
Effects of Two Straw Return Methods Coupled with Raising Ducks in Paddy Fields on Stem Lodging Characteristics

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experiment Site
2.2. Experiment Design and Treatment
- RNN—no straw return and no ducks in the paddy field;
- RND—no straw return and ducks in the paddy field;
- RSN—direct straw return and no ducks in paddy field;
- RSD—direct straw return and ducks in paddy field;
- RBN—straw carbon and no ducks in paddy field;
- RBD—straw carbon and ducks in paddy field.
2.3. Straw Processing
2.4. Experiment Details
2.4.1. Crop Management
2.4.2. Straw and Fertilization Management
2.4.3. Duck Management
2.5. Parameters Measured
2.5.1. Yield and the Compounds of Yield
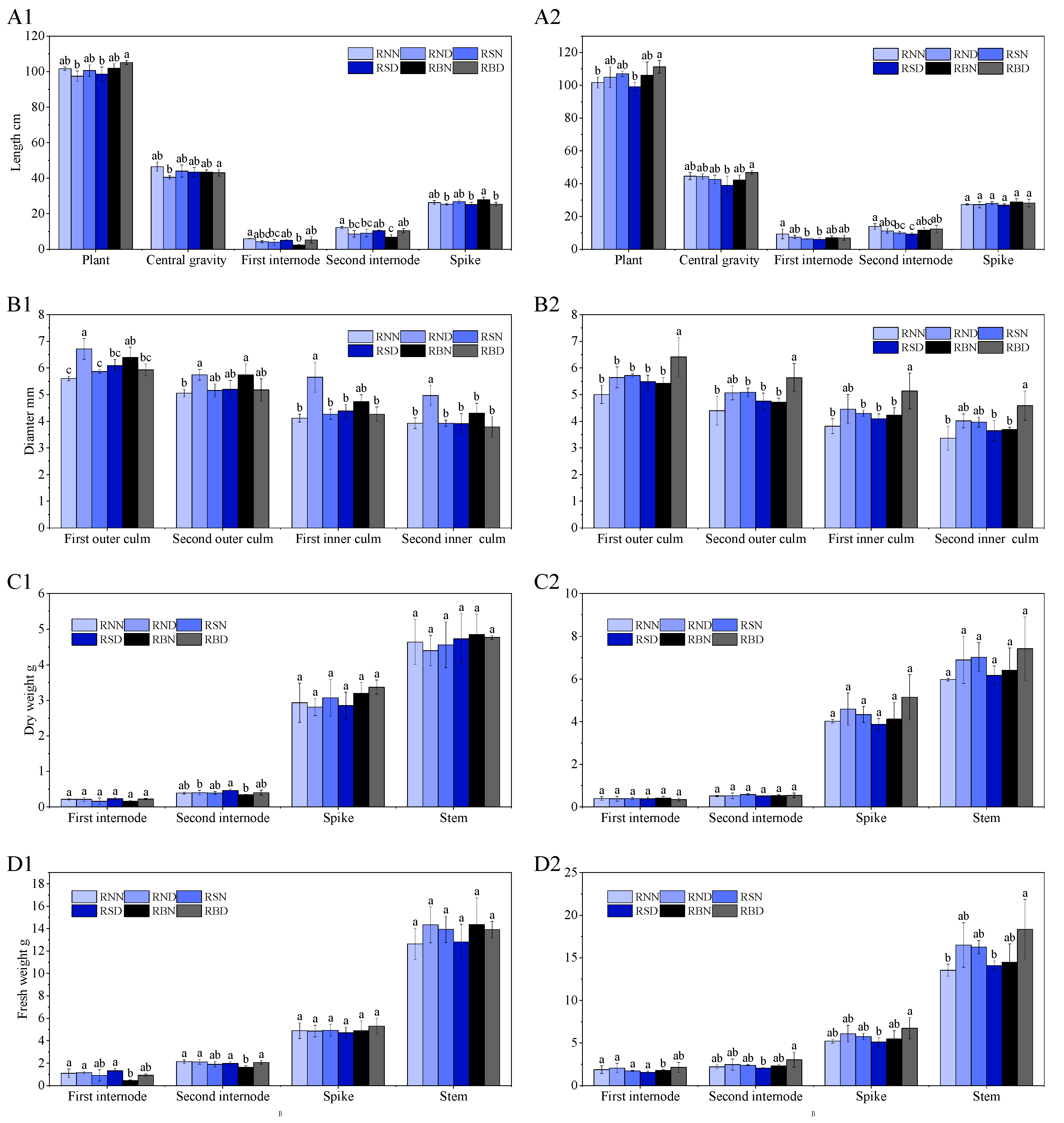
2.5.2. Culm Morphology
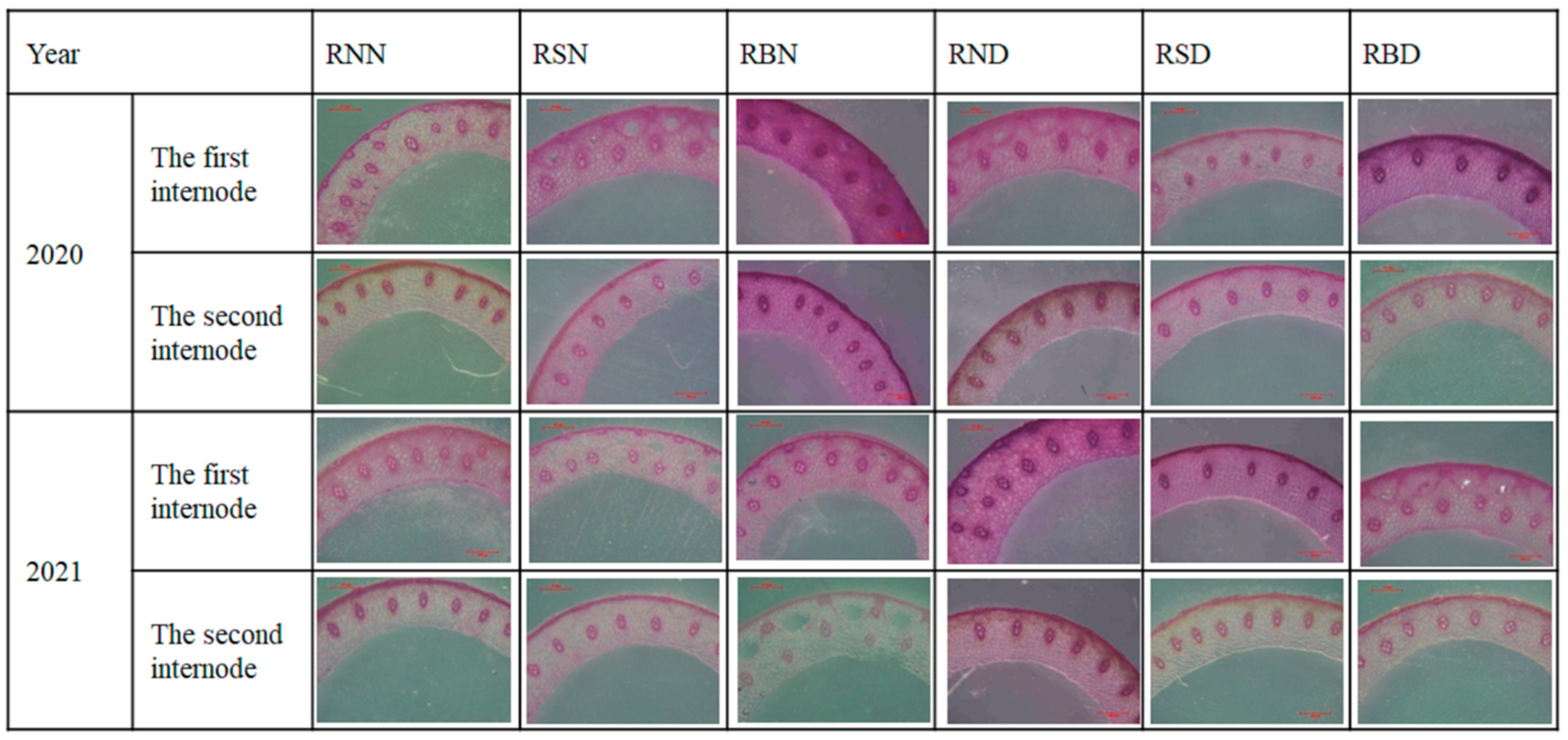
2.5.3. Culm Anatomy
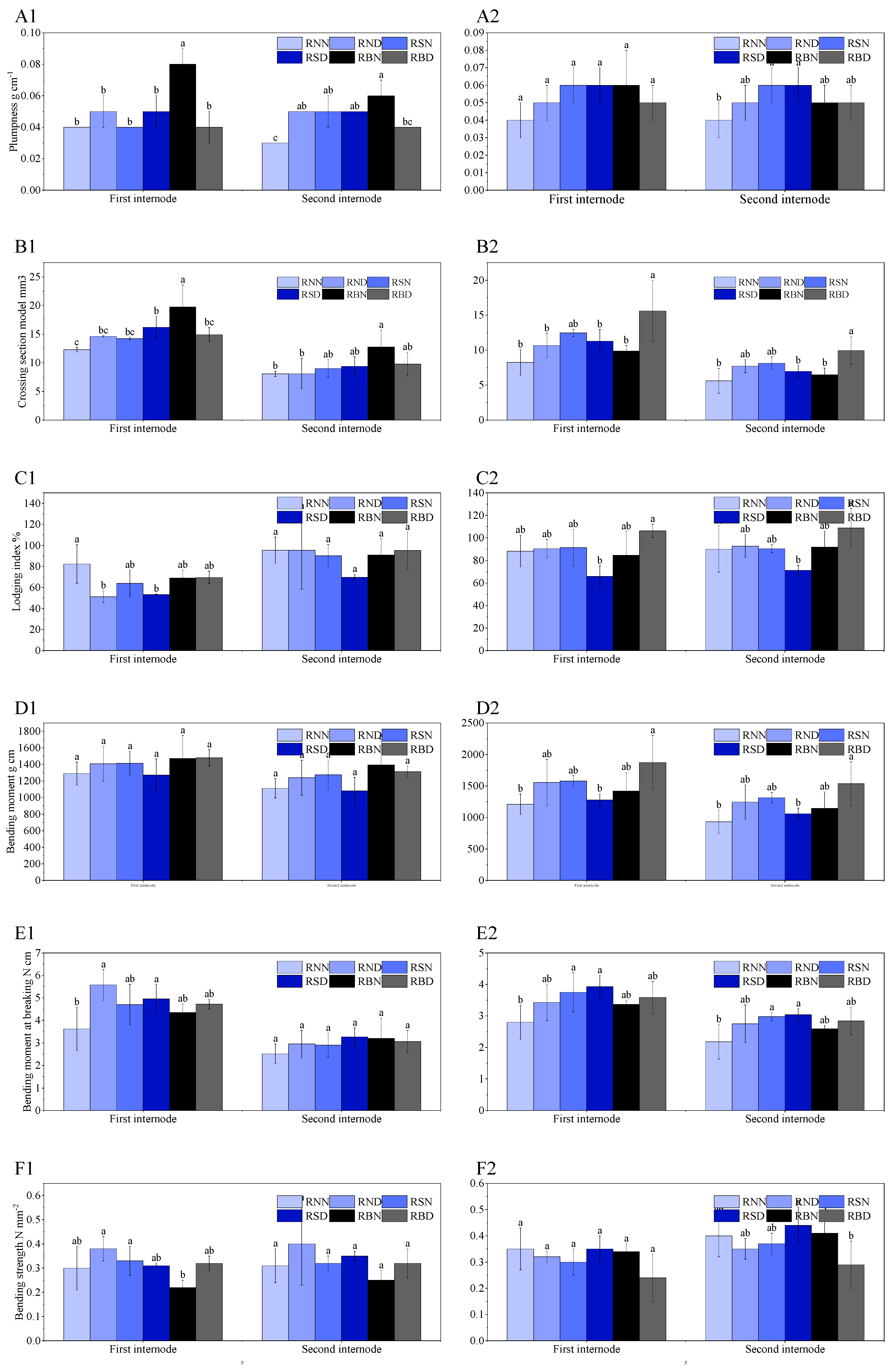
2.5.4. Lodging Resistance
- Bending moment (WP, g cm) of each internode = Length from the basal of the internode to panicle (cm) × Fresh plant weight of plant part from the basal of the internode to panicle (g).
