
Underwater Hyperspectral Imaging (UHI): A Review of Systems and Applications for Proximal Seafloor Ecosystem Studies

 , , and
, , and 
Abstract
:1. Introduction
1.1. Background
1.2. Marine Benthic Imaging—A Tale of Three Bands
2. Methods and Scope of This Review
3. