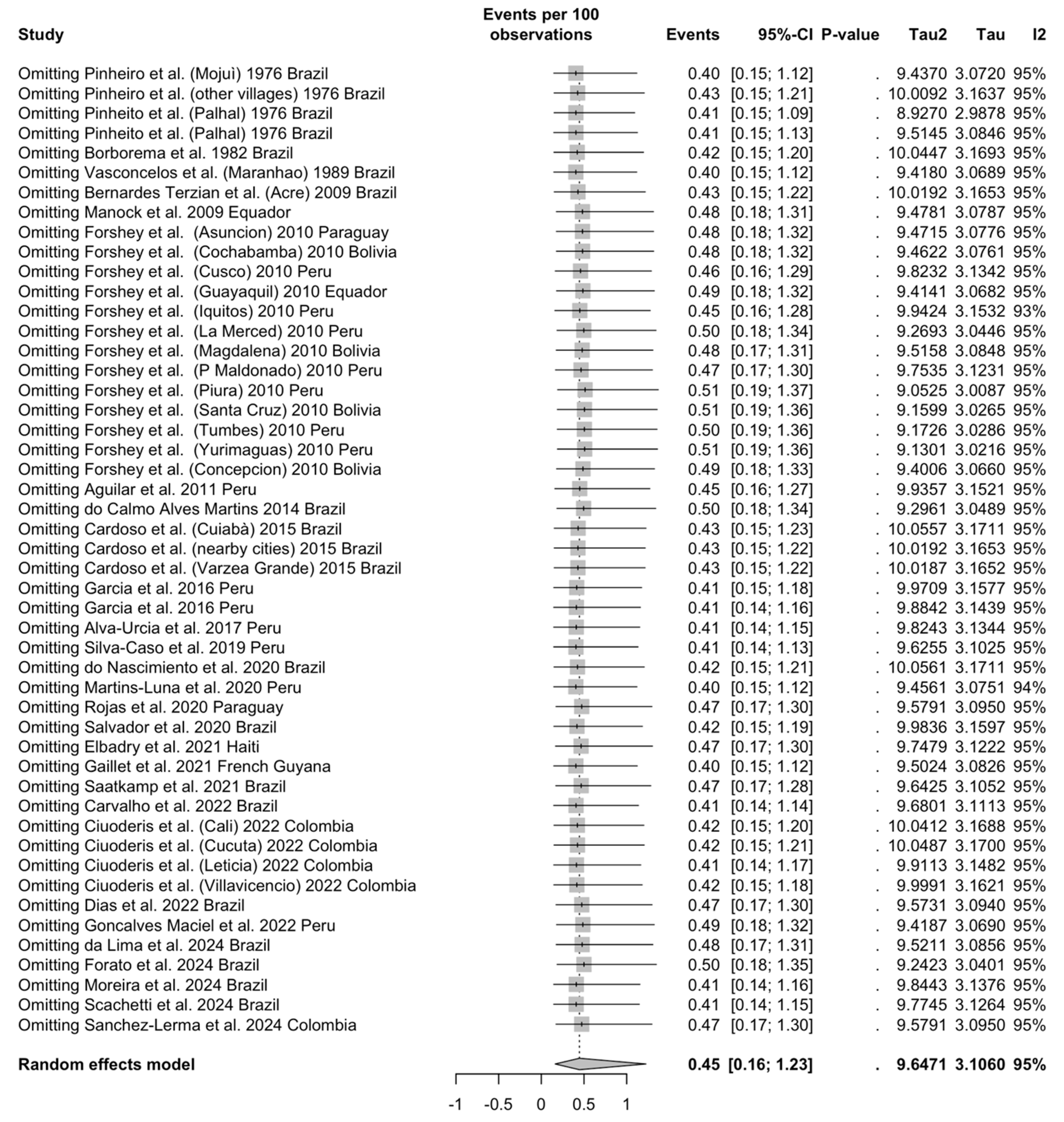
3.4.1. Crude Prevalence Estimates
OROV Identification
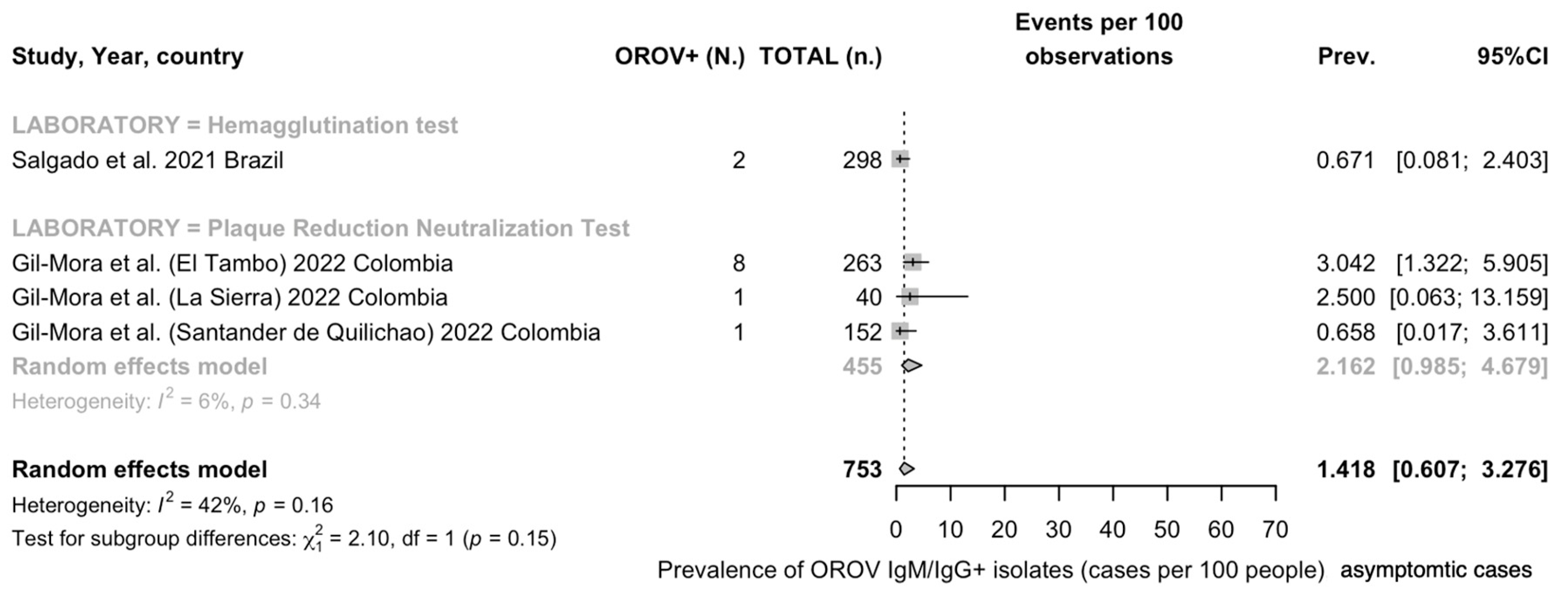
Serology of OROV Infection
3.4.2. Meta-Analysis of Prevalence Estimates
Detection Rates among Individuals with Signs/Symptoms of Arbovirus Infection
Detection Rates in General Population
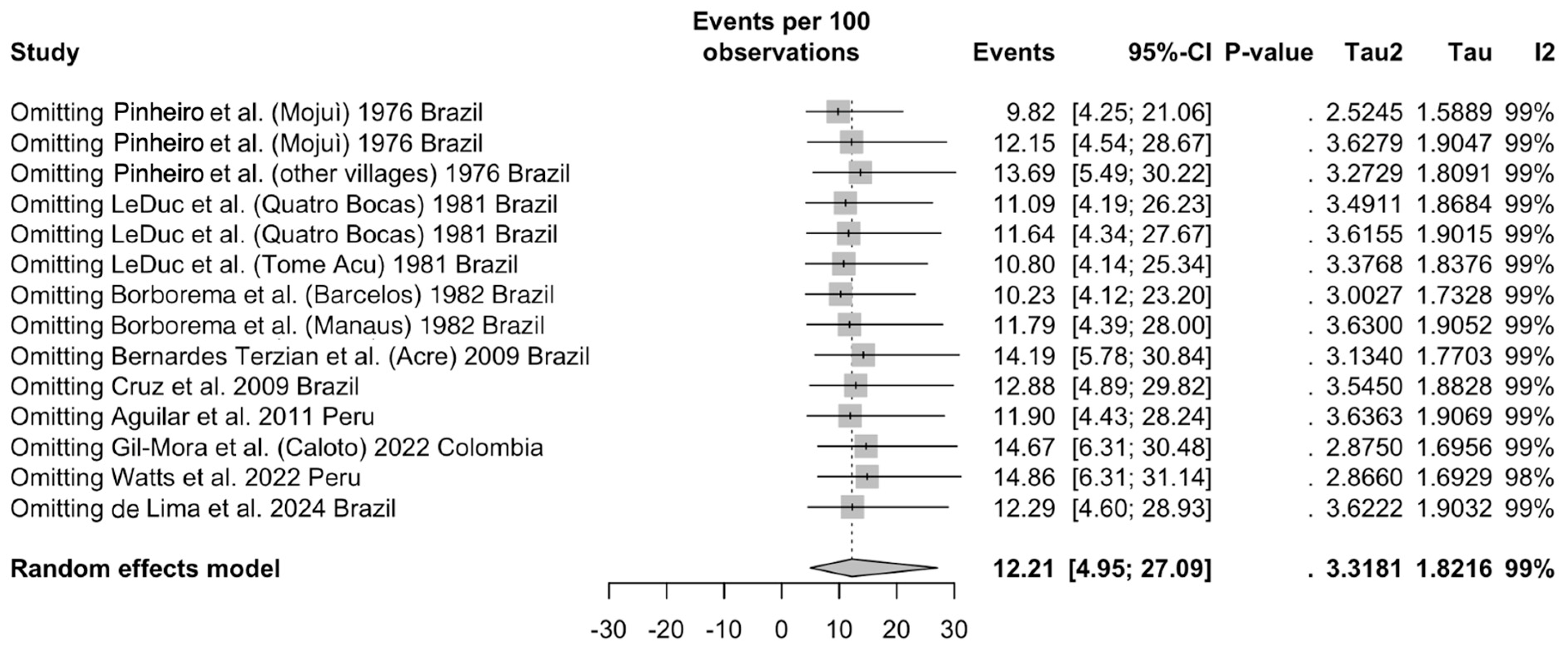
3.5. Sensitivity Analysis
3.5.1. Studies on Individuals Affected by Any Sign or Symptom of Arboviral Infection, Seroprevalence Studies
3.5.2. Studies on Individuals Affected by Any Sign or Symptom of Arboviral Infection, OROV Detection
3.5.3. Studies on General Population— Seroprevalence Estimates
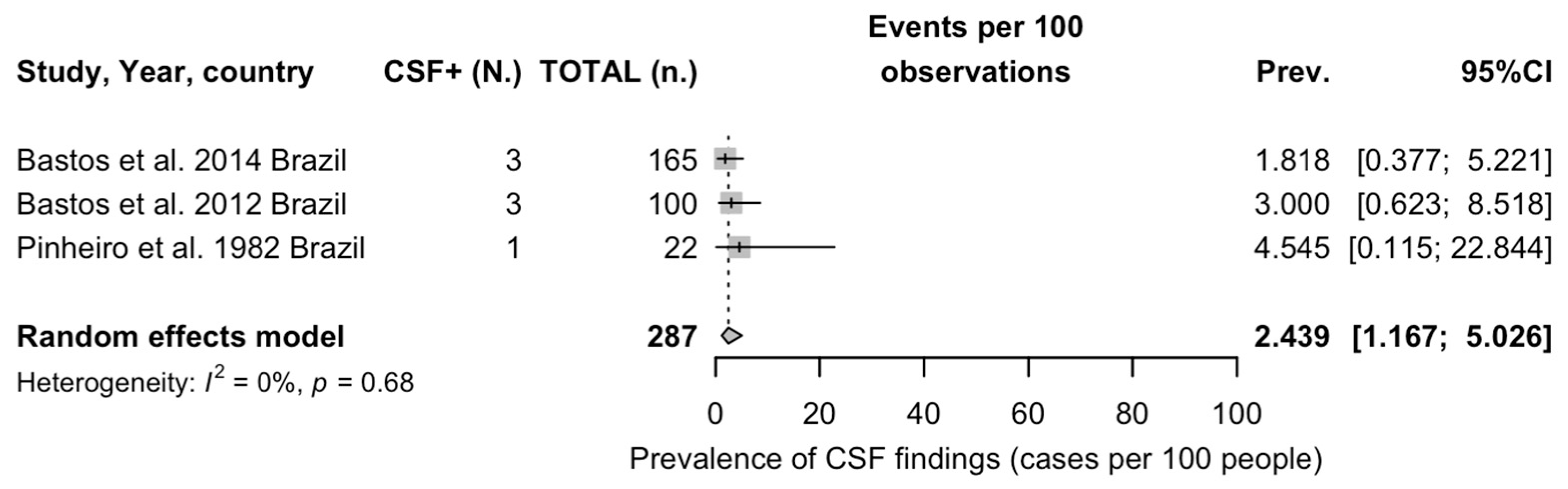
3.5.4. Studies on Detection of OROV in CNS
3.6. Publication Bias
4. Discussion
4.1. Synthesis of Main Findings
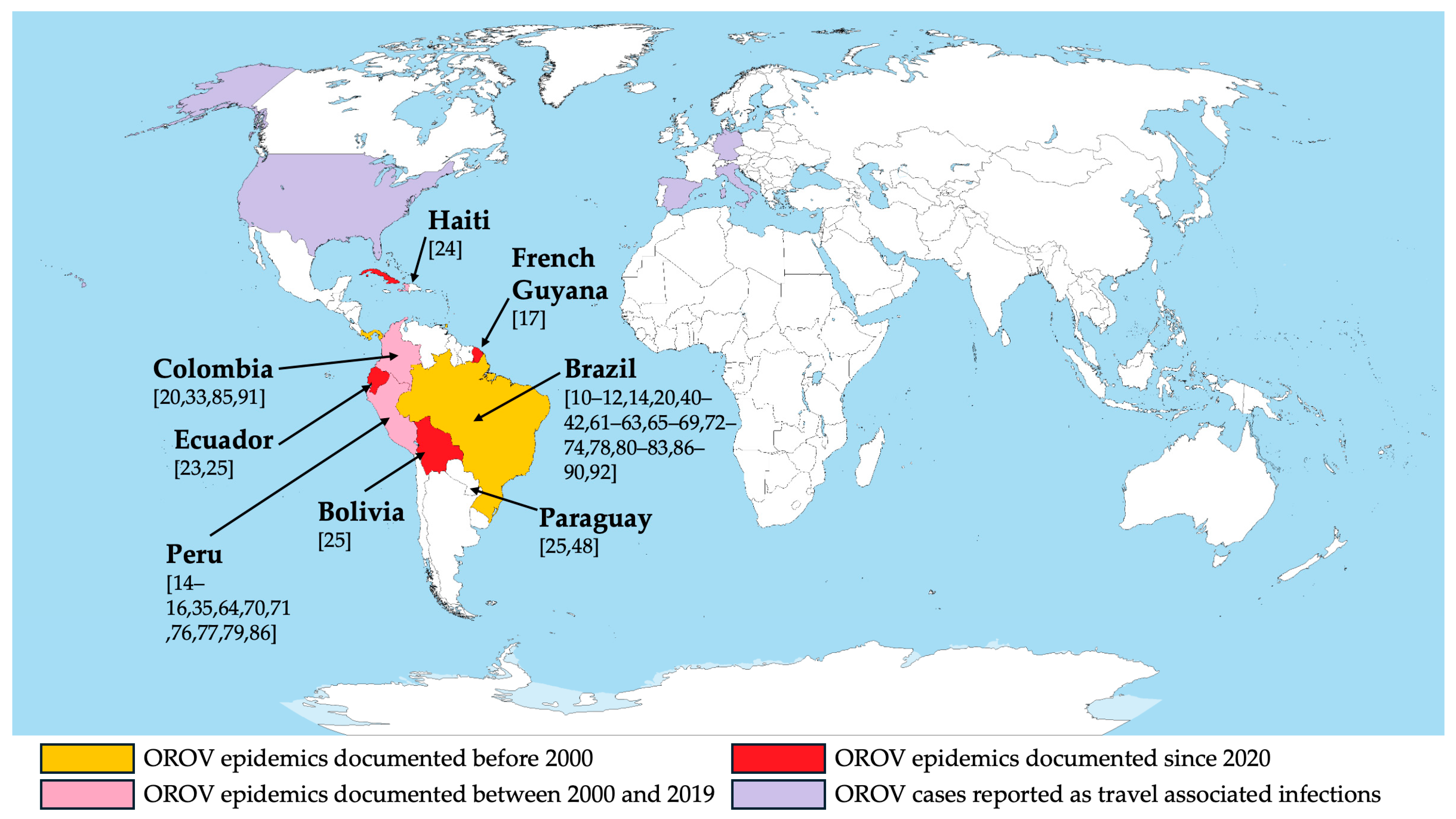
4.2. Generalizability
4.3. Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

| Database | Keywords Searched | No. of Entries Found |
|---|---|---|
| Pubmed | “Oropouche orthobunyavirus” [Supplementary Concept] OR “Oropouche fever” OR “Oropouche virus” OR “oropouche” OR “OROV” | 239 |
| EMBASE | ‘Oropouche orthobunyavirus’ OR ‘Oropouche virus’ OR ‘oropouche fever’ | 215 |
| SCOPUS | “Oropouche orthobunyavirus” OR “Oropouche virus” OR “oropouche fever” OR “oropouche” | 280 |
| MedRxiv | “Oropouche orthobunyavirus” OR “Oropouche fever” OR “Oropouche virus” OR “oropouche” OR “OROV” | 11 |
| Study | D1. Selection | D2. Confounding | D3. Attrition/Exclusion | D4. Detection | D5. Selective Reporting | D6. Other | |
|---|---|---|---|---|---|---|---|
| Did selection of study participants result in appropriate comparison groups? | Did the study design or analysis account for important confounding and modifying variables? | Were outcome data complete with respect to attrition or exclusion from analysis? | Can we be confident in the exposure characterization? | Can we be confident in the outcome assessment? | Were all measured outcomes reported? | Were there any other potential threats to internal validity? | |
| Pinheiro et al., 1976 [10] | ☺ | ☺ | ☺ | ☺☺ | ☺☺ | ☺ | ☺ |
| LeDuc et al., 1981 [11] | ☺ | ☺ | ☺ | ☺☺ | ☺☺ | ☺ | ☺ |
| Pinheiro et al., 1982 [61] | ☹ | ☺ | ☺☺ | ☺ | ☺☺ | ☺☺ | ☺☺ |
| Borborema et al., 1982 [12] | ☺ | ☺ | ☺☺ | ☺ | ☺☺ | ☺☺ | ☺☺ |
| Vasconcelos et al., 1989 [62] | ☺ | ☺ | ☺☺ | ☺ | ☺☺ | ☺☺ | ☺☺ |
| Rosa et al., 1996 [63] | ☹ | ☺ | ☺☺ | ☺ | ☺ | ☺ | ☺ |
| Watts et al., 1997 [64] | ☹ | ☺ | ☺☺ | ☺ | ☺ | ☺ | ☺ |
| Baisley et al., 1998 [14] | ☹ | ☺ | ☺☺ | ☺ | ☺ | ☺ | ☺ |
| de Figueredo et al., 2004 [65] | ☹ | ☺ | ☺☺ | ☺ | ☺ | ☺ | ☺ |
| Tavares Neto et al., 2004 [66] | ☺ | ☺ | ☺☺ | ☺ | ☺ | ☺ | ☺ |
| da Silva Azevedo et al., 2007 [40] | ☹ | ☺ | ☺☺ | ☺ | ☺ | ☺ | ☺ |
| Bernarders Terzian et al., 2009 [67] | ☹ | ☹ | ☺☺ | ☺ | ☺ | ☺ | ☹ |
| Cruz et al., 2009 [68] | ☹ | ☺ | ☺☺ | ☺ | ☺ | ☺ | ☺ |
| Manock et al., 2009 [23] | ☹ | ☺ | ☺☺ | ☺ | ☺ | ☺ | ☺ |
| Mourao et al., 2009 [69] | ☺ | ☺ | ☺ | ☺ | ☺ | ☺ | ☹ |
| Vasconcelos et al., 2009 [41] | ☺ | ☺ | ☺☺ | ☺ | ☺ | ☺ | ☺ |
| AlvarezKFalconi et al., 2010 [70] | ☺ | ☺ | ☺ | ☺ | ☺ | ☺ | ☺ |
| Forshey et al., 2010 [25] | ☺ | ☺ | ☺ | ☺ | ☺ | ☺ | ☺ |
| Aguilar et al., 2011 [71] | ☺☺ | ☺ | ☺☺ | ☺ | ☺ | ☺ | ☺ |
| Bastos et al., 2012 [72] | ☺ | ☺ | ☹ | ☺ | ☺ | ☺ | ☺ |
| Bastos et al., 2014 [73] | ☹ | ☺ | ☺☺ | ☺ | ☹ | ☺ | ☺ |
| do Calmo Martins et al., 2014 [74] | ☹ | ☺ | ☺☺ | ☺ | ☺ | ☺ | ☺ |
| Cardoso et al., 2015 [75] | ☹ | ☺ | ☺ | ☺ | ☺ | ☺ | ☺ |
| Garcia et al., 2016 [76] | ☺☺ | ☺ | ☺☺ | ☺ | ☺ | ☺ | ☺ |
| Alva-Urcia et al., 2017 [77] | ☹ | ☺ | ☺ | ☺ | ☺ | ☺ | ☺ |
| SilvaKCaso et al., 2019 [35] | ☺ | ☺ | ☺ | ☺☺ | ☺☺ | ☺ | ☺ |
| do Nascimiento et al., 2020 [78] | ☺ | ☺ | ☺ | ☺☺ | ☺☺ | ☺ | ☺ |
| MartinsKLuna et al., 2020 [79] | ☹ | ☺ | ☺☺ | ☺ | ☺☺ | ☺☺ | ☺☺ |
| Rojas et al., 2020 [48] | ☺ | ☺ | ☺☺ | ☺ | ☺☺ | ☺☺ | ☺☺ |
| Salvador et al., 2020 [80] | ☺ | ☺ | ☺☺ | ☺ | ☺☺ | ☺☺ | ☺☺ |
| Elbadry et al., 2021 [24] | ☹ | ☺ | ☺☺ | ☺ | ☺ | ☺ | ☺ |
| Gaillet et al., 2021 [17] | ☹ | ☺ | ☺☺ | ☺ | ☺ | ☺ | ☺ |
| Saatkamp et al., 2021 [81] | ☹ | ☺ | ☺☺ | ☺ | ☺ | ☺ | ☺ |
| Salgado et al., 2021 [82] | ☹ | ☺ | ☺☺ | ☺ | ☺ | ☺ | ☺ |
| Carvalho et al., 2022 [83] | ☺ | ☺ | ☺☺ | ☺ | ☺ | ☺ | ☺ |
| Ciuoderis et al., 2022 [33] | ☹ | ☺ | ☺☺ | ☺ | ☺ | ☺ | ☺ |
| Dias et al., 2022 [84] | ☹ | ☹ | ☺☺ | ☺ | ☺ | ☺ | ☹ |
| Gil-Mora et al., 2022 [85] | ☹ | ☺ | ☺☺ | ☺ | ☺ | ☺ | ☺ |
| Gonçalves Maciel et al., 2022 [86] | ☹ | ☺ | ☺☺ | ☺ | ☺ | ☺ | ☺ |
| Watts et al., 2022 [16] | ☺ | ☺ | ☺ | ☺ | ☺ | ☺ | ☹ |
| De Lima et al., 2024 [87] | ☺ | ☺ | ☺☺ | ☺ | ☺ | ☺ | ☺ |
| Forato et al., 2024 [42] | ☺ | ☺ | ☺ | ☺ | ☺ | ☺ | ☺ |
| GrisalesKNieto et al., 2024 [20] | ☺ | ☺ | ☺ | ☺ | ☺ | ☺ | ☺ |
| Moreira et al., 2024 [88] | ☺☺ | ☺ | ☺☺ | ☺ | ☺ | ☺ | ☺ |
| Scachetti et al., 2024 [89] | ☺ | ☺☺ | ☺☺ | ☺ | ☺☺ | ☺ | ☺ |
| Silva et al., 2024 [90] | ☺☺ | ☺ | ☺☺ | ☺ | ☺ | ☺ | ☺ |
| Sanchez-Lerma et al., 2024 | ☹☹ | ☺ | ☺ | ☺ | ☺ | ☺ | ☺ |




References
- Wesselmann, K.M.; Postigo-Hidalgo, I.; Pezzi, L.; de Oliveira-Filho, E.F.; Fischer, C.; de Lamballerie, X.; Drexler, J.F. Emergence of Oropouche Fever in Latin America: A Narrative Review. Lancet Infect. Dis. 2024, 24, e439–e452. [Google Scholar] [CrossRef] [PubMed]
- Sakkas, H.; Bozidis, P.; Franks, A.; Papadopoulou, C. Oropouche Fever: A Review. Viruses 2018, 10, 175. [Google Scholar] [CrossRef] [PubMed]
- Pereira, R.S.; Colangelo, J.F.; Souza, P.G.A.; de Carvalho, L.G.F.; da Cruz Nizer, W.S.; Lima, W.G. Epidemiological Aspects of the Oropouche Virus (Orthobunyavirus) in South America: A Systematic Review. Rev. Colomb. Cienc. Quim. Farm. 2022, 51, 166–184. [Google Scholar] [CrossRef]
- Tilston-Lunel, N.L.; Hughes, J.; Acrani, G.O.; da Silva, D.E.A.; Azevedo, R.S.S.; Rodrigues, S.G.; Vasconcelos, P.F.C.; Nunes, M.R.T.; Elliott, R.M. Genetic Analysis of Members of the Species Oropouche Virus and Identification of a Novel M Segment Sequence. J. Gen. Virol. 2015, 96, 1636–1650. [Google Scholar] [CrossRef]
- Sah, R.; Srivastava, S.; Kumar, S.; Golmei, P.; Rahaman, S.A.; Mehta, R.; Ferraz, C.; Apostolopoulos, V.; Rodriguez-Morales, A.J. Oropouche Fever Outbreak in Brazil: An Emerging Concern in Latin America. Lancet Microbe 2024, 404, 100904. [Google Scholar] [CrossRef]
- Martins-Filho, P.R.; Soares-Neto, R.F.; de Oliveira-Junior, J.M.; Alves dos Santos, C. The Underdiagnosed Threat of Oropouche Fever amidst Dengue in Brazil. Lancet Reg. Health Am. 2024, 32, 100718. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, X.; Wu, Z.; Feng, S.; Lu, K.; Zhu, W.; Sun, H.; Niu, G. Oropouche Virus: A Neglected Global Arboviral Threat. Virus Res 2024, 341, 199318. [Google Scholar] [CrossRef]