- Bending moment at breaking (M, N cm) of each internode = Break load (N) at each internode × Distance between fulcra (cm).
- Lodging index (LI, %) = Bending Load of each internode/The bending load of each internode.
- Crossing section model(Z, mm3) = π × D3/32 × (1 − α4) where D is the outer diameter in an oval cross-section and α is the ratio of the inner diameter and outer diameter in an oval cross-section.
- Bending strength (BS, N mm−2) of each internode = Bending moment at breaking/Crossing section model.
2.6. Statistical Analysis
3. Results
3.1. Yield and Yield Components
3.2. Culm Morphology
3.3. Culm Anatomy
3.4. Lodging Resistance
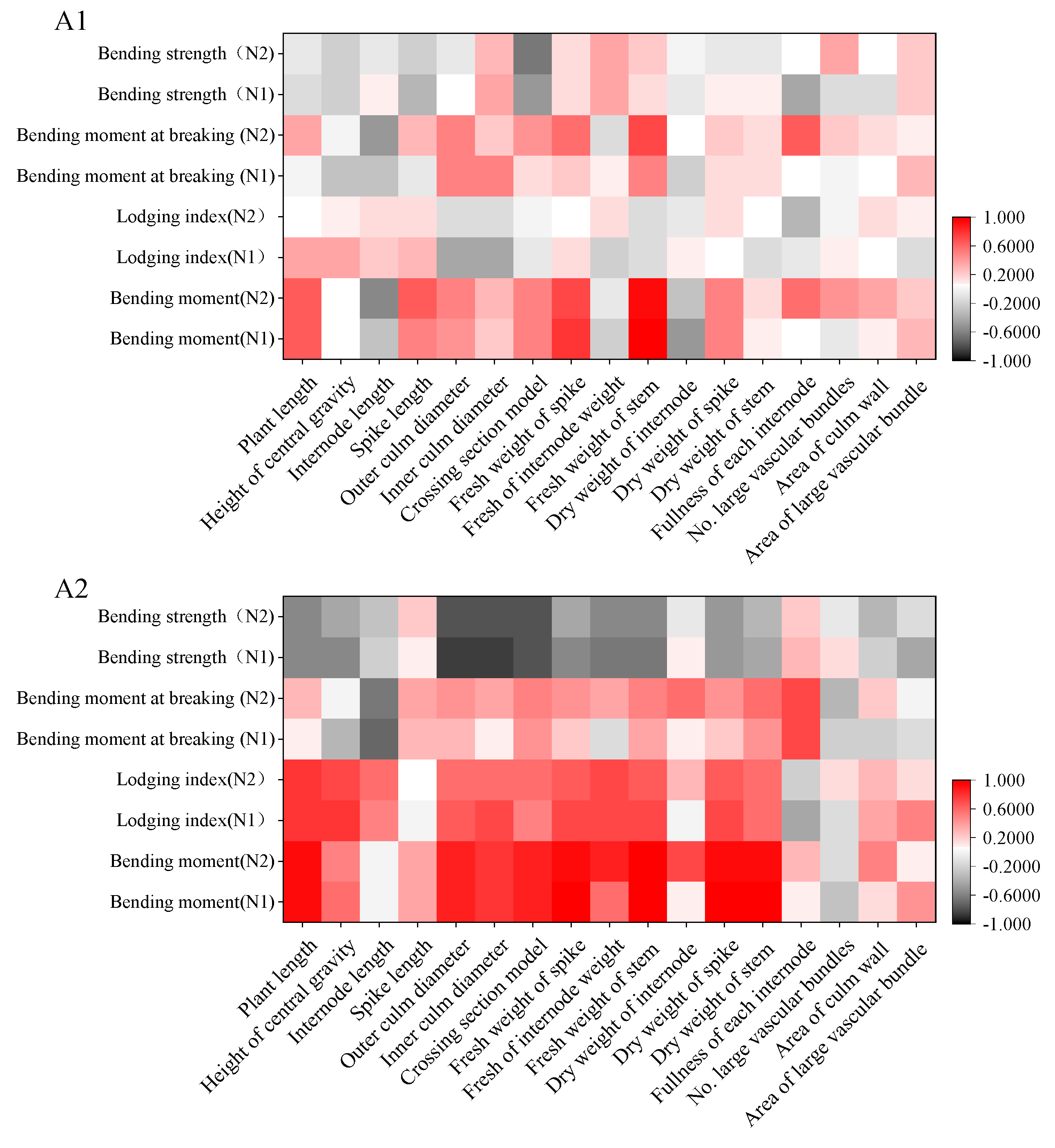
3.5. Partial Correlation Analysis of Factor Affecting Lodging Resistance
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Salassi, M.E.; Deliberto, M.A.; Linscombe, S.D.; Wilson, C.E., Jr.; Walker, T.W.; McCauley, G.N.; Blouin, D.C. Impact of Harvest Lodging on Rough Rice Milling Yield and Market Price. Agron. J. 2013, 105, 1860–1867. [Google Scholar] [CrossRef] [Green Version]
- Kazuo, T. Eco-physiological study of root lodging tolerance indirect-seeds rice cultivars. JARQ 1997, 31, 155–162. Available online: https://www.jircas.go.jp/sites/default/files/publication/jarq/31-3-155-162_0.pdf (accessed on 31 July 2022).
- Zhu, C.; Ziska, L.H.; Sakai, H.; Zhu, J.; Hasegawa, T. Vulnerability of lodging risk to elevated CO2 and increased soil temperature differs between rice cultivars. Eur. J. Agron. 2013, 46, 20–24. [Google Scholar] [CrossRef]
- Thakur, A.K.; Rath, S.; Patil, D.U.; Kumar, A. Effects on rice plant morphology and physiology of water and associated management practices of the system of rice intensification and their implications for crop performance. Paddy Water Environ. 2010, 9, 13–24. [Google Scholar] [CrossRef]
- Han, L.J.; Yan, Q.J.; Liu, X.Y.; Hu, J.Y. Straw resources and their utilization in china. Trans CSAE 2002, 18, 87–91. [Google Scholar]
- Li, H.; Dai, M.; Dai, S.; Dong, X. Current status and environment impact of direct straw return in China’s cropland—A review. Ecotoxicol. Environ. Saf. 2018, 159, 293–300. [Google Scholar] [CrossRef] [PubMed]
