
Effects of Altitude, Plant Communities, and Canopies on the Thermal Comfort, Negative Air Ions, and Airborne Particles of Mountain Forests in Summer

Abstract
:1. Introduction
2. Materials and Methods
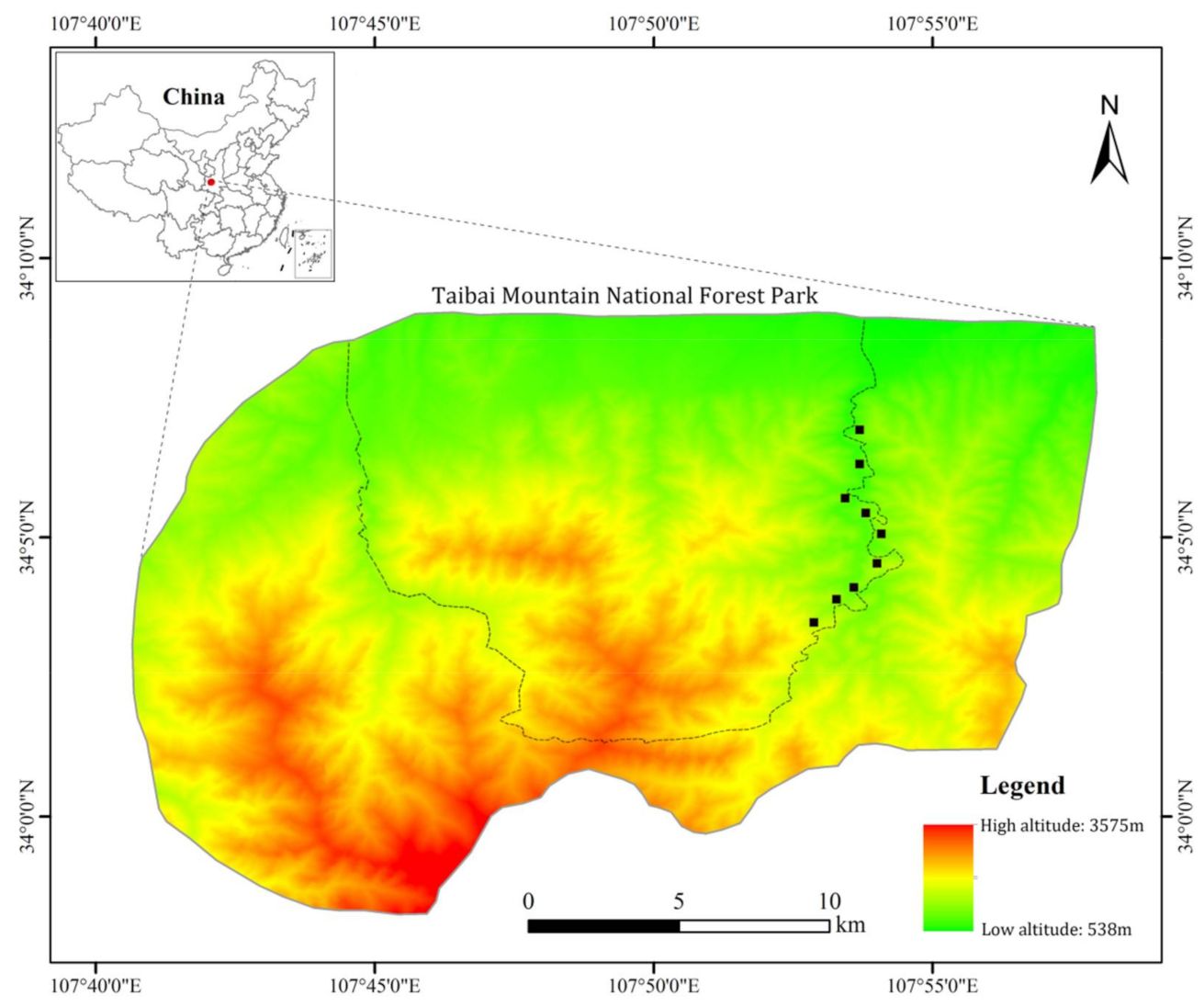
2.1. Study Area and Measurement Sites
2.2. Universal Thermal Climate Index (UTCI), Negative Air Ion (NAI), and Airborne Particulate Matter (PM2.5 and PM10) Level Measurements
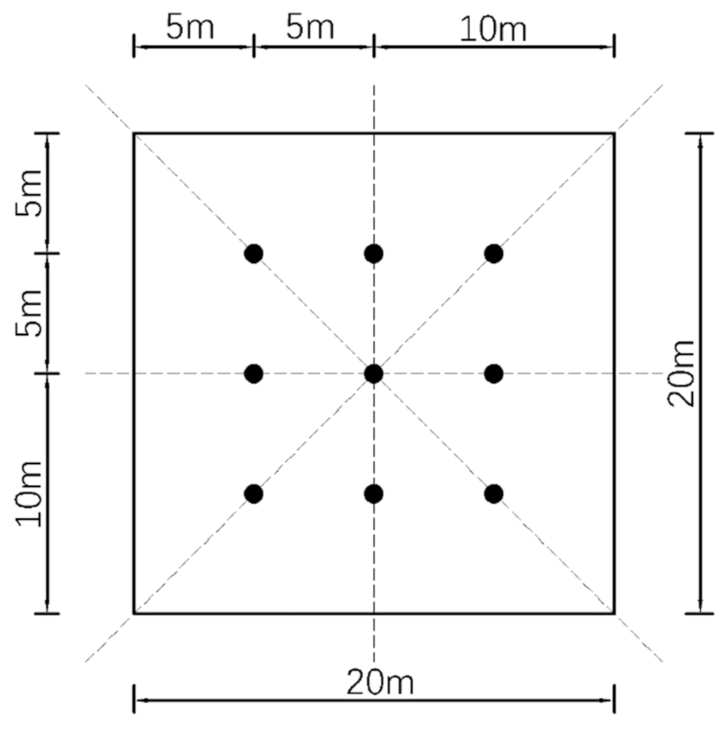
2.3. Measurements of Canopy Characteristics
2.4. Data Analysis
3. Results
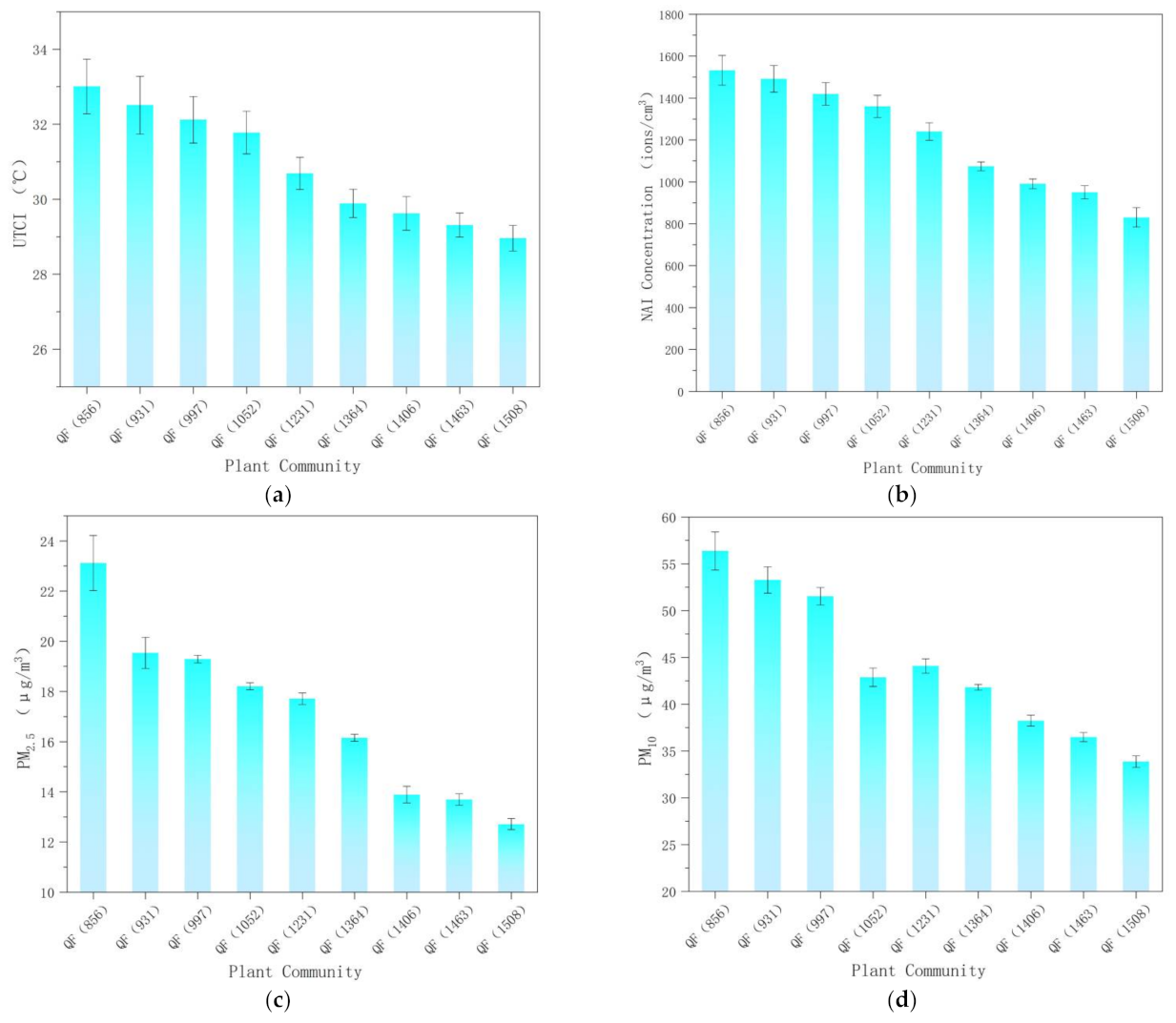
3.1. UTCI, NAI, PM2.5, and PM10 Levels of Quercus Variabilis Communities at Different Altitudes
3.1.1. Universal Thermal Climate Index (UTCI)
3.1.2. NAI Concentration
3.1.3. PM2.5 and PM10 Concentrations
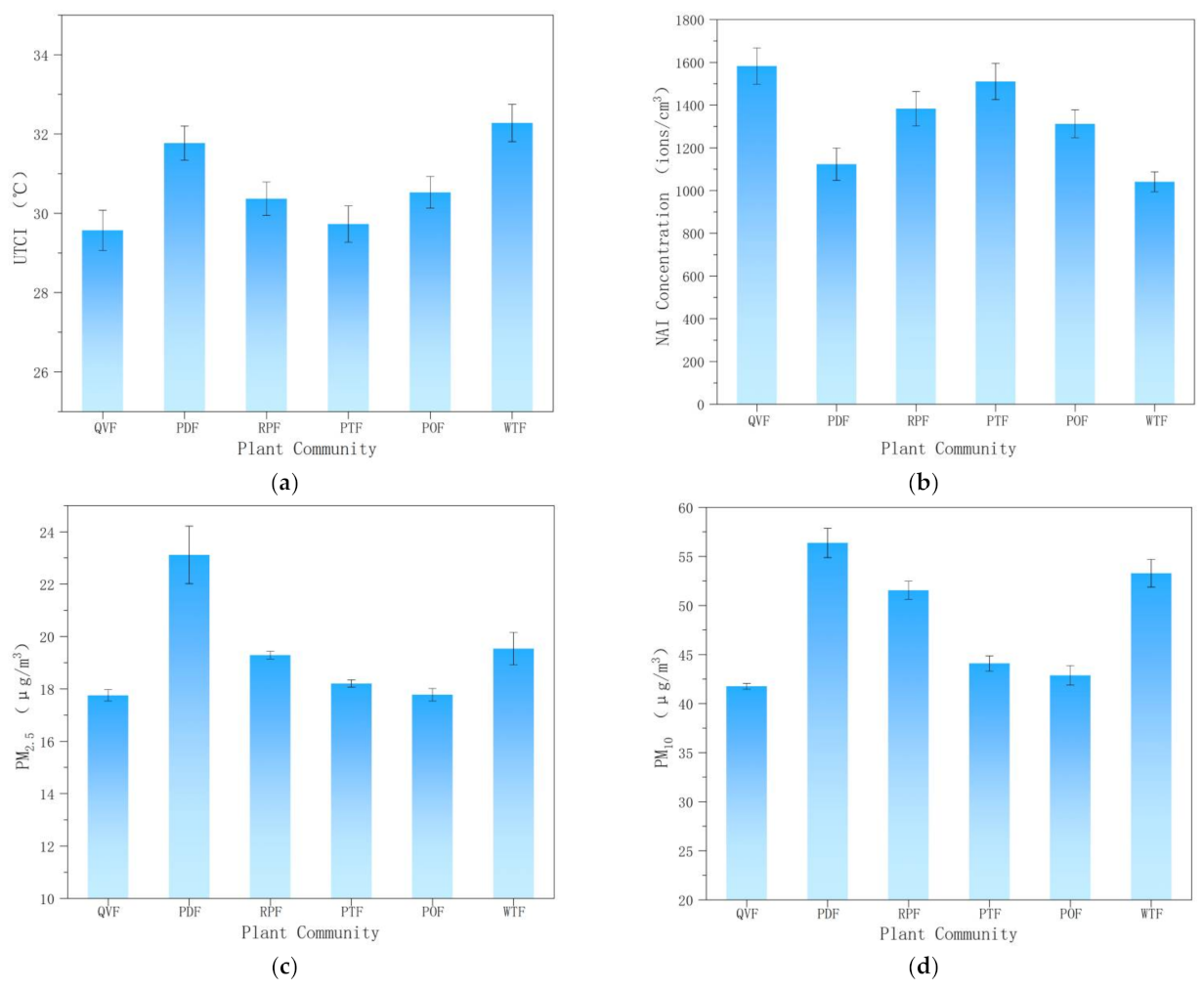
3.2. UTCI, NAI, PM2.5, and PM10 Levels of Different Plant Communities
3.2.1. Universal Thermal Climate Index (UTCI)
3.2.2. NAI Concentration
3.2.3. PM2.5 and PM10 Concentrations
3.3. Response of the UTCI, NAI, PM2.5, and PM10 Levels to Canopy Characteristics of the Plant Communities
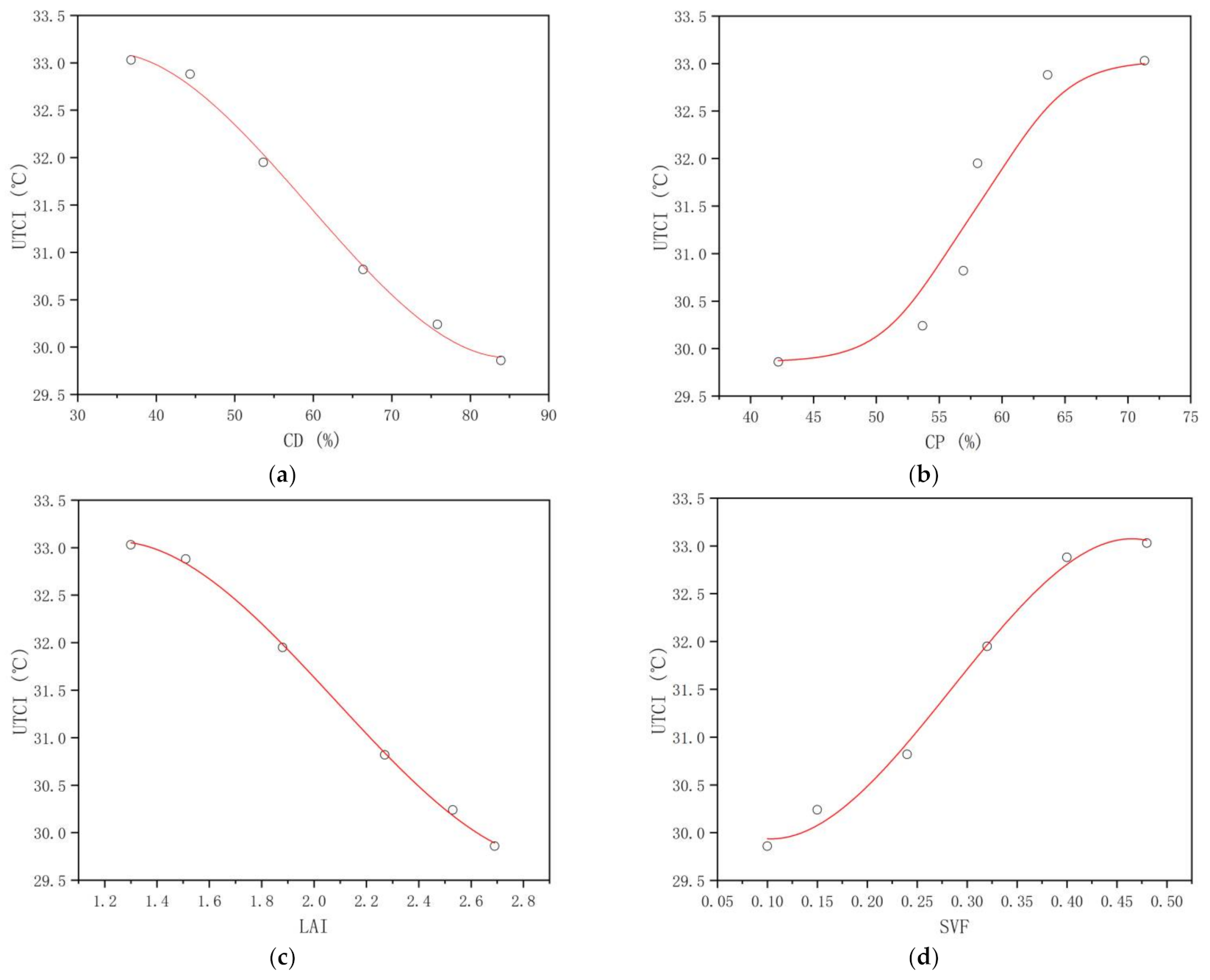
3.3.1. Universal Thermal Climate Index (UTCI)
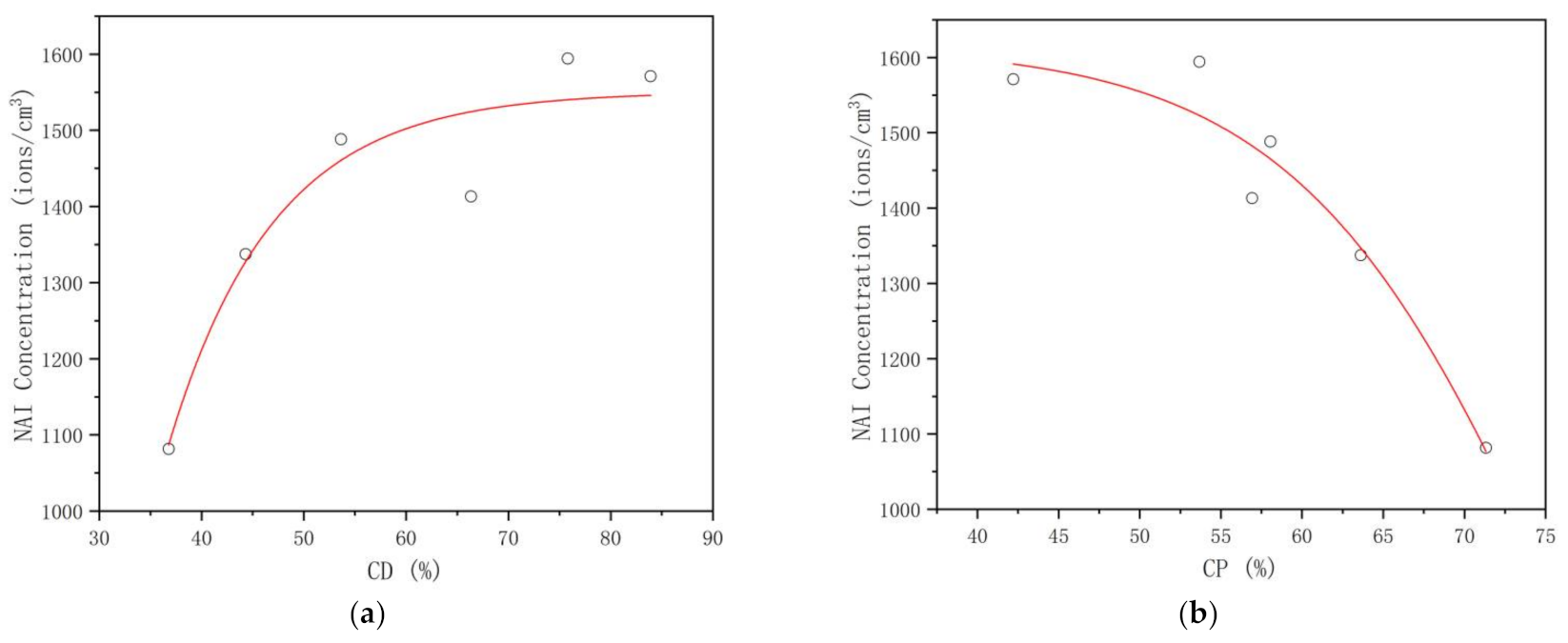
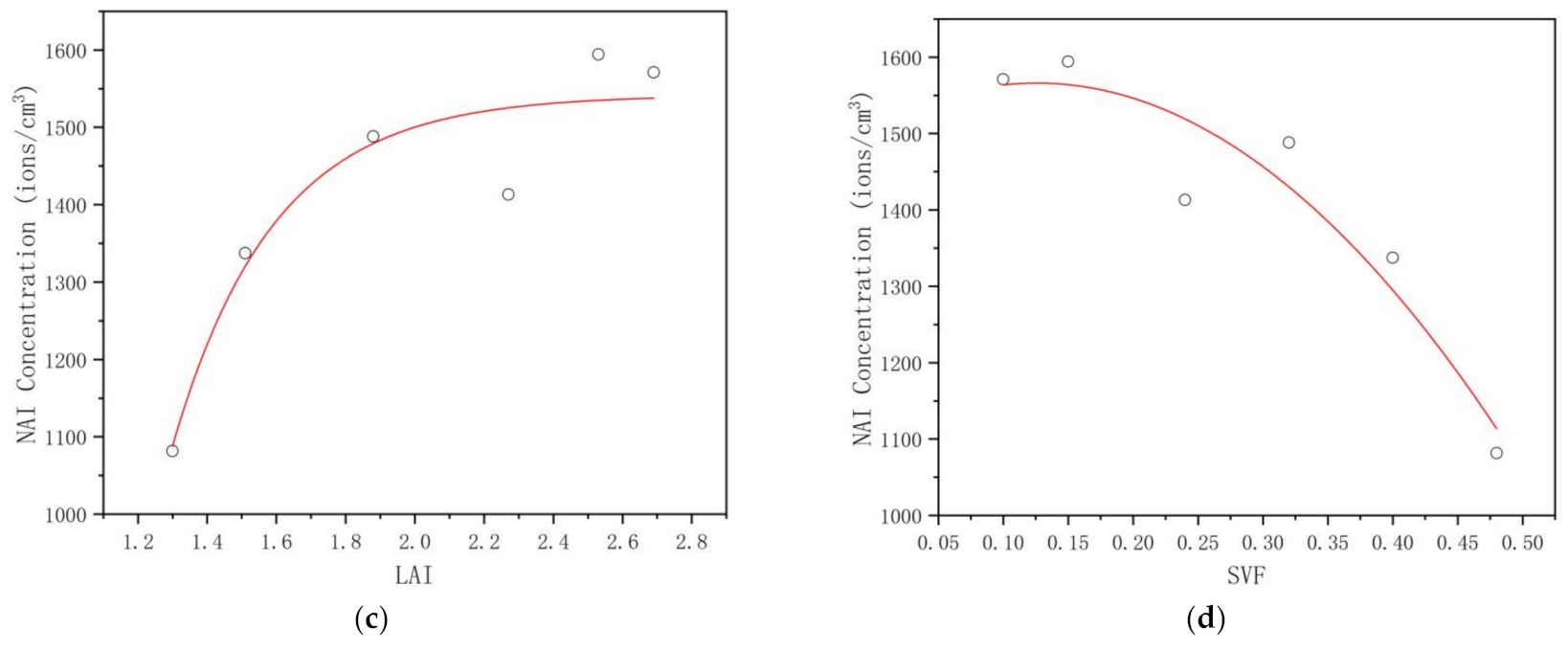
3.3.2. NAI Concentration
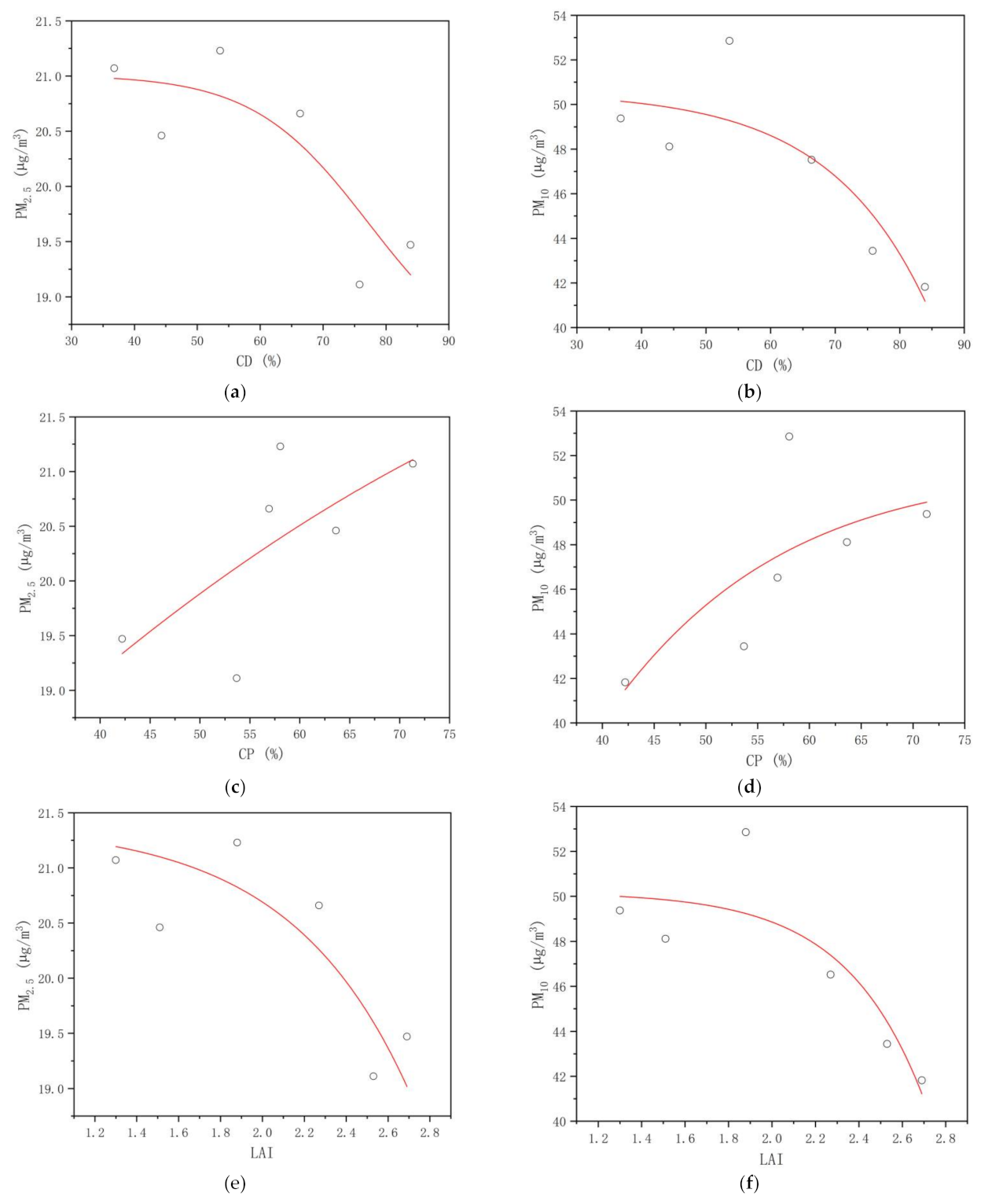
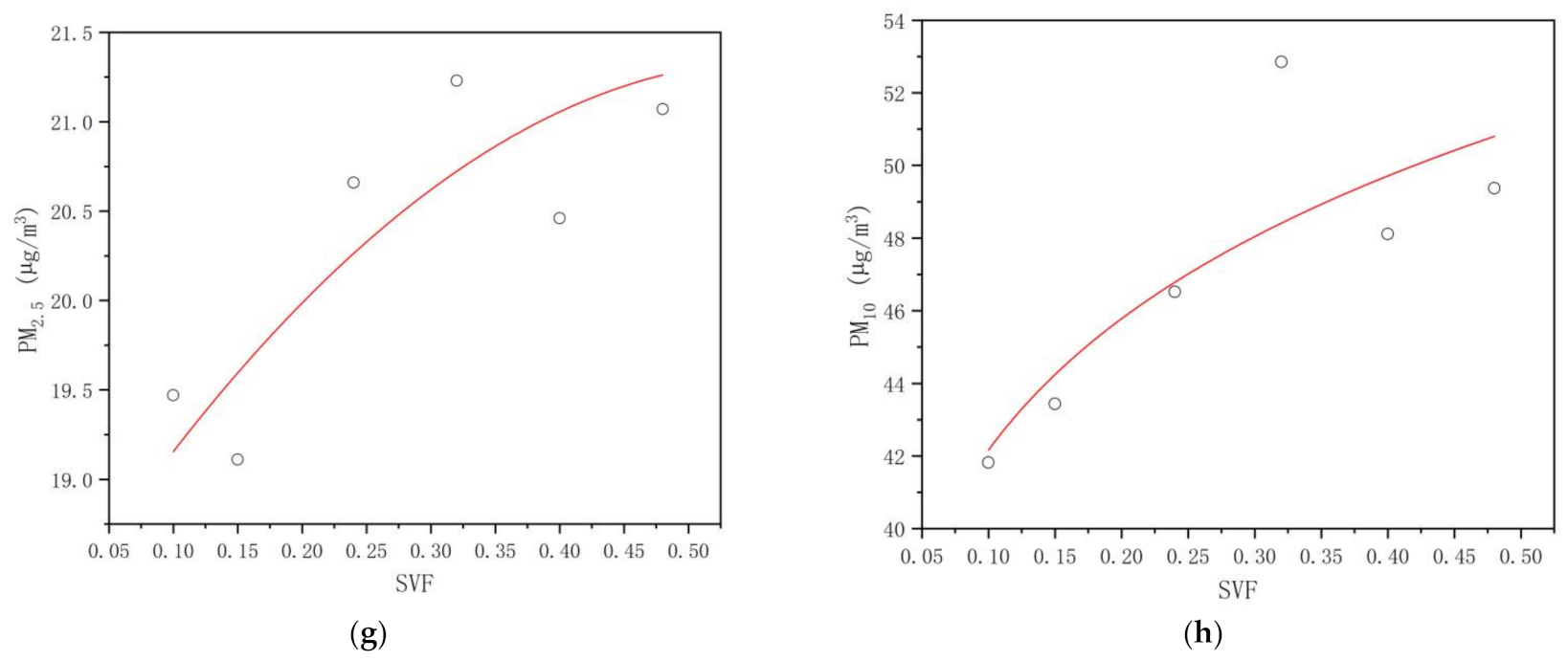
3.3.3. PM2.5 and PM10 Concentrations
4. Discussion