- Files, M.A.; Hansen, C.A.; Herrera, V.C.; Schindewolf, C.; Barrett, A.D.T.; Beasley, D.W.C.; Bourne, N.; Milligan, G.N. Baseline Mapping of Oropouche Virology, Epidemiology, Therapeutics, and Vaccine Research and Development. NPJ Vaccines 2022, 7, 38. [Google Scholar] [CrossRef]
- Anderson, C.R.; Spence, L.; Downs, W.G.; Aitken, T.H.G. Oropouche Virus: A New Human Disease Agent from Trinidad, West Indie. Am. J. Trop. Med. Hyg. 1961, 10, 574–578. [Google Scholar] [CrossRef]
- Pinheiro, F.P.; Travassos da Rosa, A.P.; Travassos da Rosa, J.F.; Bensabath, G. An Outbreak of Oropouche Virus Disease in the Vicinity of Santarem, Para, Brazil. Tropenmed Parasitol. 1976, 27, 213–223. [Google Scholar]
- Leduc, J.W.; Hoch, A.L.; Pinheiro, F.P.; Travassos Da Rosa, A.P.A. Epidemic Oropouche Virus Disease in Northern Brazil. Bull. Pan. Am. Hdh. Orp. 1981, 15, 97–103. [Google Scholar]
- Borborema, C.A.T.; Pinheiro, F.P.; Albuqiierque, B.C.; da Rosa, A.P.; Da Rosa, J.F.; Dourado, H.V. Primeiro Registro de Epidemias Causadas Pelo Virus Oropouche No Estado Do Amazonas. Rev. Inst. Med. Trop. São. Paulo. 1982, 24, 132–139. [Google Scholar]
- Baisley, K.J.; Watts, D.M.; Munstermann, L.E.; Wilson, M.L. Epidemiology of Endemic Oropouche Virus Transmission in Upper Amazonian Peru. Am. J. Trop. Med. Hyg. 1998, 59, 710–716. [Google Scholar] [CrossRef]
- Watts, D.M.; Phillips, I.; Callahan, J.D.; Griebenow, W.; Hyams, K.C.; Hayes, A.G. Oropouche Virus Transmission in the Amazon River Basin of Peru. Am. J. Trop. Med. Hyg. 1997, 56, 148–152. [Google Scholar] [CrossRef] [PubMed]
- Watts, D.M.; Russell, K.L.; Wooster, M.T.; Sharp, T.W.; Morrison, A.C.; Kochel, T.J.; Bautista, C.T.; Block, K.; Guevara, C.; Aguilar, P.; et al. Etiologies of Acute Undifferentiated Febrile Illnesses in and near Iquitos from 1993 to 1999 in the Amazon River Basin of Peru. Am. J. Trop. Med. Hyg. 2022, 107, 1114–1128. [Google Scholar] [CrossRef]
- Gaillet, M.; Pichard, C.; Restrepo, J.; Lavergne, A.; Perez, L.; Enfissi, A.; Abboud, P.; Lambert, Y.; Ma, L.; Monot, M.; et al. Outbreak of Oropouche Virus in French Guiana. Emerg. Infect. Dis. 2021, 27, 2711–2714. [Google Scholar] [CrossRef] [PubMed]
- Bonifay, T.; Le Turnier, P.; Epelboin, Y.; Carvalho, L.; De Thoisy, B.; Djossou, F.; Duchemin, J.B.; Dussart, P.; Enfissi, A.; Lavergne, A.; et al. Review on Main Arboviruses Circulating on French Guiana, An Ultra-Peripheric European Region in South America. Viruses 2023, 15, 1268. [Google Scholar] [CrossRef]
- Gomez-Camargo, D.E.; Egurrola-Pedraza, J.A.; Cruz, C.D.; Popuche, D.; Ochoa-Diaz, M.M.; Guevara, C.; Silva, M.; Abente, E.J.; Ampuero, J.S. Evidence of Oropouche Infection, Colombia, 2017. Emerg. Infect. Dis. 2021, 27, 1756–1758. [Google Scholar] [CrossRef]
- Grisales-Nieto, J.C.; Luz, S.L.B.; Alves Do Nascimento, V.; Gomes Naveca, F.; Murcia-Montaño, L.M.; Romero-Vesga, K.N.; Bellido-Cuellar, O.E.; Carvajal-Cortés, J.J. First Case of Oropouche Fever Detected in the International Border Region of the Colombian Amazon: Clinical Characteristics and Molecular Diagnosis. Mem. Inst. Oswaldo. Cruz. 2024, 119, e230221. [Google Scholar] [CrossRef]
- Silva-Ramos, C.R.; Faccini-Martínez, Á.A.; Serna-Rivera, C.C.; Mattar, S.; Hidalgo, M. Etiologies of Zoonotic Tropical Febrile Illnesses That Are Not Part of the Notifiable Diseases in Colombia. Microorganisms 2023, 11, 2154. [Google Scholar] [CrossRef]
- Wise, E.L.; Pullan, S.T.; Marquez, S.; Paz, V.; Mosquera, J.D.; Zapata, S.; Jackson, S.K.; Fejer, G.; Trueba, G.; Logue, C.H.; et al. Isolation of Oropouche Virus from Febrile Patient, Ecuador. Emerg. Infect. Dis. 2018, 24, 935–937. [Google Scholar] [CrossRef] [PubMed]
- Manock, S.R.; Jacobsen, K.H.; Brito De Bravo, N.; Russell, K.L.; Negrete, M.; Olson, J.G.; Sanchez, J.L.; Blair, P.J.; Smalligan, R.D.; Quist, B.K.; et al. Etiology of Acute Undifferentiated Febrile Illness in the Amazon Basin of Ecuador. Am. J. Trop. Med. Hyg. 2009, 81, 146–151. [Google Scholar] [CrossRef] [PubMed]
- Elbadry, M.A.; Durães-Carvalho, R.; Blohm, G.M.; Stephenson, C.J.; Loeb, J.C.; White, S.K.; Telisma, T.; Chavannes, S.; Beau De Rochars, V.M.; Salemi, M.; et al. Orthobunyaviruses in the Caribbean: Melao and Oropouche Virus Infections in School Children in Haiti in 2014. PLoS Negl. Trop. Dis. 2021, 15, e0009494. [Google Scholar] [CrossRef] [PubMed]
- Forshey, B.M.; Guevara, C.; Laguna-Torres, V.A.; Cespedes, M.; Vargas, J.; Gianella, A.; Vallejo, E.; Madrid, C.; Aguayo, N.; Gotuzzo, E.; et al. Arboviral Etiologies of Acute Febrile Illnesses in Western South America, 2000–2007. PLoS Negl. Trop. Dis. 2010, 4, e787. [Google Scholar] [CrossRef] [PubMed]
- Toledo, M.E.; Monteagudo Diaz, S.; Montenegro Calderón, T.; Kreppel, K.; Van Damme, E.; Vanlerberghe, V. Preparedness for Emerging Epidemic Threats: Detection of Oropouche Circulation in Cuba. Lancet Infect. Dis. 2024, 24, e484. [Google Scholar] [CrossRef]
- Castilletti, C.; Mori, A.; Matucci, A.; Ronzoni, N.; Van Duffel, L.; Rossini, G.; Sponga, P.; D’Errico, M.L.; Rodari, P.; Cristini, F.; et al. Oropouche Fever Cases Diagnosed in Italy in Two Epidemiologically Non-Related Travellers from Cuba, Late May to Early June 2024. Eurosurveillance 2024, 29, 2400362. [Google Scholar] [CrossRef]
- McGregor, B.L.; Connelly, C.R.; Kenney, J.L. Infection, Dissemination, and Transmission Potential of North American Culex Quinquefasciatus, Culex Tarsalis, and Culicoides Sonorensis for Oropouche Virus. Viruses 2021, 13, 226. [Google Scholar] [CrossRef]
- de Mendonça, S.F.; Rocha, M.N.; Ferreira, F.V.; Leite, T.H.J.F.; Amadou, S.C.G.; Sucupira, P.H.F.; Marques, J.T.; Ferreira, A.G.A.; Moreira, L.A. Evaluation of Aedes Aegypti, Aedes Albopictus, and Culex Quinquefasciatus Mosquitoes Competence to Oropouche Virus Infection. Viruses 2021, 13, 755. [Google Scholar] [CrossRef]
- Pinheiro, F.P.; Travassos Da Rosa, A.P.A.; Gomes, M.L.C.; LeDuc, J.W.; Hoch, A.L. Transmission of Oropouche Virus from Man to Hamster by the Midge Culicoides Paraensis. Science 1982, 215, 1251–1253. [Google Scholar] [CrossRef]
- Romero-Alvarez, D.; Escobar, L.E. Vegetation Loss and the 2016 Oropouche Fever Outbreak in Peru. Mem. Inst. Oswaldo. Cruz. 2017, 112, 292–298. [Google Scholar] [CrossRef]
- Roney, M.; Aluwi, M.F.F.M. Oropouche Fever Outbreak Is Emerging Concern in American Countries. Bull. Natl. Res. Cent. 2024, 48, 76. [Google Scholar] [CrossRef]