- Ren, J.; Yu, P.; Xu, X. Straw Utilization in China—Status and Recommendations. Sustainability 2019, 11, 1762. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Lu, M.; Cui, J.; Li, B.; Fang, C. Effects of Straw Carbon Input on Carbon Dynamics in Agricultural Soils: A Meta-Analysis. Glob. Chang. Biol. 2014, 20, 1366–1381. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Qi, Y.; Wang, J.; Jiang, Y.; Geng, M. Synergistic effects of crop residue and microbial inoculant on soil properties and soil disease resistance in a Chinese Mollisol. Sci. Rep. 2021, 11, 24225. [Google Scholar] [CrossRef] [PubMed]
- Zhang, A.; Bian, R.; Pan, G.; Cui, L.; Hussain, Q.; Li, L.; Zheng, J.; Zheng, J.; Zhang, X.; Han, X.; et al. Effects of biochar amendment on soil quality, crop yield and greenhouse gas emission in a Chinese rice paddy: A field study of 2 consecutive rice growing cycles. Field Crop. Res. 2012, 127, 153–160. [Google Scholar] [CrossRef]
- Zhang, Q.; Song, Y.; Wu, Z.; Yan, X.; Gunina, A.; Kuzyakov, Y.; Xiong, Z. Effects of six-year biochar amendment on soil aggregation, crop growth, and nitrogen and phosphorus use efficiencies in a rice-wheat rotation. J. Clean. Prod. 2019, 242, 118435. [Google Scholar] [CrossRef]
- Huang, D.; Liu, L.; Zeng, G.; Xu, P.; Huang, C.; Deng, L.; Wang, R.; Wan, J. The effects of rice straw biochar on indigenous microbial community and enzymes activity in heavy metal-contaminated sediment. Chemosphere 2017, 174, 545–553. [Google Scholar] [CrossRef] [PubMed]
- Karam, D.S.; Nagabovanalli, P.; Rajoo, K.S.; Ishak, C.F.; Abdu, A.; Rosli, Z.; Muharam, F.M.; Zulperi, D. An overview on the preparation of rice husk biochar, factors affecting its properties, and its agriculture application. J. Saudi Soc. Agric. Sci. 2021, 21, 149–159. [Google Scholar] [CrossRef]
- Bonanomi, G.; Ippolito, F.; Scala, F. A “black” future for plant pathology? Biochar as a new soil amendment for controlling plant diseases. J. Plant Pathol. 2015, 97, 223–234. [Google Scholar] [CrossRef]
- Hou, X.; Meng, L.Q.; Li, L.; Pan, G.; Li, B. Biochar amendment to soils impairs developmental and reproductive performances of a major rice pest Nilaparvata lugens (Homopera: Delphacidae). J. Appl. Èntomol. 2015, 139, 727–733. [Google Scholar] [CrossRef]
- Xia, Y.; Li, F.; Song, Q.; Zhang, J.; Liu, X.; Sun, L. Effects of Biochar on Morphological Characters of Rice Stem. IOP Conf. Series: Mater. Sci. Eng. 2019, 493, 012129. [Google Scholar] [CrossRef]