4.1. Effects of Altitude on the UTCI, NAI, PM2.5, and PM10 Levels of Plant Communities in the Mountain Forest
4.2. Effects of Plant Community Composition on the UTCI, NAI, PM2.5, and PM10 Levels in the Mountain Forest
4.3. Critical Thresholds of the Plant Community Canopy’s Regulating Effects on the UTCI, NAI, PM2.5, and PM10 Levels in the Mountain Forest
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kalnay, E.; Cai, M. Impact of urbanization and land-use change on climate. Nature 2003, 423, 528–531. [Google Scholar] [CrossRef] [PubMed]
- Dong, H.; Xue, M.; Xiao, Y.; Liu, Y. Do carbon emissions impact the health of residents? Considering China’s industrialization and urbanization. Sci. Total Environ. 2021, 758, 143688. [Google Scholar] [CrossRef] [PubMed]
- Meinshausen, M.; Meinshausen, N.; Hare, W.; Raper, S.C.; Frieler, K.; Knutti, R.; Frame, D.J.; Allen, M.R. Greenhouse-gas emission targets for limiting global warming to 2 degrees C. Nature 2009, 458, 1158–1162. [Google Scholar] [CrossRef] [PubMed]
- Liang, W.; Yang, M. Urbanization, economic growth and environmental pollution: Evidence from China. Sustain. Comput.-Inf. 2019, 21, 1–9. [Google Scholar] [CrossRef]
- Zhao, L.; Lee, X.; Smith, R.B.; Oleson, K. Strong contributions of local background climate to urban heat islands. Nature 2014, 511, 216–219. [Google Scholar] [CrossRef]
- Kampa, M.; Castanas, E. Human health effects of air pollution. Environ. Pollut. 2008, 151, 362–367. [Google Scholar] [CrossRef]