- Ciuoderis, K.A.; Berg, M.G.; Perez, L.J.; Hadji, A.; Perez-Restrepo, L.S.; Aristizabal, L.C.; Forberg, K.; Yamaguchi, J.; Cardona, A.; Weiss, S.; et al. Oropouche Virus as an Emerging Cause of Acute Febrile Illness in Colombia. Emerg. Microbes Infect. 2022, 11, 2645–2657. [Google Scholar] [CrossRef]
- Romero-Alvarez, D.; Escobar, L.E.; Auguste, A.J.; Del Valle, S.Y.; Manore, C.A. Transmission Risk of Oropouche Fever across the Americas. Infect. Dis. Poverty 2023, 12, 47. [Google Scholar] [CrossRef]
- Silva-Caso, W.; Aguilar-Luis, M.A.; Palomares-Reyes, C.; Mazulis, F.; Weilg, C.; del Valle, L.J.; Espejo-Evaristo, J.; Soto-Febres, F.; Martins-Luna, J.; del Valle-Mendoza, J. First Outbreak of Oropouche Fever Reported in a Non-Endemic Western Region of the Peruvian Amazon: Molecular Diagnosis and Clinical Characteristics. Int. J. Infect. Dis. 2019, 83, 139–144. [Google Scholar] [CrossRef] [PubMed]
- Romero-Alvarez, D.; Escobar, L.E. Oropouche Fever, an Emergent Disease from the Americas. Microbes Infect. 2018, 20, 135–146. [Google Scholar] [CrossRef] [PubMed]
- Fujita, D.M.; Salvador, F.S.; da Silva Nali, L.H.; de Andrade Júnior, H.F. Oropouche in Brazil in 2024. J. Travel. Med. 2024, 31, taae075. [Google Scholar] [CrossRef] [PubMed]
- Lorenz, C.; Chiaravalloti-Neto, F. Brazil Reports an Increased Incidence of Oropouche and Mayaro Fever in the Amazon Region. Travel. Med. Infect. Dis. 2024, 58, 102692. [Google Scholar] [CrossRef]
- Pan American Health Organization. Public Health Risk Assessment Related to Oropouche Virus (OROV) in the Region of the Americas General Risk Statement; Pan American Health Organization: Washington, DC, USA, 2024. [Google Scholar]
- da Silva Azevedo, R.D.S.; Nunes, M.R.T.; Chiang, J.O.; Bensabath, G.; Vasconcelos, H.B.; das Neves Pinto, A.Y.; Martins, L.C.; de Oliveira Monteiro, H.A.; Rodrigues, S.G.; da Costa Vasconcelos, P.F. Reemergence of Oropouche Fever, Northern Brazil. Emerg. Infect. Dis. 2007, 13, 912–915. [Google Scholar] [CrossRef]
- Vasconcelos, H.B.; Azevedo, R.S.S.; Casseb, S.M.; Nunes-Neto, J.P.; Chiang, J.O.; Cantuária, P.C.; Segura, M.N.O.; Martins, L.C.; Monteiro, H.A.O.; Rodrigues, S.G.; et al. Oropouche Fever Epidemic in Northern Brazil: Epidemiology and Molecular Characterization of Isolates. J. Clin. Virol. 2009, 44, 129–133. [Google Scholar] [CrossRef]
- Forato, J.; Meira, C.A.; Claro, I.M.; Amorim, M.R.; de Souza, G.F.; Muraro, S.P.; Toledo-Teixeira, D.A.; Dias, M.F.; Meneses, C.A.R.; Angerami, R.N.; et al. Molecular Epidemiology of Mayaro Virus among Febrile Patients, Roraima State, Brazil, 2018–2021. Emerg. Infect. Dis. 2024, 30, 1013–1016. [Google Scholar] [CrossRef]
- Tesh, R.B. The Emerging Epidemiology of Venezuelan Hemorrhagic Fever and Oropouche Fever in Tropical South America. Ann. N. Y. Acad. Sci. 1994, 740, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Chiang, J.O.; Azevedo, R.S.; Justino, M.C.A.; Matos, H.J.; Cabeça, H.L.S.; Silva, S.P.; Henriques, D.F.; Silva, E.V.P.; Andrade, G.S.S.; Vasconcelos, P.F.; et al. Neurological Disease Caused by Oropouche Virus in Northern Brazil: Should It Be Included in the Scope of Clinical Neurological Diseases? J. Neurovirol. 2021, 27, 626–630. [Google Scholar] [CrossRef] [PubMed]
- Pavon, J.A.R.; Neves, N.A.D.S.; Silva, L.C.F.; Azevedo, F.K.D.; Junior, F.; Nunes, M.R.T.; Slhessarenko, R.D. Neurological Infection by Chikungunya and a Triple Arbovirus Co-Infection in Mato Grosso, Central Western Brazil during 2019. J. Clin. Virol. 2022, 146, 105056. [Google Scholar] [CrossRef]
- Garcia Filho, C.; Silva Lima Neto, A.; Maria Peixoto Cabral Maia, A.; Osvaldo Rodrigues Silva, L.; da Costa Cavalcante, R.; da Silva Monteiro, H.; Carneiro Alves Marques, K.; de Souza Oliveira, R.; de Andrade Cordeiro Gadelha, S.; Nunes de Melo, D.; et al. Vertical Transmission of Oropouche Virus in a Newly Affected Extra-Amazon Region: A Case Study of Fetal Infection and Death in Ceará, Brazil. SciELO Preprints 2024. [Google Scholar] [CrossRef]
- Carlos Bandeira, A.; Fernandes, A.C.; Da, N.; Barbosa, S.; Souza, M.; Da, R.; Saavedra, C.; Pereira, M.; Patricia, S.; Santos, O.; et al. Clinical Profile of Oropouche Fever in Bahia, Brazil: Unexpected Fatal Cases. SciELO Preprints 2024. [Google Scholar] [CrossRef]
- Rojas, A.; Stittleburg, V.; Cardozo, F.; Bopp, N.; Cantero, C.; López, S.; Bernal, C.; Mendoza, L.; Aguilar, P.; Pinsky, B.A.; et al. Real-Time RT-PCR for the Detection and Quantitation of Oropouche Virus. Diagn. Microbiol. Infect. Dis. 2020, 96, 114894. [Google Scholar] [CrossRef]
- Lachmann, R.; Meincke, M.; Grünewald, T.; Brockmann, S.; Frank, C.; Schmidt-Chanasit, J.; Wilking, H. Erste Oropouche-Fieber-Fälle in Deutschland Unter Reiserückkehrenden Aus Kuba. Epidemiol. Bull. 2024, 31, 12–14. [Google Scholar]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G.; Altman, D.; Antes, G.; Atkins, D.; Barbour, V.; Barrowman, N.; Berlin, J.A.; et al. Preferred Reporting Items for Systematic Reviews and Meta-Analyses: The PRISMA Statement. PLoS Med. 2009, 7, 889–896. [Google Scholar] [CrossRef]
- Rethlefsen, M.L.; Kirtley, S.; Waffenschmidt, S.; Ayala, A.P.; Moher, D.; Page, M.J.; Koffel, J.B. PRISMA-S: An Extension to the PRISMA Statement for Reporting Literature Searches in Systematic Reviews. Syst. Rev. 2021, 10, 39. [Google Scholar] [CrossRef]
- Von Hippel, P.T. The Heterogeneity Statistic I2 Can Be Biased in Small Meta-Analyses. BMC Med. Res. Methodol. 2015, 15, 35. [Google Scholar] [CrossRef]
- Krumpal, I. Determinants of Social Desirability Bias in Sensitive Surveys: A Literature Review. Qual. Quant. 2013, 47, 2025–2047. [Google Scholar] [CrossRef]
- National Institutes of Health. OHAT Risk of Bias Rating Tool for Human and Animal Studies; National Institutes of Health: Bethesda, MD, USA, 2015. [Google Scholar]
- Eick, S.M.; Goin, D.E.; Chartres, N.; Lam, J.; Woodruff, T.J. Assessing Risk of Bias in Human Environmental Epidemiology Studies Using Three Tools: Different Conclusions from Different Tools. Syst. Rev. 2020, 9, 249. [Google Scholar] [CrossRef] [PubMed]
- Higgins, J.P.T.; Thompson, S.G.; Spiegelhalter, D.J. A Re-Evaluation of Random-Effects Meta-Analysis. J. R. Stat. Soc. Ser. A Stat. Soc. 2009, 172, 137–159. [Google Scholar] [CrossRef] [PubMed]
- Higgins, J.P.T.; Thompson, S.G.; Deeks, J.J.; Altman, D.G. Measuring Inconsistency in Meta-Analyses. Br. Med. J. 2003, 327, 557–560. [Google Scholar] [CrossRef]
- Begg, C.B.; Mazumdar, M. Operating Characteristics of a Rank Correlation Test for Publication Bias. Biometrics 1994, 50, 1088–1101. [Google Scholar] [CrossRef] [PubMed]
- R Development Core Team. R a Language and Environment for Statistical Computing: Reference Index; R Foundation for Statistical Computing: Vienna, Austria, 2010; ISBN 3900051070. [Google Scholar]
- Haddaway, N.R.; Page, M.J.; Pritchard, C.C.; McGuinness, L.A. PRISMA2020: An R Package and Shiny App for Producing PRISMA 2020-Compliant Flow Diagrams, with Interactivity for Optimised Digital Transparency and Open Synthesis. Campbell Syst. Rev. 2022, 18, e1230. [Google Scholar] [CrossRef]
- Pinheiro, F.P.; Rocha, A.G.; Freitas, R.B.; Ohana, B.A.; Travassos da Rosa, A.P.A.; Rogerio, J.S.; Linhares, A.C. Meningite associada as infeccoes por virus Oropouche. Rev. Inst. Med. Trop. Sao. Paolo. 1982, 24, 246–251. [Google Scholar]
- Vasconcelos, P.F.C.D.; Travassos Da Rosa, J.F.S.; Guerreiro, S.C.; Dégallier, N.; Travassos Da Rosa, E.S.; Travassos Da Rosa, A.P.D.A. Primeiro Registro De Epidemias Causadas Pelo Vírus Oropouche Nos Estados Do Maranhão E Goiás, Brasil. Rev. Inst. Med. Trop. Sao. Paolo. 1989, 31, 271–278. [Google Scholar] [CrossRef]
- Rosa, P.A.T.; Rodrigues, S.G.; Nunes, M.R.T.; Magalhães, T.F.; Rosa, J.F.S.T.; Vasconcelos, P.F.C. Epidemia De Febre Do Oropouche Em Serra Pelada, Município De Curionópolis, Pará, 1994. Rev. Soc. Bras. Med. Trop. 1996, 29, 537–541. [Google Scholar] [CrossRef]
- Watts, D.M.; Lavera, V.; Callahan, J.; Rossi, C.; Oberste, M.S.; Roehrig, J.T.; Cropp, C.B.; Karabatsos, N.; Smith, J.F.; Gubler, D.J.; et al. Venezuelan Equine Encephalitis and Oropouche Virus Infections among Peruvian Army Troops in the Amazon Region of Peru. Am. J. Trop. Med. Hyg. 1997, 56, 661–667. [Google Scholar] [CrossRef]
- De Figueiredo, R.M.P.; Thatcher, B.D.; de Lima, M.L.; Almeida, T.C.; Alecrim, W.D.; de Farias Guerra, M.V. Doenças Exantemáticas e Primeira Epidemia de Dengue Ocorrida Manaus, Amazonas, No Período de 1998–1999. Rev. Soc. Bras. Med. Trop. 2004, 37, 476–479. [Google Scholar] [CrossRef] [PubMed]
- Tavares-Neto, J.; Freitas-Carvalho, J.; Teixeira Nunes, M.R.; Rocha, G.; Rodrigues, S.G.; Damasceno, E.; Darub, R.; Viana, S.; da Costa Vasconcelos, P.F. Pesquisa de Anticorpos Contra Arbovírus e o Vírus Vacinal Da Febre Amarela Em Uma Amostra Da População de Rio Branco, Antes e Três Meses Após a Vacina 17D. Rev. Soc. Bras. Med. Trop. 2004, 37, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Terzian, B.; de Moraes Bronzoni, R.V.; Paiva Drumond, B.; Da Silva-Nunes, M.; Santos da Silva, N.; Ferreira, M.U.; Sperança, M.A.; Nogueira, M.L. Sporadic Oropuche Virus Infection, Acre, Brazil. Emerg. Infect. Dis. 2009, 15, 348–350. [Google Scholar] [CrossRef]
- Cruz, A.C.R.; do Socorro Correa dos Prazeres, A.; Colares Gama, E.; Furtado de Lima, M.; do Socorro Silva Azevedo, R.; Medeiros Neves Casseb, L.; Pinto Nunes Neto, J.; Carício Martins, L.; Oliveira Chiang, J.; Guerreiro Rodrigues, S.; et al. Vigilância Sorológica Para Arbovírus Em Juruti, Pará, Brasil Serological Survey for Arboviruses. Cad. Saude Publica 2009, 25, 2517–2523. [Google Scholar] [CrossRef]
- Mourao, M.P.G.; Bastos, M.S.; Gimaque, J.B.L.; Mota, B.R.; Souza, G.S.; Grimmer, G.H.N.; Galusso, E.S.; Arruda, E.; Figuereido, L.T.M. Oropouche Fever Outbreak, Manaus, Brazil, 2007–2008. Emerg. Infect. Dis. 2009, 16, 2063–2064. [Google Scholar] [CrossRef]
- Alvarez-Falconi, P.P.; Amanda, B.; Ruiz, R. Brote de Fiebre de Oropuche En Bagazán, Martín-Perú: Evaluación Epidemiológica, Manifestaciones Gastrointestinales y Hemorrágicas. Rev. Gastroenterol. Perú 2010, 30–34, 334–340. [Google Scholar]
- Aguilar, P.V.; Barrett, A.D.; Saeed, M.F.; Watts, D.M.; Russell, K.; Guevara, C.; Ampuero, J.S.; Suarez, L.; Cespedes, M.; Montgomery, J.M.; et al. Iquitos Virus: A Novel Reassortant Orthobunyavirus Associated with Human Illness in Peru. PLoS Negl. Trop. Dis. 2011, 5, e1315. [Google Scholar] [CrossRef]
- Bastos, M.D.S.; Figueiredo, L.T.M.; Naveca, F.G.; Monte, R.L.; Lessa, N.; De Figueiredo, R.M.P.; Gimaque, J.B.D.L.; João, G.P.; Ramasawmy, R.; Mourão, M.P.G. Short Report: Identification of Oropouche Orthobunyavirus in the Cerebrospinal Fluid of Three Patients in the Amazonas, Brazil. Am. J. Trop. Med. Hyg. 2012, 86, 732–735. [Google Scholar] [CrossRef] [PubMed]
- Bastos, M.S.; Lessa, N.; Naveca, F.G.; Monte, R.L.; Braga, W.S.; Figueiredo, L.T.M.; Ramasawmy, R.; Mourão, M.P.G. Detection of Herpesvirus, Enterovirus, and Arbovirus Infection in Patients with Suspected Central Nervous System Viral Infection in the Western Brazilian Amazon. J. Med. Virol. 2014, 86, 1522–1527. [Google Scholar] [CrossRef]
- Martins, V.D.C.A.; De Bastos, M.S.; Ramasawmy, R.; De Figueiredo, R.P.; Gimaque, J.B.L.; Braga, W.S.M.; Nogueira, M.L.; Nozawa, S.; Naveca, F.G.; Figueiredo, L.T.M.; et al. Clinical and Virological Descriptive Study in the 2011 Outbreak of Dengue in the Amazonas, Brazil. PLoS ONE 2014, 9, e100535. [Google Scholar] [CrossRef]
- Cardoso, B.F.; Serra, O.P.; Da Silva Heinen, L.B.; Zuchi, N.; De Souza, V.C.; Naveca, F.G.; Dos Santos, M.A.M.; Slhessarenko, R.D. Detection of Oropouche Virus Segment s in Patients and in Culex Quinquefasciatus in the State of Mato Grosso, Brazil. Mem. Inst. Oswaldo. Cruz. 2015, 110, 745–754. [Google Scholar] [CrossRef] [PubMed]
- García, M.P.; Merino, N.S.; Figueroa, D.; Marcelo, A.; Tineo, V.E.; Manrique, C.; Donaires, F.; Céspedes, M.; Cabrera, R. Detection of Oropouche Viral Circulation in Madre de Dios Region, Peru (December 2015 to January 2016). Rev. Peru. Med. Exp. Salud. Publica 2016, 33, 380–381. [Google Scholar] [CrossRef]
- Alva-Urcia, C.; Aguilar-Luis, M.A.; Palomares-Reyes, C.; Silva-Caso, W.; Suarez-Ognio, L.; Weilg, P.; Manrique, C.; Vasquez-Achaya, F.; Del Valle, L.J.; Del Valle-Mendoza, J. Emerging and Reemerging Arboviruses: A New Threat in Eastern Peru. PLoS ONE 2017, 12, e0187897. [Google Scholar] [CrossRef]
- Do Nascimento, V.A.; Santos, J.H.A.; Monteiro, D.C.A.S.; Pessoa, K.P.; Cardoso, A.J.L.; de Souza, V.C.; Abdalla, L.F.; Naveca, F.G. Oropouche Virus Detection in Saliva and Urine. Mem. Inst. Oswaldo. Cruz. 2020, 115, e190338. [Google Scholar] [CrossRef]
- Martins-Luna, J.; Del Valle-Mendoza, J.; Silva-Caso, W.; Sandoval, I.; Del Valle, L.J.; Palomares-Reyes, C.; Carrillo-Ng, H.; Peña-Tuesta, I.; Aguilar-Luis, M.A. Oropouche Infection a Neglected Arbovirus in Patients with Acute Febrile Illness from the Peruvian Coast. BMC Res. Notes 2020, 13, 67. [Google Scholar] [CrossRef] [PubMed]
- dos Santos Fonseca, L.M.; Carvalho, R.H.; Bandeira, A.C.; Sardi, S.I.; Campos, G.S. Oropouche Virus Detection in Febrile Patients’ Saliva and Urine Samples. Jpn. J. Infect. Dis. 2020, 73, 164–165. [Google Scholar] [CrossRef]
- Saatkamp, C.J.; Rodrigues, L.R.R.; Pereira, A.M.N.; Coelho, J.A.; Marques, R.G.B.; Souza, V.C.D.; Nascimento, V.A.D.; Saatkamp, J.G.D.S.; Naveca, F.G.; Figueiredo, R.M.P.D. Mayaro Virus Detection in the Western Region of Pará State, Brazil. Rev. Soc. Bras. Med. Trop. 2021, 54, e0055. [Google Scholar] [CrossRef] [PubMed]
- Salgado, B.B.; de Jesus Maués, F.C.; Pereira, R.L.; Chiang, J.O.; de Oliveira Freitas, M.N.; Ferreira, M.S.; Martins, L.C.; da Costa Vasconcelos, P.F.; Ganoza, C.; Lalwani, P. Prevalence of Arbovirus Antibodies in Young Healthy Adult Population in Brazil. Parasit. Vectors 2021, 14, 403. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, V.L.; Azevedo, R.S.S.; Carvalho, V.L.; Azevedo, R.S.; Henriques, D.F.; Cruz, A.C.R.; Vasconcelos, P.F.C.; Martins, L.C. Arbovirus Outbreak in a Rural Region of the Brazilian Amazon. J. Clin. Virol. 2022, 150–151, 105155. [Google Scholar] [CrossRef]
- Dias, H.G.; Dos Santos, F.B.; Pauvolid-Corrêa, A. An Overview of Neglected Orthobunyaviruses in Brazil. Viruses 2022, 14, 987. [Google Scholar] [CrossRef]
- Gil-Mora, J.; Acevedo-Gutierrez, L.Y.; Betancourt-Ruiz, P.L.; Martınez-Diaz, H.C.; Fernandez, D.; Bopp, N.E.; Olaya-Masmela, L.A.; Bolaños, E.; Benavides, E.; Villasante-Tezanos, A.; et al. Arbovirus Antibody Seroprevalence in the Human Population from Cauca, Colombia. Am. J. Trop. Med. Hyg. 2022, 107, 1218–1225. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves Maciel, L.H.; Vieira da Rocha Neto, C.; Ferreira Martins, Y.; de Azevedo Furtado, F.; Cunha Teixeira, P.; Oliveira Dias, M.Y.; Batista Rodrigues, Y.K.; Ribeiro Piauilino, I.C.; Pinto, S.D.; Côrte Alencar, A.C.; et al. Prevalence of Arboviruses and Other Infectious Causes of Skin Rash in Patients Treated at a Tertiary Health Unit in the Brazilian Amazon. PLoS Negl. Trop. Dis. 2022, 16, e0010727. [Google Scholar] [CrossRef] [PubMed]
- de Lima, R.C.; Dias, H.G.; de Souza, T.M.A.; Familiar-Macedo, D.; Ribeiro, E.D.; Corrêa, V.C.E.; Pauvolid-Corrêa, A.; de Azeredo, E.L.; dos Santos, F.B. Oropouche Virus Exposure in Febrile Patients during Chikungunya Virus Introduction in the State of Amapá, Amazon Region, Brazil. Pathogens 2024, 13, 469. [Google Scholar] [CrossRef] [PubMed]
- Moreira, H.M.; Sgorlon, G.; Queiroz, J.A.S.; Roca, T.P.; Ribeiro, J.; Teixeira, K.S.; Passos-Silva, A.M.; Araújo, A.; Gasparelo, N.W.F.; Dos Santos, A.D. Outbreak of Oropouche Virus in Frontier Regions in Western Amazon. Microbiol. Spectr. 2024, 12, e0162923. [Google Scholar] [CrossRef] [PubMed]
- Scachetti, G.C.; Forato, J.; Claro, I.M.; Hua, X.; Salgado, B.B.; Vieira, A.; Simeoni, C.L.; C Barbosa, A.R.; Rosa, I.L.; de Souza, G.F.; et al. Reemergence of Oropouche Virus between 2023 and 2024 in Brazil. MedRxiv 2024. 2024.07.27.24310296 (epub ahead of print). [Google Scholar] [CrossRef]
- Silva, J.C.D.O.; Siqueira, I.R.F.; Dornelas, L.F.; Ribeiro, C.M.; Gomes, J.P.B.; Guadagnin, I.W.; Pereira, A.R.; Julião, G.R.; Camargo, J.D.S.A.A.; Basano, S.A.; et al. Occurrence of Arbovirus Infections in Two Riverine Populations in the Municipality of Humaitá, Amazonas, Brazil. Rev. Soc. Bras. Med. Trop. 2024, 57, e004032024. [Google Scholar] [CrossRef]
- Sánchez-Lerma, L.; Rojas-Gulloso, A.; Miranda, J.; Tique, V.; Patiño, L.H.; Rodriguez, D.; Contreras, V.; Paniz-Mondolfi, A.; Pavas, N.; Ramírez, J.D.; et al. Unexpected Arboviruses Found in an Epidemiological Surveillance of Acute Tropical Febrile Syndrome in the Department of Meta, Eastern Colombia. J. Infect. Public. Health 2024, 17, 102510. [Google Scholar] [CrossRef]
- Dias, H.G.; de Lima, R.C.; Barbosa, L.S.; de Souza, T.M.A.; Badolato-Correa, J.; Maia, L.M.S.; da Silva Ferreira, R.; da Silva Neves, N.A.; de Souza Costa, M.C.; Martins, L.R.; et al. Retrospective Molecular Investigation of Mayaro and Oropouche Viruses at the Human-Animal Interface in West-Central Brazil, 2016–2018. PLoS ONE 2022, 17, e0277612. [Google Scholar] [CrossRef]
- National Toxicology Program. Handbook for Conducting a Literature-Based Health Assessment Using OHAT Approach for Systematic Review and Evidence Integration; National Toxicology Program: Triangle Park, NC, USA, 2019. [Google Scholar]
- Liu, B.M. Epidemiological and Clinical Overview of the 2024 Oropouche Virus Disease Outbreaks, an Emerging/Re-Emerging Neurotropic Arboviral Disease and Global Public Health Threat. J. Med. Virol. 2024, 96, https. [Google Scholar] [CrossRef]
- Molyneux, D.H.; Hotez, P.J.; Fenwick, A. “Rapid-Impact Interventions”: How a Policy of Integrated Control for Africa’s Neglected Tropical Diseases Could Benefit the Poor. PLoS Med. 2005, 2, 1064–1070. [Google Scholar] [CrossRef]
- Hotez, P.J.; Molyneux, D.H.; Fenwick, A.; Ottesen, E.; Sachs, S.E.; Sachs, J.D. Incorporating a Rapid-Impact Package for Neglected Tropical Diseases with Programs for HIV/AIDS, Tuberculosis, and Malaria: A Comprehensive pro-Poor Health Policy and Strategy for the Developing World. PLoS Med. 2006, 3, 576–584. [Google Scholar] [CrossRef] [PubMed]
- Hotez, P.J.; Molyneux, D.H.; Fenwick, A.; Kumaresan, J.; Ehrlich Sachs, S.; Sachs, J.D.; Savioli, L. Control of Neglected Tropical Diseases. N. Engl. J. Med. 2007, 357, 1018–1027. [Google Scholar] [CrossRef] [PubMed]
- Brady, M.A.; Hooper, P.J.; Ottesen, E.A. Projected Benefits from Integrating NTD Programs in Sub-Saharan Africa. Trends Parasitol. 2006, 22, 285–291. [Google Scholar] [CrossRef] [PubMed]
- Yajima, A.; Lin, Z.; Mohamed, A.J.; Dash, A.P.; Rijal, S. Finishing the Task of Eliminating Neglected Tropical Diseases(NTDs) in WHO South-East Asia Region: Promises Kept, Challenges, and the Way Forward. Lancet Reg. Health-Southeast Asia 2023, 18, 100302. [Google Scholar] [CrossRef] [PubMed]
- United Nations. Preliminary Overview of the Economies of Latin America and the Caribbean 2023; United Nations: Santiago, Chile, 2023. [Google Scholar]
- Giglio, E. Extractivism and Its Socio-Environmental Impact in South America. Overview of the “Lithium Triangle”. America. Crit. 2021, 5, 47–53. [Google Scholar] [CrossRef]
- Abramo, L.; Cecchini, S.; Ullmann, H. Addressing Health Inequalities in Latin America: The Role of Social Protection. Ciência Saúde Coletiva 2020, 25, 1587–1598. [Google Scholar] [CrossRef]
- World Economic Forum Latin America’s Cities Are Ready to Take Off. But Their Infrastructure Is Failing Them. Available online: https://www.weforum.org/agenda/2018/06/latin-america-cities-urbanization-infrastructure-failing-robert-muggah/#:~:text=Sprawling%20Latin%20American%20cities%20often,most%20congested%20on%20the%20planet. (accessed on 28 August 2024).
- Siedner, M.J.; Ryan, E.T.; Bogoch, I.I. Gone or Forgotten? The Rise and Fall of Zika Virus. Lancet Public Health 2018, 3, e109–e110. [Google Scholar] [CrossRef]
- Baud, D.; Gubler, D.J.; Schaub, B.; Lanteri, M.C.; Musso, D. An Update on Zika Virus Infection. Lancet 2017, 390, 2099–2109. [Google Scholar] [CrossRef]
- Ly, H. Dengue Fever in the Americas. Virulence 2024, 15, 2375551. [Google Scholar] [CrossRef]
- Travassos Da Rosa, J.F.; De Souza, W.M.; De Paula Pinheiro, F.; Figueiredo, M.L.; Cardoso, J.F.; Acrani, G.O.; Teixeira Nunes, M.R. Oropouche Virus: Clinical, Epidemiological, and Molecular Aspects of a Neglected Orthobunyavirus. Am. J. Trop. Med. Hyg. 2017, 96, 1019–1030. [Google Scholar] [CrossRef]
- Collins, Á.B.; Doherty, M.L.; Barrett, D.J.; Mee, J.F. Schmallenberg Virus: A Systematic International Literature Review (2011–2019) from an Irish Perspective. Ir. Vet. J. 2019, 72, 9. [Google Scholar] [CrossRef] [PubMed]
- Wernike, K.; Beer, M. Schmallenberg Virus: To Vaccinate, or Not to Vaccinate? Vaccines 2020, 8, 287. [Google Scholar] [CrossRef]
- Reusken, C.; van den Wijngaard, C.; van Beek, P.; Beer, M.; Bouwstra, R.; Godeke, G.J.; Isken, L.; van den Kerkhof, H.; van Pelt, W.; van der Poel, W.; et al. Lack of Evidence for Zoonotic Transmission of Schmallenberg Virus. Emerg. Infect. Dis. 2012, 18, 1746–1754. [Google Scholar] [CrossRef] [PubMed]
- Mansfield, K.L.; Folly, A.J.; Hernández-Triana, L.M.; Sewgobind, S.; Johnson, N. Batai Orthobunyavirus: An Emerging Mosquito-Borne Virus in Europe. Viruses 2022, 14, 1868. [Google Scholar] [CrossRef] [PubMed]
- Charles, J.A. Akabane Virus. Vet. Clin. N. Am. Food Anim. Pr. 1994, 10, 525–546. [Google Scholar] [CrossRef]
- Brenner, J.; Rotenberg, D.; Jaakobi, S.; Stram, Y.; Guini-Rubinstein, M.; Menasherov, S.; Bernstein, M.; Yaakobovitch, Y.; David, D.; Perl, S. Analogie e Differenze Tra Malattia di Akabane e Altre Arbovirosi. Vet. Ital. 2016, 52, 353–362. [Google Scholar] [CrossRef]
- Kirkland, P.D. Akabane Virus Infection the Virus. Rev. Sci. Tech. Off. Int. Epiz. 2015, 34, 403–410. [Google Scholar] [CrossRef]
- Santos, R.I.; Bueno-Júnior, L.S.; Ruggiero, R.N.; Almeida, M.F.; Silva, M.L.; Paula, F.E.; Correa, V.M.A.; Arruda, E. Spread of Oropouche Virus into the Central Nervous System in Mouse. Viruses 2014, 6, 3827–3836. [Google Scholar] [CrossRef] [PubMed]
- Turell, M.J.; Gozalo, A.S.; Guevara, C.; Schoeler, G.B.; Carbajal, F.; López-Sifuentes, V.M.; Watts, D.M. Lack of Evidence of Sylvatic Transmission of Dengue Viruses in the Amazon Rainforest Near Iquitos, Peru. Vector-Borne Zoonotic Dis. 2019, 19, 685–689. [Google Scholar] [CrossRef]
- Alonso, L.E.; Romeo Aznar, V.; Solari, H.G. Why Is Aedes Aegypti Moving South in South America? Mathematics 2022, 10, 4510. [Google Scholar] [CrossRef]
- Mulderij-Jansen, V.; Pundir, P.; Grillet, M.E.; Lakiang, T.; Gerstenbluth, I.; Duits, A.; Tami, A.; Bailey, A. Effectiveness of Aedes-Borne Infectious Disease Control in Latin America and the Caribbean Region: A Scoping Review. PLoS ONE 2022, 17, e0277038. [Google Scholar] [CrossRef] [PubMed]
- Bardach, A.E.; García-Perdomo, H.A.; Alcaraz, A.; Tapia López, E.; Gándara, R.A.R.; Ruvinsky, S.; Ciapponi, A. Interventions for the Control of Aedes Aegypti in Latin America and the Caribbean: Systematic Review and Meta-Analysis. Trop. Med. Int. Health 2019, 24, 530–552. [Google Scholar] [CrossRef] [PubMed]
- Remoli, M.E.; Fiorentini, C.; Marchi, A.; Di Renzi, S.; Vonesch, N.; Peri, V.M.; Bastianini, L.; Rossi, S.; Bartoccini, G.; Kuttappasery, M.L.; et al. Seroprevalence Survey of Arboviruses in Workers from Tuscany, Italy. Med. Del. Lav. 2018, 109, 125–131. [Google Scholar] [CrossRef]
- Mingione, M.; Branda, F.; Maruotti, A.; Ciccozzi, M.; Mazzoli, S. Monitoring the West Nile Virus Outbreaks in Italy Using Open Access Data. Sci. Data 2023, 10, 777. [Google Scholar] [CrossRef]
- Rossi, B.; Barreca, F.; Benvenuto, D.; Braccialarghe, N.; Campogiani, L.; Lodi, A.; Aguglia, C.; Cavasio, R.A.; Giacalone, M.L.; Kontogiannis, D.; et al. Human Arboviral Infections in Italy: Past, Current, and Future Challenges. Viruses 2023, 15, 368. [Google Scholar] [CrossRef]
- World Health Organization. Global Vector Control Response 2017–2030; World Health Organization: Geneva, Switzerland, 2017. [Google Scholar]
- European Centre for Disease Prevention and Control (ECDC). ECDC One Health Framework; European Centre for Disease Prevention and Control (ECDC): Stockholm, Sweden, 2024. [Google Scholar]
- Ricco, M.; Peruzzi, S.; Balzarini, F. Epidemiology of West Nile Virus Infections in Humans, Italy, 2012–2020: A Summary of Available Evidences. Trop. Med. Infect. Dis. 2021, 6, 61. [Google Scholar] [CrossRef] [PubMed]
- Riccò, M.; Peruzzi, S.; Balzarini, F.; Zaniboni, A.; Ranzieri, S. Dengue Fever in Italy: The “Eternal Return” of an Emerging Arboviral Disease. Trop. Med. Infect. Dis. 2022, 7, 10. [Google Scholar] [CrossRef]
- Riccò, M.; Peruzzi, S. Epidemiology of Toscana Virus in Italy (2018–2020), a Summary of Available Evidences. Acta Biomed. 2021, 92, e2021230. [Google Scholar]
- Ministero della Salute (DG Prevenzione Sanitataria). Piano Nazionale Di Sorveglianza e Risposta Alle Arbovirosi Trasmesse Da Zanzare Invasive (Aedes Sp.) Con Particolare Riferimento Ai Virus Chikungunya, Dengue e Zika-2018.
- Italian Health Ministry. Piano Nazionale di Prevenzione, Sorveglianza e Risposta alle Arbovirosi (PNA) 2020–2025; Italian Health Ministry: Rome, Italy, 2019. [Google Scholar]
- Riccò, M. Epidemiology of Tick-Borne Encephalitis in North-Eastern Italy (2017–2020): International Insights from National Notification Reports. Acta Biomed. 2021, 92, e2021229. [Google Scholar]
- Roberto Teixeira Nunes, M.; Carício Martins, L.; Guerreiro Rodrigues, S.; Oliveira Chiang, J.; do Socorro da Silva Azevedo, R.; Travassos da Rosa, A.P.; Fernando da Costa Vasconcelos, P. Oropouche Virus Isolation, Southeast Brazil. Emerg. Infect. Dis. 2005, 11, 1610–1613. [Google Scholar] [CrossRef]
- Unidas, N.; Nations, U. Population Trends in Latin America and the Caribbean: Demographic Effects of the COVID-19 Pandemic. In Latin America and the Caribbean Demographic Observatory 2022; United Nations Publications: New York, NY, USA, 2022. [Google Scholar]
- Esterhuizen, T.M.; Thabane, L. Con: Meta-Analysis: Some Key Limitations and Potential Solutions. Nephrol. Dial. Transplant. 2016, 31, 882–885. [Google Scholar] [CrossRef] [PubMed]
- Dettori, J.R.; Norvell, D.C.; Chapman, J.R. Fixed-Effect vs Random-Effects Models for Meta-Analysis: 3 Points to Consider. Glob. Spine J. 2022, 12, 1624–1626. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study | Country | Timeframe | Area | Sampling Strategy | Sampled Medium | Laboratory Analysis | Sample Size (N) | Positive (n/N, %) |
|---|---|---|---|---|---|---|---|---|
| Pinheiro et al., 1976 [10] | Brazil | February 1975–April 1975 | State of Pará (Mojoì, Palhal, and nearby villages) | All subjects with acute febrile illness during local outbreak | Blood | Virus isolation | 247 | 69 (27.94%) |
| Serology (hemagglutination) | 282 | 119 (42.20%) | ||||||
| LeDuc et al., 1981 [11] | Brazil | July 1978– September 1978 May 1979–June 1979 | State of Pará (Quatro Bocas and Tome Acu) | All subjects with acute febrile illness during local outbreak | Blood | Serology (hemagglutination) | 555 | 164 (29.55%) |
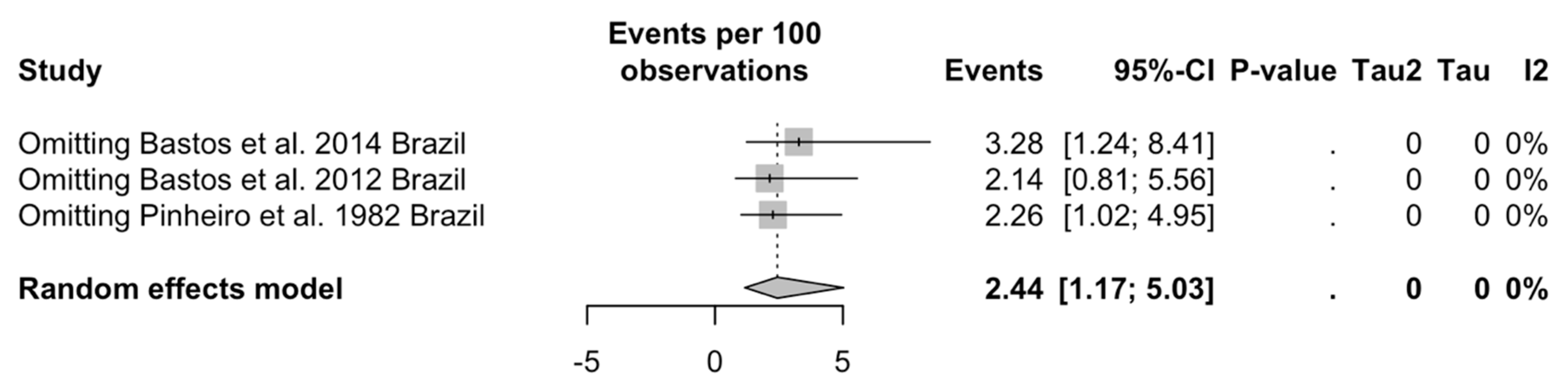
| Pinheiro et al., 1982 [60] | Brazil | 1980 | State of Pará | All subjects with suspected meningitis/encephalitis | CSF | Virus isolation | 22 | 1 (4.55%) |
| Detection of Ig in CSF (hemagglutination) | 22 | 3 (13.64%) | ||||||
| Borborema et al., 1982 [12] | Brazil | May 1981–July 1981 | State of Amazonas (Barcelos and Manaus) | All subjects with acute febrile illness during local outbreak | Blood | Serology (hemagglutination) | 760 | 254 (33.42%) |
| Vasconcelos et al., 1989 [61] | Brazil | 1987 | State of Maranhao (Porto Franco) | All subjects with acute febrile illness during local outbreak | Blood | Virus isolation | 75 | 22 (29.33%) |
| Serology (hemagglutination + ELISA (IgM)) | 197 | 128 (64.97%) | ||||||
| Rosa et al., 1996 [62] | Brazil | June 1994 | State of Pará | Random sampling among residents | Blood | Virus isolation | 296 | 10 (3.38%) |