- Zheng, H.; Huang, H.; Li, D.; Li, X.; Fu, Z.; Chen, C. Assessment of nomadic rice-duck complex ecosystem on energy and economy. Ecol. Process. 2014, 3, 52. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Li, R.; Liu, S.; Zhang, J.; Luo, H.; Qiu, S. Rice-duck co-culture benefits grain 2-acetyl-1-pyrroline accumulation and quality and yield enhancement of fragrant rice. Crop, J. 2019, 7, 419–430. [Google Scholar] [CrossRef]
- Zhang, J.-E.; Quan, G.-M.; Huang, Z.-X.; Luo, S.-M.; Ouyang, Y. Evidence of Duck Activity Induced Anatomical Structure Change and Lodging Resistance of Rice Plant. Agroecol. Sustain. Food Syst. 2013, 37, 975–984. [Google Scholar] [CrossRef]
- Tian, B.H.; Liu, L.Y.; Zhang, L.X.; Song, S.X.; Wang, J.G.; Wu, L.F.; Li, H.J. Characterization of culm morphology, anatomy and chemical composition of foxtail millet cultivars differing in lodging resistance. J. Agric. Sci. 2015, 153, 1437–1448. [Google Scholar] [CrossRef]
- Sreeja, R.; Arul, L.; Kannan, B.J.R.; Nirmala, K.A.; Subramanian, A. Culm anatomy revealed association of vascular bundle number and silicon content with lodging behavior in Kodo millet (Paspalum scrobiculatum L.). Indian J. Exp. Biol. 2019, 57, 893–898. [Google Scholar]
- Liang, Y.G.; Chen, Y.S.; Chen, L.; Ma, W.W.; Meng, X.J.; Huang, H.; Yu, Z.J. Effect of rice-fish-chicken ridge cultivation on stem lodging resistance, panicle traits, and yield of rice. Chin. J. Eco-Agri. 2021, 29, 379–388. [Google Scholar] [CrossRef]
- Zhang, J.; Li, G.; Song, Y.; Liu, Z.; Yang, C.; Tang, S.; Zheng, C.; Wang, S.; Ding, Y. Lodging resistance characteristics of high-yielding rice populations. Field Crop. Res. 2014, 161, 64–74. [Google Scholar] [CrossRef]
- Ookawa, T.; Hobo, T.; Yano, M.; Murata, K.; Ando, T.; Miura, H.; Asano, K.; Ochiai, Y.; Ikeda, M.; Nishitani, R.; et al. New approach for rice improvement using a pleiotropic QTL gene for lodging resistance and yield. Nat. Commun. 2010, 1, 132. [Google Scholar] [CrossRef] [Green Version]
- Yu, S.; Ouyang, Y.; Zhang, Q.; Peng, G.; Xu, D.; Jin, Q. Effects of rice-duck farming system on Oryza sativa growth and its yield. Ying Yong Sheng Tai Xue Bao = J. Appl. Ecol. 2005, 16, 1252–1256. [Google Scholar]
- Wang, J.; Wang, X.; Xu, M.; Feng, G.; Zhang, W.; Lu, C. Crop yield and soil organic matter after long-term straw return to soil in China. Nutr. Cycl. Agroecosystems 2015, 102, 371–381. [Google Scholar] [CrossRef]