- Tan, J.; Zheng, Y.; Tang, X.; Guo, C.; Li, L.; Song, G.; Zhen, X.; Yuan, D.; Kalkstein, A.J.; Li, F. The urban heat island and its impact on heat waves and human health in Shanghai. Int. J. Biometeorol. 2010, 54, 75–84. [Google Scholar] [CrossRef]
- Dominici, F.; Peng, R.D.; Barr, C.D.; Bell, M.L. Protecting Human Health from Air Pollution. Epidemiology 2010, 21, 187–194. [Google Scholar] [CrossRef] [Green Version]
- Hansen, M.M.; Reo, J.; Kirsten, T. Shinrin-Yoku (Forest Bathing) and Nature Therapy: A State-of-the-Art Review. Int. J. Environ. Res. Public Health 2017, 14, 851. [Google Scholar] [CrossRef] [Green Version]
- Han, J.; Choi, H.; Jeon, Y.; Yoon, C.; Woo, J.; Kim, W. The Effects of Forest Therapy on Coping with Chronic Widespread Pain: Physiological and Psychological Differences between Participants in a Forest Therapy Program and a Control Group. Int. J. Environ. Res. Public Health 2016, 13, 255. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Tsunetsugu, Y.; Takayama, N.; Park, B.; Li, Q.; Song, C.; Komatsu, M.; Ikei, H.; Tyrvainen, L.; Kagawa, T. Influence of Forest Therapy on Cardiovascular Relaxation in Young Adults. Evid.-Based Complement. Altern. 2014, 2014, 834360. [Google Scholar] [CrossRef] [PubMed]
- Jia, B.B.; Yang, Z.X.; Mao, G.X.; Lyu, Y.D.; Wen, X.L.; Xu, W.H.; Lyu, X.L.; Cao, Y.B.; Wang, G.F. Health Effect of Forest Bathing Trip on Elderly Patients with Chronic Obstructive Pulmonary Disease. Biomed. Environ. Sci. 2016, 29, 212–218. [Google Scholar] [PubMed]