| Serology (hemagglutination + ELISA (IgG)) | 296 | 245 (82.77%) | ||||||
| Watts et al., 1997 [63] | Peru | 1992 | Loreto Department (Manacamiri, Padre Cocha, Porvenir, Primavera, and Villa Punchana) | Random sampling among residents Follow-up among subjects with negative samples | Blood | ELISA (IgG) | 1616 | 448 (27.72%) |
| Baisley et al., 1998 [13] | Brazil | June 1996–September 1999 | State of Amazonas (Santa Clara) | Random sampling among residents (age > 5 years) | Blood | Serology (hemagglutination + ELISA (IgG)) | 1227 | 390 (31.78%) |
| de [63] Figueredo et al., 2004 [64] | Brazil | March 1998–December 1999 | State of Amazonas | All subjects with acute febrile illness during local outbreak | Blood | Serology (hemagglutination + ELISA (IgM)) | 8557 | 3 (0.04%) |
| Tavares-Neto et al., 2004 [65] | Brazil | 14 August 1999 | State of Acre (Rio Branco) | Random sampling among residents (during HBV vaccination campaign) | Blood | Serology (hemagglutination + ELISA (IgM)) | 394 | 9 (2.28%) |
| da Silva Azevedo et al., 2007 [39] | Brazil | March 2003–May 2003 July 2004–August 2004 | State of Pará (Parauapebas and Porto de Moz) | All subjects with acute febrile illness during local outbreak | Blood | Serology (hemagglutination + ELISA (IgM)) | 234 | 93 (39.74%) |
| Bernarders Terzian et al., 2009 [66] | Brazil | March 2004–October 2006 | State of Acre | All subjects with acute febrile illness during local outbreak (age > 5 years) | Blood | RT-qPCR | 69 | 1 (1.45%) |
| Serology (plaque reduction neutralization test) | 357 | 6 (1.68%) | ||||||
| Cruz et al., 2009 [67] | Brazil | October 2006–December 2007 | State of Pará | All subjects with acute febrile illness during local outbreak | Blood | Serology (hemagglutination) | 1597 | 90 (5.64%) |
| Serology (hemagglutination + ELISA (IgM)) | 1597 | 23 (1.44%) | ||||||
| Manock et al., 2009 [22] | Ecuador | Arril 2001–September 2004 | Pastaza Province | All subjects with acute febrile illness during local outbreak | Blood | RT-qPCR | 229 | 0 (-) |
| Serology (plaque reduction neutralization test) | 304 | 1 (0.32%) | ||||||
| Mourao et al., 2009 [68] | Brazil | January 2007–November 2008 | State of Amazonas (Manaus) | All subjects with acute febrile illness during local outbreak (age > 5 years) | Blood | Serology (ELISA (IgM)) | 631 | 128 (20.29%) |
| Vasconcelos et al., 2009 [40] | Brazil | May 2006–June 2006 | State of Pará (Magalhaes Barata and Maracana) | All subjects with acute febrile illness during local outbreak | Blood | Serology (hemagglutination + ELISA (IgM)) | 744 | 113 (15.19%) |
| Alvarez-Falconi et al., 2010 [69] | Peru | May 2010 | Loreto Department (Bagazan) | All subjects with acute febrile illness during local outbreak | Blood | Serology (ELISA (IgM)) | 171 | 108 (63.16%) |
| Forshey et al., 2010 [24] | Bolivia | 2000–2007 | Region of Cochabamba, Conception, Magdalena, and Santa Crus | All subjects with acute febrile illness during local outbreak (age > 5 years) Children with suspected Dengue hemorrhagic fever | Blood | Indirect immunofluorescence assay followed by RT-qPCR | 2089 | 0 (-) |
| Ecuador | Region of Guayaquil | 350 | 0 (-) | |||||
| Peru | Departments of Iquitos, La Merced, Padre Maldonado, Piura, Tumbes, and Yurimaguas | 18201 | 18 (0.10%) | |||||
| Paraguay | Region of Asunción | 240 | 0 (-) | |||||
| Aguilar et al., 2011 [70] | Peru | 1995–2006 | Loreto Department (Iquitos) | All subjects with acute febrile illness during local outbreak | Blood | Serology (plaque reduction neutralization test) | 1037 | 154 (14.85%) |
| 2006 | 1037 | 2 (0.19%) | ||||||
| Bastos et al., 2012 [71] | Brazil | 2005–2010 | Amazonas | All subjects with suspected CNS infection | CSF | RT-qPCR | 100 | 3 (3.00%) |
| Bastos et al., 2014 [72] | Brazil | January 2010–August 2012 | Amazonas | All subjects with suspected CNS infection | CSF | RT-qPCR | 165 | 3 (1.82%) |
| Martins et al., 2014 [73] | Brazil | January 2011–May 2011 | Amazonas (Manaus) | All subjects with acute febrile illness during local outbreak | Blood | RT-qPCR | 677 | 0 (-) |
| Cardoso et al., 2015 [74] | Brazil | October 2011–July 2012 | Mato Groso | All subjects with acute febrile illness during local outbreak | Blood | RT-qPCR | 529 | 5 (0.95%) |
| Garcia et al., 2016 [75] | Peru | 13 December 2014–8 January 2016 | Madre de Dios | Specimens from individuals sampled for Dengue and Leptospira | Blood | RT-qPCR | 508 | 19 (3.74%) |
| Virus isolation | 508 | 32 (6.30%) | ||||||
| Serology (ELISA (IgM)) | 508 | 122 (24.02%) | ||||||
| Alva-Urcia et al., 2017 [76] | Peru | January 2016–March 2016 | Madre de Dios | All subjects with acute febrile illness during local outbreak | Blood | RT-qPCR | 139 | 12 (8.63%) |
| Silva-Caso et al., 2019 [34] | Peru | January 2016–July 2016 | Huanuco Region | Adults with acute febrile illness lasting < 7 days | Blood | RT-qPCR | 268 | 46 (17.16%) |
| do Nascimiento et al., 2020 [77] | Brazil | February 2016–June 2016 | Amazonas | All subjects with acute febrile illness during local outbreak | Blood | RT-qPCR | 352 | 5 (1.42%) |
| Martins-Luna et al., 2020 [78] | Peru | February 2016–September 2016 | Piura Region | Adults with acute febrile illness | Blood | RT-qPCR | 496 | 131 (26.41%) |
| Rojas et al., 2020 [47] | Paraguay | April 2019 | Not reported | Random sampling among people with suspected arboviral illness | Blood | RT-qPCR | 100 | 0 (-) |
| Salvador et al., 2020 [79] | Brazil | November 2016–December 2017 | Bahia (Salvador) | Subjects referring to a private local hospital with Dengue-like symptoms | Blood | RT-qPCR | 53 | 2 (3.77%) |
| Elbadry et al., 2021 [23] | Haiti | 2014 | Gressier | All children from a local school with an acute febrile illness | Blood | RT-qPCR | 1250 | 1 (0.08%) |
| Gaillet et al., 2021 [16] | French Guyana | 11 August 2020–15 October 2020 | Saúl | All cases with Dengue-like symptoms | Blood | RT-qPCR on microneutralization test | 95 | 23 (24.21%) |
| Saatkamp et al., 2021 [80] | Brazil | 2016 | State of Pará | All febrile subjects of adult age with an acute febrile status | Blood | RT-qPCR | 49 | 0 (-) |
| Salgado et al., 2021 [81] | Brazil | Jaunuary 2014–December 2015 | Nationwide | Random sampling from Brazilian armed forces | Blood | Serology (Hemagglutination test) | 298 | 2 (0.67%) |
| Carvalho et al., 2022 [82] | Brazil | Jaunuary 2018–February 2018 | State of Pará | All subjects reporting a febrile illness in the previous 30 days or had contact with them | Blood | Virus isolation | 94 | 14 (14.89%) |
| Serology (ELISA (IgM)) | 94 | 36 (38.30%) | ||||||
| Ciuoderis et al., 2022 [32] | Colombia | February 2019–Jaunuary 2022 | Regions of Calì, Cucuta, Leticia, and Villavicencio | All febrile subjects aged over 5 years | Blood | RT-qPCR | 2967 | 105 (3.54%) |
| Dias et al., 2022 [83] | Brazil | February 2016–March 2016 | Mato Groso | Retrospective analysis of samples collected in subjects with a febrile illness from < 7 days | Blood | RT-qPCR | 106 | 0 (-) |
| Gil-Mora et al., 2022 [84] | Colombia | 2018 | Cauca Department | All subjects > 18 years from the parent municipalities | Blood | Serology (plaque reduction neutralization test) | 505 | 10 (1.98%) |
| Gonçalves Maciel et al., 2022 [85] | Peru | February 2018–May 2019 | Puerto Maldonado, Piura, and Huanuco | All subjects reporting skin rash | Blood | RT-qPCR | 340 | 0 (-) |
| Watts et al., 2022 [15] | Peru | 1993–1997 | Loreto department | All febrile subjects aged 1 to 60 years, symptoms lasting < 5 days | Blood | IFAT | 6607 | 68 (1.03%) |
| De Lima et al., 2024 [86] | Brazil | August 2014–May 2015 | Amapà region | All subjects with acute febrile illness during local outbreak | Blood | RT-qPCR | 166 | 0 (-) |
| Serology (plaque reduction neutralization test) | 166 | 17 (10.24%) | ||||||
| Forato et al., 2024 [41] | Brazil | December 2018–December 2021 | Roraima (11 municipalities) | All subjects with acute febrile illness during local outbreak | Blood | RT-qPCR | 883 | 0 (-) |
| Grisales-Nieto et al., 2024 [19] | Colombia + Brazil | November 2020 | Department of Leticia and Amazonas (Colombia) and State of Amazonas (Brazil) | Random sampling from residents negative to other arboviruses | Blood | RT-qPCR | 175 | 1 (0.57%) |