- Huang, M.; Fan, L.; Jiang, L.-G.; Yang, S.-Y.; Zou, Y.-B.; Uphoff, N. Continuous applications of biochar to rice: Effects on grain yield and yield attributes. J. Integr. Agric. 2019, 18, 563–570. [Google Scholar] [CrossRef]
- Zhang, Z.C.; Chen, J.Y.; Zhang, Y.M. Effect of returning all wheat straw to field on soil fertility and rice growth in hilly land. South Chin. Agr. 2021, 15, 43–46. [Google Scholar] [CrossRef]
- Navabi, A.; Iqbal, M.; Strenzke, K.; Spaner, D. The relationship between lodging and plant height in a diverse wheat population. Can. J. Plant Sci. 2006, 86, 723–726. [Google Scholar] [CrossRef]
- Xiang, D.B.; Zhao, G.; Wan, Y.; Tan, M.L.; Song, C.; Song, Y. Effect of planting density on lodging-related morphology, lodging rate, and yield of tartary buckwheat (Fagopyrum tataricum). Plant Prod. Sci. 2016, 19, 479–488. [Google Scholar] [CrossRef] [Green Version]
- Xiang, D.; Song, Y.; Wu, Q.; Ma, C.; Zhao, J.; Wan, Y.; Zhao, G. Relationship between stem characteristics and lodging resistance of Tartary buckwheat (Fagopyrum tataricum). Plant Prod. Sci. 2019, 22, 202–210. [Google Scholar] [CrossRef] [Green Version]
- Stubbs, C.J.; Oduntan, Y.A.; Keep, T.R.; Noble, S.D.; Robertson, D.J. The effect of plant weight on estimations of stalk lodging resistance. Plant Methods 2020, 16, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.Y.; Jin, Z.; Wen, J.; Song, Y.Q.; Kong, A.U.; Xu, Y.; Cui, H.G.; Xin, M.J.; Zhang, B.H.; Liu, C.H. Effects of Rice-duck Farming on Soil Nutrient Content and Stem Lodging Resistance of Rice. J. Shenyang Agr. Uni. 2019, 50, 365–370. [Google Scholar]
- Meng, X.J.; Liang, Y.G.; Zou, Z.Y.; Wang, R.; Su, Y.T.; Gong, X.S.; Huang, H.; Chen, C. Effects of biochar application on rice stem and lodging traits under rice duck co-breeding. Chin. J. Ecol. 2021, 40, 3125–3134. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Treatment | Panicles (m−2) | Spike Number/Panicle | 1000-Grain Weight (g) | Filled Spike (%) | Yield (kg ha−1) |
|---|---|---|---|---|---|---|
| 2020 | RNN | 268.52 ± 16.04 abc | 200.75 ± 34.30 a | 25.32 ± 0.10 bc | 64.6 ± 4.28 a | 6866.27 ± 1213.9 a |
| RND | 259.26 ± 21.22 bc | 197.86 ± 13.58 a | 24.98 ± 0.30 bc | 62.87 ± 2.45 ab | 7014.24 ± 216.11 a | |