- Kim, W.; Lim, S.; Chung, E.; Woo, J. The Effect of Cognitive Behavior Therapy-Based Psychotherapy Applied in a Forest Environment an Physiological Changes and Remission of Major Depressive Disorder. Psychiatry Investig. 2009, 6, 245–254. [Google Scholar] [CrossRef]
- Janeczko, E.; Bielinis, E.; Wojcik, R.; Woznicka, M.; Kedziora, W.; Lukowski, A.; Elsadek, M.; Szyc, K.; Janeczko, K. When Urban Environment Is Restorative: The Effect of Walking in Suburbs and Forests on Psychological and Physiological Relaxation of Young Polish Adults. Forests 2020, 11, 591. [Google Scholar] [CrossRef]
- Song, C.; Ikei, H.; Igarashi, M.; Takagaki, M.; Miyazaki, Y. Physiological and Psychological Effects of a Walk in Urban Parks in Fall. Int. J. Environ. Res. Public Health 2015, 12, 14216–14228. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Wang, R.; Zhang, X.; Liu, J.; Wang, D. Effects of Different Site Conditions on the Concentration of Negative Air Ions in Mountain Forest Based on an Orthogonal Experimental Study. Sustainability 2021, 13, 12012. [Google Scholar] [CrossRef]
- Liu, X.; Yu, X.; Zhang, Z. PM2.5 Concentration Differences between Various Forest Types and Its Correlation with Forest Structure. Atmosphere 2015, 6, 1801–1815. [Google Scholar] [CrossRef] [Green Version]