| Moreira et al., 2024 [87] | Brazil | January 2022–March 2023 | States of Rondonia and Amazonas | All subjects with acute febrile illness lasting 5 to 7 days during local outbreak Excluded indigenous people, pregnant women, and all subjects with positive testing for other arboviruses | Blood | RT-qPCR | 351 | 27 (7.69%) |
| Scachetti et al., 2024 [88] | Brazil | December 2023–March 2024 | State of Amazonas | All subjects with acute febrile illness during local outbreak | Blood | RT-qPCR | 93 | 10 (10.75%) |
| Silva et al., 2024 [89] | Brazil | 2019–2020 | State of Amazonas | All subjects > 3 years from the communities of Cararà and Espirito Santo | Blood | Serology (immunochromatography IgM) | 205 | 0 (-) |
| Serology (immunochromatography IgG) | 205 | 0 (-) | ||||||
| Sanchez-Lerma et al., 2024 [90] | Colombia | January 2021–June 2023 | Department of Meta | All subjects with acute febrile illness during local outbreak | Blood | RT-qPCR | 100 | 0 (-) |
| Total Samples (N) | OROV-Positive Cases (n/N, %) | Risk Ratio (95%CI) | |
|---|---|---|---|
| Overall | 32,439 | 532, 1.64% | - |
| Timeframe | |||
| Before 1990 | 344 | 92, 26.74% | REFERENCE |
| 1991–2010 | 21,178 | 19, 0.09% | 0.003 (0.002; 0.005) |
| After 2010 | 10,917 | 421, 3.86% | 0.144 (0.118; 0.175) |
| Country | |||
| Bolivia | 2089 | 0, - | 0.006 (0.001; 0.096) |
| Brazil | 4030 | 162, 4.02% | REFERENCE |
| Colombia | 3067 | 105, 3.42% | 0.852 (0.669; 1.084) |
| Ecuador | 579 | 0, - | 0.021 (0.001; 0.344) |
| French Guyana | 95 | 23, 24.21% | 6.023 (4.092; 8.864) |
| Haiti | 1250 | 1, 0.08% | 0.020 (0.003; 0.142) |
| Paraguay | 340 | 0, - | 0.037 (0.002; 0.586) |
| Peru | 20,989 | 241, 1.15% | 0.286 (0.235; 0.348) |
| Sample | |||
| Blood | 32,152 | 525, 1.63% | REFERENCE |
| CSF | 287 | 7, 2.44% | 1.494 (0.715; 3.120) |
| Diagnostic procedure | |||
| Virus isolation | 946 | 138, 14.59% | 4.347 (3.612; 5.231) |
| RT-qPCR | 10,518 | 353, 3.36% | REFERENCE |
| RT-qPCR + IFAT | 20,880 | 18, 0.08% | 0.026 (0.016; 0.041) |
| RT-qPCR + MNT | 95 | 23, 24.21% | 7.214 (4.981; 10.446) |
| Total Samples (N) | OROV-Positive Cases (n/N, %) | Risk Ratio (95%CI) | |
|---|---|---|---|
| Overall | 24,470 | 1630, 6.66% | - |
| Timeframe | |||
| Before 1990 | 1816 | 668, 36.78% | REFERENCE |
| 1991–2010 | 14,192 | 565, 3.98% | 0.108 (0.097; 0.120) |
| After 2010 | 8462 | 397, 4.69% | 0.128 (0.114; 0.143) |
| Country | |||
| Bolivia | 2089 | 0, - | 0.003 (0.001; 0.051) |
| Brazil | 15,793 | 1177, 7.45% | REFERENCE |
| Colombia | 50 | 0, - | 0.134 (0.009; 2.117) |
| Ecuador | 304 | 1, 0.33% | 0.044 (0.006; 0.313) |
| Peru | 8323 | 452, 5.43% | 0.729 (0.656; 0.809) |
| Antibody | |||
| IgG/IgM | 13,030 | 898, 6.89% | 1.077 (0.980; 1.184) |
| IgM | 11,440 | 732, 6.40% | REFERENCE |
| Sample | |||
| Blood | 24,448 | 1627, 6.65% | REFERENCE |
| CSF | 22 | 3, 13.64% | 2.049 (0.715; 5.871) |
| Diagnostic procedure | |||
| ELISA | 13,037 | 755, 5.79% | REFERENCE |
| HAT | 3216 | 630, 19.59% | 3.383 (3.065; 3.733) |
| PRT | 1610 | 177, 10.99% | 1.898 (1.625; 2.217) |
| IFAT | 6607 | 68, 1.03% | 0.178 (0.139; 0.227) |
| Total Sample (N) | OROV-Positive Cases (n/N, %) | Risk Ratio (95%CI) | |
|---|---|---|---|
| Overall | 5247 | 1113, 21.21% | - |
| Timeframe | |||
| Before 1990 | 0 | - | - |
| 1991–2010 | 4085 | 1101, 26.95% | REFERENCE |
| After 2010 | 1162 | 12, 1.03% | 0.038 (0.022; 0.067) |
| Country | |||
| Brazil | 2625 | 646, 24.61% | REFERENCE |
| Colombia | 455 | 10, 2.20% | 0.089 (0.048; 0.165) |
| Peru | 2168 | 457, 21.08% | 0.857 (0.771; 0.952) |
| Antibody | |||
| IgG | 3896 | 1092, 28.03% | 18.655 (9.735; 35.746) |
| IgG/IgM | 753 | 12, 1.59% | 1.061 (0.450; 2.500) |
| IgM | 599 | 9, 1.50% | REFERENCE |
| Diagnostic procedure | |||
| ELISA | 4085 | 1101, 26.95% | REFERENCE |
| HAT | 298 | 2, 0.67% | 0.025 (0.006; 0.099) |
| PRT | 455 | 10, 2.20% | 0.082 (0.044; 0.151) |
| ICT | 410 | 0, - | 0.005 (0.001; 0.072) |
| Pathogen | Tested (N) | OROV Positive (n/N, %) | Arbovirus Positive (n/N, %) | Risk Ratio (95%CI) |
|---|---|---|---|---|
| Virology | ||||
| DENV | 27,660 | 152, 0.55% | 3772, 13.64% | 24.816 (21.119; 29.159) |
| ZIKV | 2758 | 29, 1.05% | 229, 8.30% | 7.900 (5.386; 11.578) |
| CHKV | 2924 | 29, 0.99% | 64, 2.19% | 2.207 (1.427; 3.412) |
| Serology | ||||
| DENV | 20,269 | 206, 1.02% | 4868, 24.02% | 23.631 (20.584; 27.129) |
| ZIKV | 708 | 2, 0.28% | 99, 13.98% | 49.500 (12.256; 199.921) |
| CHKV | 1379 | 29, 2.10% | 90, 6.53% | 3.103 (2.056; 4.685) |
| Testing | No. of Series | No. of Observations | No. of Events | Detection Rate (%, 95%CI) | I2 (95%CI) | Tau2 | Q | p-Value |
|---|---|---|---|---|---|---|---|---|
| Virological assay | ||||||||
| Overall | 49 | 33,156 | 553 | 0.45 (0.16 to 1.23) | 94.9% (85.7 to 93.9) | 9.647 | 942.13 | <0.001 |
| Virus isolation | 8 | 1420 | 146 | 13.91 (5.66 to 30.34) | 95.3% | 1.875 | 147.87 | |
| RT-qPCR | 27 | 10,761 | 18 | 0.80 (0.31 to 2.06) | 93.5% | 5.585 | 402.38 | |
| RT-qPCR (on IFAT) | 13 | 20,880 | 366 | 0.03 (0.01 to 0.16) | 0.0% | 1.115 | 0.359 | |
| RT-qPCR (on MNT) | 1 | 95 | 23 | 24.21 (16.65 to 33.81) | - | - | - | |
| Serology, IgM only | ||||||||
| Overall | 12 | 13,037 | 755 | 12.45 (3.28 to 37.39) | 98.5% (98.1 to 98.8) | 6.310 | 726.71 | <0.001 |
| ELISA | 4 | 1404 | 394 | 34.83 (20.42 to 52.69) | 97.4% | 0.539 | 114.57 | |
| ELISA (on HAT) | 8 | 11633 | 361 | 6.61 (0.95 to 34.27) | 98.8% | 8.124 | 596.32 | |
| Serology, IgM/IgG | ||||||||
| Overall | 14 | 11,411 | 872 | 12.21 (4.96 to 27.09) | 99.0% (98.8 to 99.2) | 3.318 | 1337.11 | <0.001 |
| HAT | 9 | 3194 | 627 | 24.66 (10.97 to 46.51) | 98.7% | 2.143 | 629.52 | |
| PRT | 4 | 1610 | 177 | 4.31 (1.12 to 15.14) | 90.6% | 1.501 | 31.90 | |
| IFAT | 1 | 6607 | 68 | 1.03 (0.81 to 1.30) | - | - | ||
| Testing | No. of Series | No. of Observations | No. of Events | Detection Rate (%, 95%CI) | I2 (95%CI) | Tau2 | Q | p-Value |
|---|---|---|---|---|---|---|---|---|
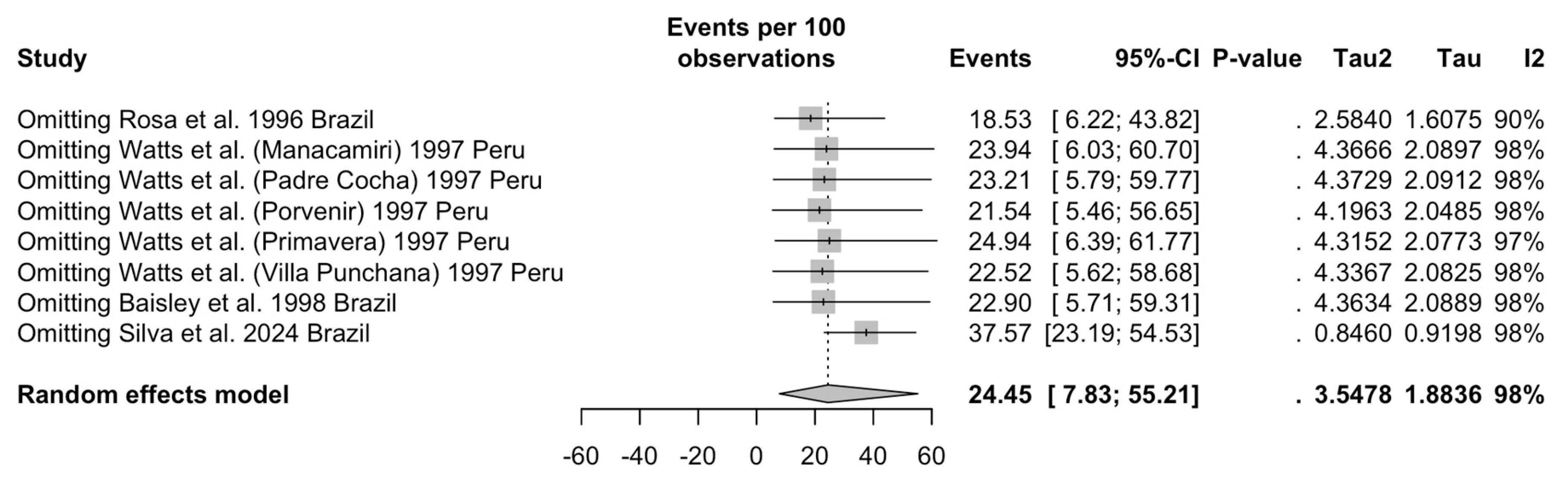
| Serology, IgG | ||||||||
| Overall | 8 | 3344 | 1083 | 24.45 (7.83 to 55.21) | 97.6% (96.5 to 98.3) | 3.548 | 287.63 | <0.001 |
| ELISA | 2 | 1616 | 338 | 59.82 (22.71 to 88.29) | 99.5% | 1.357 | 198.31 | |
| ELISA (on HAT) | 5 | 1523 | 635 | 29.31 (21.91 to 37.99) | 92.6% | 0.176 | 54.26 | |
| ICT | 1 | 205 | 0 | 0.0 (0.00 to 100) | - | - | - | |
| Serology, IgM/IgG | ||||||||
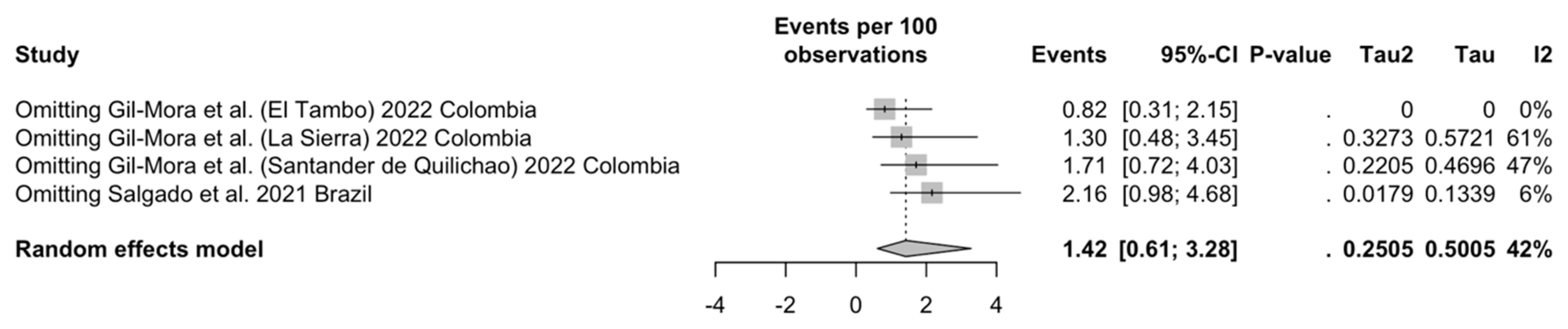
| Overall | 4 | 753 | 12 | 1.42 (0.61 to 3.28) | 41.9% (0.0 to 80.5) | 0.251 | 5.17 | 0.160 |
| HAT | 1 | 0.67 (0.17 to 2.64) | - | - | - | |||
| PRT | 3 | 2.16 (0.99 to 4.68) | 6.2% | 0.018 | 2.13 | |||
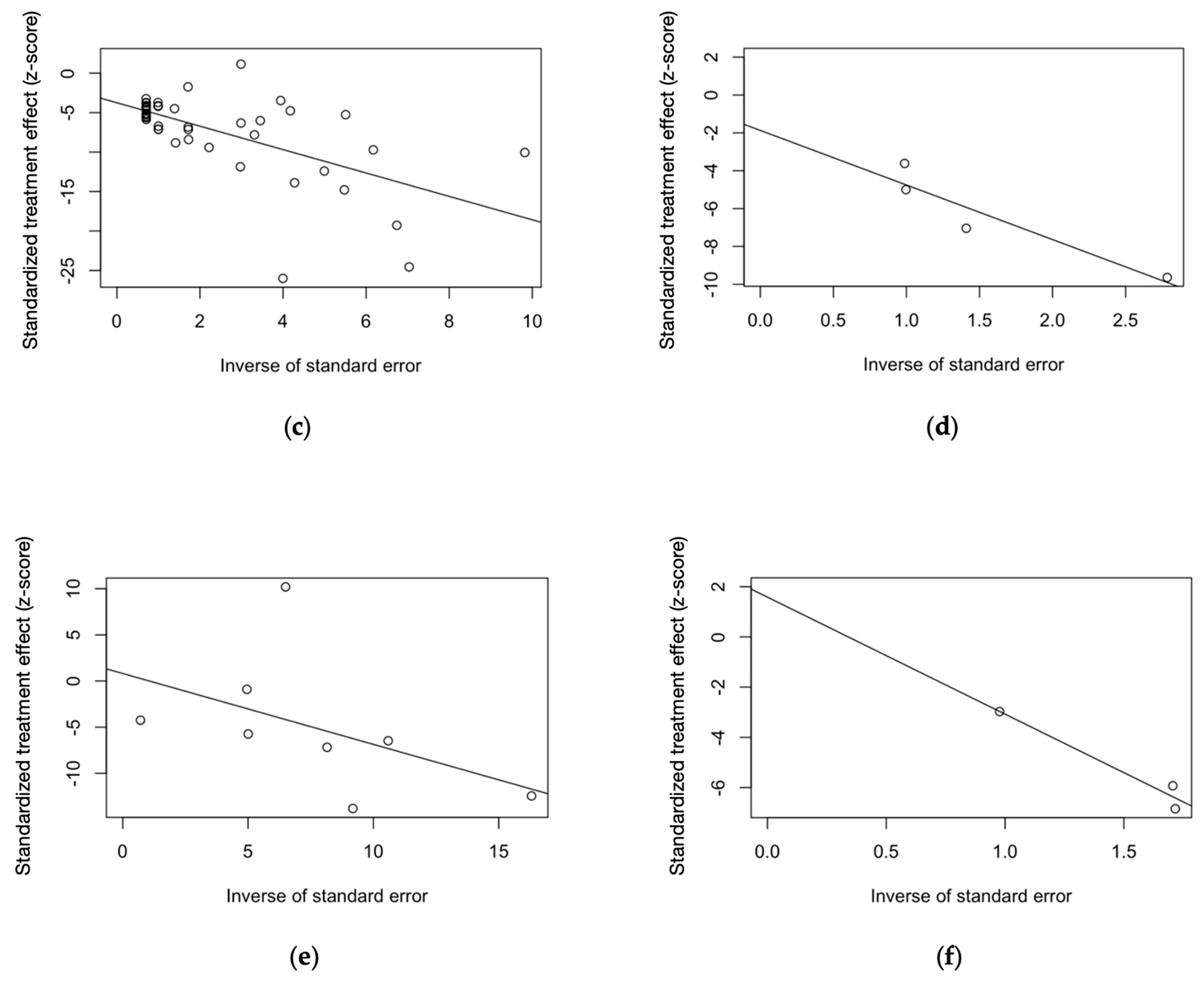
| Finding | t | df | Bias (SE) | Tau2 | p-Value |
|---|---|---|---|---|---|
| Symptomatic individuals | |||||
| Virological assay | −4.37 | 47 | −3.745 (0.857) | 16.964 | <0.001 |
| Serology, IgM only | −0.84 | 10 | −4.607 (5.466) | 67.851 | 0.419 |
| Serology, IgM + IgG | 0.20 | 12 | 1.137 (5.796) | 111.413 | 0.848 |
| CSF findings | 1.03 | 1 | 1.587 (1.543) | 0.377 | 0.491 |
| General population | |||||
| Serology, IgG only | 0.16 | 6 | 0.817 (5.091) | 50.087 | 0.878 |
| Serology, IgM + IgG | −1.42 | 2 | −1.871 (1.318) | 1.287 | 0.292 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Riccò, M.; Corrado, S.; Bottazzoli, M.; Marchesi, F.; Gili, R.; Bianchi, F.P.; Frisicale, E.M.; Guicciardi, S.; Fiacchini, D.; Tafuri, S.; et al. (Re-)Emergence of Oropouche Virus (OROV) Infections: Systematic Review and Meta-Analysis of Observational Studies. Viruses 2024, 16, 1498. https://doi.org/10.3390/v16091498
Riccò M, Corrado S, Bottazzoli M, Marchesi F, Gili R, Bianchi FP, Frisicale EM, Guicciardi S, Fiacchini D, Tafuri S, et al. (Re-)Emergence of Oropouche Virus (OROV) Infections: Systematic Review and Meta-Analysis of Observational Studies. Viruses. 2024; 16(9):1498. https://doi.org/10.3390/v16091498
Chicago/Turabian StyleRiccò, Matteo, Silvia Corrado, Marco Bottazzoli, Federico Marchesi, Renata Gili, Francesco Paolo Bianchi, Emanuela Maria Frisicale, Stefano Guicciardi, Daniel Fiacchini, Silvio Tafuri, and et al. 2024. "(Re-)Emergence of Oropouche Virus (OROV) Infections: Systematic Review and Meta-Analysis of Observational Studies" Viruses 16, no. 9: 1498. https://doi.org/10.3390/v16091498