| RSN | 254.63 ± 8.02 c | 179.56 ± 20.79 a | 24.76 ± 0.51 c | 65.06 ± 3.77 a | 6802.78 ± 1687.84 a | |
| RSD | 300.93 ± 28.91 a | 205.65 ± 49.59 a | 25.51 ± 0.19 b | 47.73 ± 4.38 c | 6910.19 ± 533.61 a | |
| RBN | 259.26 ± 8.02 bc | 211.05 ± 31.66 a | 25.58 ± 0.44 ab | 57.81 ± 16.33 abc | 6218.55 ± 705.97 a | |
| RBD | 291.67 ± 13.89 ab | 187.22 ± 15.51 a | 26.21 ± 0.42 a | 49.64 ± 3.48 bc | 6015.07 ± 688.32 a | |
| 2021 | RNN | 180.56 ± 0.00 b | 133.56 ± 11.83 a | 24.83 ± 0.50 ab | 80.26 ± 3.07 a | 5649.77 ± 268.2 d |
| RND | 203.7 ± 8.02 ab | 165.94 ± 24.11 a | 24.35 ± 0.09 b | 76.54 ± 7.22 a | 7443.73 ± 649.17 a | |
| RSN | 226.85 ± 21.22 a | 135.58 ± 6.68 a | 25.17 ± 0.54 a | 81.71 ± 3.00 a | 6096.30 ± 410.83 cd | |
| RSD | 203.70 ± 8.02 ab | 164.88 ± 32.68 a | 24.59 ± 0.37 ab | 75.67 ± 2.12 a | 6886.67 ± 225.11 ab | |
| RBN | 222.22 ± 0.00 a | 156.1 ± 21.76 a | 24.45 ± 0.46 ab | 82.04 ± 7.13 a | 6496.40 ± 381.81 bc | |
| RBD | 212.96 ± 21.22 a | 151.14 ± 17.71 a | 24.50 ± 0.07 ab | 78.00 ± 3.87 a | 5774.23 ± 275.91 cd | |
| F-value | ||||||
| Year(Y) | 153.089 ** | 27.927 ** | 35.932 ** | 100.593 ** | 1.002 | |
| Factor1(F1) | 3.756 | 1.163 | 0.002 | 10.574 ** | 1.675 | |
| Y × F1 | 6.422 * | 1.215 | 7.567 * | 1.133 | 1.499 | |
| Factor2(F2) | 5.689 ** | 0.112 | 2.157 | 1.532 | 2.511 | |
| Y × F2 | 0.622 | 0.057 | 7.838 ** | 2.632 | 0.291 | |
| F1 × F2 | 0.089 | 2.062 | 3.165 | 1.545 | 2.89 | |
| Y × F1 × F2 | 8.622 ** | 0.285 | 1.961 | 0.832 | 1.614 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gong, X.; Meng, X.; Zhang, Y.; Liang, Y.; Chen, C.; Huang, H.; Liao, X. Effects of Two Straw Return Methods Coupled with Raising Ducks in Paddy Fields on Stem Lodging Characteristics. Sustainability 2022, 14, 12984. https://doi.org/10.3390/su142012984
Gong X, Meng X, Zhang Y, Liang Y, Chen C, Huang H, Liao X. Effects of Two Straw Return Methods Coupled with Raising Ducks in Paddy Fields on Stem Lodging Characteristics. Sustainability. 2022; 14(20):12984. https://doi.org/10.3390/su142012984
Chicago/Turabian StyleGong, Xiangsheng, Xiangjie Meng, Ya Zhang, Yugang Liang, Can Chen, Huang Huang, and Xin Liao. 2022. "Effects of Two Straw Return Methods Coupled with Raising Ducks in Paddy Fields on Stem Lodging Characteristics" Sustainability 14, no. 20: 12984. https://doi.org/10.3390/su142012984