- Sahar, S.; Zhang, H.; Chi, X.; Felix, M.; Li, H. The influence of spatial configuration of green areas on microclimate and thermal comfort. Urban For. Urban Green. 2018, 34, S945839190. [Google Scholar]
- Salbitano, F. Thermal Comfort and Perceptions of the Ecosystem Services and Disservices of Urban Trees in Florence. Forests 2021, 12, 1387. [Google Scholar]
- Zhu, S.; Hu, F.; He, S.; Qiu, Q.; Su, Y.; He, Q.; Li, J. Comprehensive Evaluation of Healthcare Benefits of Different Forest Types: A Case Study in Shimen National Forest Park, China. Forests 2021, 12, 207. [Google Scholar] [CrossRef]
- Zhang, Z.; Long, T.; He, G.; Wei, M.; Tang, C.; Wang, W.; Wang, G.; She, W.; Zhang, X. Study on Global Burned Forest Areas Based on Landsat Data. Photogramm. Eng. Remote Sens. 2020, 86, 503–508. [Google Scholar] [CrossRef]
- Fan, S.; Zhang, M.; Li, Y.; Li, K.; Dong, L. Impacts of Composition and Canopy Characteristics of Plant Communities on Microclimate and Airborne Particles in Beijing, China. Sustainability 2021, 14, 4791. [Google Scholar] [CrossRef]
- Peters, E.B.; Mcfadden, J.P. Influence of seasonality and vegetation type on suburban microclimates. Urban Ecosyst. 2010, 13, 443–460. [Google Scholar] [CrossRef] [Green Version]
- Yan, X.; Wang, H.; Hou, Z.; Wang, S.; Zhang, D.; Xu, Q.; Tokola, T. Spatial analysis of the ecological effects of negative air ions in urban vegetated areas: A case study in Maiji, China. Urban For. Urban Green. 2015, 13, 94791. [Google Scholar] [CrossRef]
- de Freitas, C.R.; Grigorieva, E.A. A comparison and appraisal of a comprehensive range of human thermal climate indices. Int. J. Biometeorol. 2017, 61, 487–512. [Google Scholar] [CrossRef] [PubMed]
- De Jendritzky, G.; Dear, R.; Havenith, G. UTCI-Why another thermal index? Int. J. Biometeorol. 2012, 56, 421–428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brode, P.; Fiala, D.; Błażejczyk, A.; Ejczyk, K.; Holmér, I.; Jendritzky, G.; Kampmann, B.; Tinz, B.; Havenith, G. Deriving the operational procedure for the Universal Thermal Climate Index (UTCI). Int. J. Biometeorol. 2012, 56, 481–494. [Google Scholar] [CrossRef] [Green Version]
- Potchter, O.; Cohen, P.; Lin, T.P.; Matzarakis, A. Outdoor human thermal perception in various climates: A comprehensive review of approaches, methods and quantification. Sci. Total Environ. 2018, 631, 390–406. [Google Scholar] [CrossRef]
- Xu, M.; Hong, B.; Jiang, R.; An, L.; Zhang, T. Outdoor thermal comfort of shaded spaces in an urban park in the cold region of China. Build. Environ. 2019, 155, 408–420. [Google Scholar] [CrossRef]
- ISO 7726. Thermal Environment-Instruments and Method for Measuring Physical Quantities. International Organization for Standardization, Case postale 56, 1211 Genève, Switzerland, 1 July 1985. 39p. Illus. Available online: http://www.ilo.org/dyn/cisdoc2/cismain.details?p_lang=es&p_doc_id=44729 (accessed on 15 February 2022).
- Chen, Y.; Chang, Z.; Xu, S.; Qi, P.; Tang, X.; Song, Y.; Liu, D. Altitudinal Gradient Characteristics of Spatial and Temporal Variations of Snowpack in the Changbai Mountain and Their Response to Climate Change. Water 2021, 13, 3580. [Google Scholar] [CrossRef]
- Jing, Y.; Fang, M.; Ohsawa Tatuo, K. Vertical vegetation zones along 30° N latitude in humid East Asia. Plant Ecol. 1996, 126, 135–149. [Google Scholar]
- Rajsnerova, P.; Klem, K.; Holub, P.; Novotna, K.; Vecerova, K.; Kozacikova, M.; Rivas-Ubach, A.; Sardans, J.; Marek, M.V.; Penuelas, J. Morphological, biochemical and physiological traits of upper and lower canopy leaves of European beech tend to converge with increasing altitude. Tree Physiol. 2015, 35, 47–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, W.; Zeng, C.; Lam, N.N.; Liu, Z.; Tao, J.; Zhang, X.; Lyu, B.; Li, N.; Li, D.; Chen, Q. Study of the relationship between the spatial structure and thermal comfort of a pure forest with four distinct seasons at the microscale level. Urban For. Urban Green. 2021, 62, 127168. [Google Scholar] [CrossRef]
- Hardin, P.J.; Jensen, R.R. The effect of urban leaf area on summertime urban surface kinetic temperatures: A Terre Haute case study. Urban For. Urban Green. 2007, 6, 63–72. [Google Scholar] [CrossRef]
- Jones, A.M.; Harrison, R.M. The effects of meteorological factors on atmospheric bioaerosol concentrations—A review. Sci. Total Environ. 2004, 326, 151–180. [Google Scholar] [CrossRef] [PubMed]
- Ye, W.; Hao, J.; Fu, L.; Wang, Z.; Tang, U. Vertical and horizontal profiles of airborne particulate matter near major roads in Macao, China. Atmos. Environ. 2002, 36, 4907–4918. [Google Scholar]
- Lee, L.; Jim, C.Y. Urban woodland on intensive green roof improved outdoor thermal comfort in subtropical summer. Int. J. Biometeorol. 2019, 63, 895–909. [Google Scholar] [CrossRef]
- Zheng, B.; Bedra, K.B.; Zheng, J.; Wang, G. Combination of Tree Configuration with Street Configuration for Thermal Comfort Optimization under Extreme Summer Conditions in the Urban Center of Shantou City, China. Sustainability 2018, 10, 4192. [Google Scholar] [CrossRef] [Green Version]
- Rui, L.; Riccardo, B.; Gao, Z.; Ding, W.; Shen, J. The Impact of Green Space Layouts on Microclimate and Air Quality in Residential Districts of Nanjing, China. Forests 2018, 9, 224. [Google Scholar] [CrossRef] [Green Version]
- Ling, X.; Jayaratne, R.; Morawska, L. Air ion concentrations in various urban outdoor environments. Atmos. Environ. 2010, 44, 2186–2193. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Xie, Z.; Chen, B.; Kuang, K.; Xu, D.; Liu, J.; He, Z. Different Time Scale Distribution of Negative Air Ions Concentrations in Mount Wuyi National Park. Int. J. Environ. Res. Public Health 2021, 18, 5037. [Google Scholar] [CrossRef] [PubMed]
- Iwama, H. Negative air ions created by water shearing improve erythrocyte deformability and aerobic metabolism. Indoor Air 2004, 14, 293–297. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Li, S. Changes in negative air ions concentration under different light intensities and development of a model to relate light intensity to directional change. J. Environ. Manag. 2009, 90, 2746–2754. [Google Scholar] [CrossRef] [PubMed]
- Miao, S.; Zhang, X.; Han, Y.; Sun, W.; Liu, C.; Yin, S. Random Forest Algorithm for the Relationship between Negative Air Ions and Environmental Factors in an Urban Park. Atmosphere 2018, 9, 463. [Google Scholar] [CrossRef] [Green Version]
- Kwak, M.J.; Lee, J.; Kim, H.; Park, S.; Lim, Y.; Kim, J.E.; Baek, S.G.; Seo, S.M.; Kim, K.N.; Woo, S.Y. The Removal Efficiencies of Several Temperate Tree Species at Adsorbing Airborne Particulate Matter in Urban Forests and Roadsides. Forests 2019, 10, 960. [Google Scholar] [CrossRef] [Green Version]
- Lu, S.; Yang, X.; Li, S.; Chen, B.; Jiang, Y.; Wang, D.; Xu, L. Effects of plant leaf surface and different pollution levels on PM2.5 adsorption capacity. Urban For. Urban Green. 2018, 34, 64–70. [Google Scholar] [CrossRef]
- Zhang, W.; Wang, B.; Niu, X. Study on the Adsorption Capacities for Airborne Particulates of Landscape Plants in Different Polluted Regions in Beijing (China). Int. J. Environ. Res. Public Health 2015, 12, 9623–9638. [Google Scholar] [CrossRef] [Green Version]
- Srivanit, M.; Hokao, K. Evaluating the cooling effects of greening for improving the outdoor thermal environment at an institutional campus in the summer. Build. Environ. 2013, 66, 158–172. [Google Scholar] [CrossRef]
- Wang, X.; Cheng, H.; Xi, J.; Yang, G.; Zhao, Y. Relationship between Park Composition, Vegetation Characteristics and Cool Island Effect. Sustainability 2018, 10, 587. [Google Scholar] [CrossRef] [Green Version]
- Massetti, L.; Petralli, M.; Napoli, M.; Brandani, G.; Orlandini, S.; Pearlmutter, D. Effects of deciduous shade trees on surface temperature and pedestrian thermal stress during summer and autumn. Int. J. Biometeorol. 2019, 63, 467–479. [Google Scholar] [CrossRef]
- Afshar, N.K.; Karimian, Z.; Doostan, R.; Nokhandan, M.H. Influence of planting designs on winter thermal comfort in an urban park. J. Environ. Eng. Landsc. 2018, 26, 232–240. [Google Scholar] [CrossRef]
- Cao, Z.; Wu, X.; Wang, T.; Zhao, Y.; Zhao, Y.; Wang, D.; Chang, Y.; Wei, Y.; Yan, G.; Fan, Y. Characteristics of airborne particles retained on conifer needles across China in winter and preliminary evaluation of the capacity of trees in haze mitigation. Sci. Total Environ. 2022, 806, 150704. [Google Scholar] [CrossRef] [PubMed]
- Tong, Z.; Whitlow, T.H.; MacRae, P.F.; Landers, A.J.; Harada, Y. Quantifying the effect of vegetation on near-road air quality using brief campaigns. Environ. Pollut. 2015, 201, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.; Guo, J.; Wheeler, S.; Kan, L.; Che, S. Evaluation of impacts of trees on PM2.5 dispersion in urban streets. Atmos. Environ. 2014, 99, 277–287. [Google Scholar] [CrossRef]
- Janhall, S. Review on urban vegetation and particle air pollution–Deposition and dispersion. Atmos. Environ. 2015, 105, 130–137. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Sample Code | Altitude (m) | Plant Community | Average Height (m) | Canopy Density | Average DBH (cm) |
|---|---|---|---|---|---|---|
| A1 | QF856 | 856 | Quercus variabilis forest | 16.81 | 0.68 | 19.82 |
| A2 | QF931 | 931 | Quercus variabilis forest | 16.72 | 0.70 | 20.45 |
| A3 | QF997 | 997 | Quercus variabilis forest | 18.45 | 0.65 | 22.64 |
| A4 | QF1052 | 1052 | Quercus variabilis forest | 15.29 | 0.73 | 21.26 |
| A5 | QF1231 | 1231 | Quercus variabilis forest | 17.93 | 0.62 | 24.47 |
| A6 | QF1364 | 1364 | Quercus variabilis forest | 16.57 | 0.68 | 23.70 |
| A7 | QF1406 | 1406 | Quercus variabilis forest | 16.05 | 0.71 | 21.95 |
| A8 | QF1463 | 1463 | Quercus variabilis forest | 15.70 | 0.69 | 22.51 |
| A9 | QF1508 | 1508 | Quercus variabilis forest | 15.16 | 0.70 | 20.23 |
| No. | Composition Types | Altitude (m) | Plant Community | Average Height (m) | Canopy Density | Average DBH (cm) |
|---|---|---|---|---|---|---|
| B1 | QVF | 956 | Quercus variabilis forest | 16.77 | 0.73 | 18.84 |
| B2 | 937 | Populus davidiana forest | 13.31 | 0.53 | 17.43 | |
| B3 | RPF | 863 | Robinia pseudoacacia forest | 9.28 | 0.64 | 13.57 |
| B4 | PTF | 876 | Pinus tabuliformis forest | 12.83 | 0.58 | 15.28 |
| B5 | POF | 921 | Platycladus orientalis forest | 8.14 | 0.65 | 12.35 |
| B6 | WTF | 834 | Weed-tree forest | 7.62 | 0.67 | 10.26 |
| No. | Altitude (m) | Plant Community | Average Height (m) | Canopy Density | Average DBH (cm) |
|---|---|---|---|---|---|
| C1 | 924 | Quercus variabilis forest | 23.83 | 83.92 | 17.03 |
| C2 | 909 | Quercus variabilis forest | 21.18 | 75.82 | 19.24 |
| C3 | 896 | Quercus variabilis forest | 16.34 | 66.37 | 23.61 |
| C4 | 871 | Quercus variabilis forest | 17.67 | 53.64 | 26.77 |
| C5 | 933 | Quercus variabilis forest | 16.86 | 44.31 | 24.58 |
| C6 | 859 | Quercus variabilis forest | 14.25 | 36.79 | 28.23 |
| Stress Category | Standard UTCI (°C) Range | UTCI (°C) Range (Shaanxi Province, China) [29] |
|---|---|---|
| Extreme heat stress | >46 | >42.1 |
| Very strong heat stress | 38 to 46 | 40.6 to 42.1 |
| Strong heat stress | 32 to 38 | 36.0 to 40.6 |
| Moderate heat stress | 26 to 32 | 29.1 to 36.0 |
| No thermal stress | 9 to 26 | 18.0 to 29.1 |
| Slight cold stress | 9 to 0 | 11.3 to 18.0 |
| Moderate cold stress | 0 to −13 | 7.8 to 11.3 |
| Strong cold stress | −13 to −27 | 4.2 to 7.8 |
| Very strong cold stress | −27 to −40 | 2.0 to 4.2 |
| Extreme cold stress | <−40 | <2.0 |
| No. | CD (%) | CP (%) | LAI | SVF |
|---|---|---|---|---|
| C1 | 83.92 | 42.21 | 2.69 | 0.10 |
| C2 | 75.82 | 53.68 | 2.53 | 0.15 |
| C3 | 66.37 | 56.92 | 2.27 | 0.24 |
| C4 | 53.64 | 58.06 | 1.88 | 0.32 |
| C5 | 44.31 | 63.63 | 1.51 | 0.40 |
| C6 | 36.79 | 71.33 | 1.30 | 0.48 |
| No. | dUTCI (°C) | dNAI (ions/cm3) | dPM2.5 (µg/m3) | dPM10 (µg/m3) |
|---|---|---|---|---|
| C1 | 29.86 | 1571.03 | 19.47 | 41.82 |
| C2 | 30.24 | 1594.17 | 19.11 | 43.44 |
| C3 | 30.82 | 1413.11 | 20.66 | 46.52 |
| C4 | 31.95 | 1488.34 | 21.23 | 52.85 |
| C5 | 32.88 | 1337.26 | 20.46 | 48.11 |
| C6 | 33.03 | 1081.63 | 21.07 | 49.37 |
| No. | CD | CP | LAI | SVF | ||||
|---|---|---|---|---|---|---|---|---|
| cc | Sig. | cc | Sig. | cc | Sig. | cc | Sig. | |
| dUTCI | −0.992 ** | 0.000 | 0.909 * | 0.012 | −0.995 ** | 0.000 | 0.986 ** | 0.000 |
| dNAI | 0.866 * | 0.026 | −0.877 * | 0.022 | 0.877 * | 0.022 | −0.899 * | 0.015 |
| dPM2.5 | −0.792 | 0.060 | 0.692 | 0.127 | −0.763 | 0.078 | 0.794 | 0.060 |
| dPM10 | −0.793 | 0.060 | 0.692 | 0.128 | −0.755 | 0.075 | 0.767 | 0.075 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, R.; Chen, Q.; Wang, D. Effects of Altitude, Plant Communities, and Canopies on the Thermal Comfort, Negative Air Ions, and Airborne Particles of Mountain Forests in Summer. Sustainability 2022, 14, 3882. https://doi.org/10.3390/su14073882
Wang R, Chen Q, Wang D. Effects of Altitude, Plant Communities, and Canopies on the Thermal Comfort, Negative Air Ions, and Airborne Particles of Mountain Forests in Summer. Sustainability. 2022; 14(7):3882. https://doi.org/10.3390/su14073882
Chicago/Turabian StyleWang, Rui, Qi Chen, and Dexiang Wang. 2022. "Effects of Altitude, Plant Communities, and Canopies on the Thermal Comfort, Negative Air Ions, and Airborne Particles of Mountain Forests in Summer" Sustainability 14, no. 7: 3882. https://doi.org/10.3390/su14073882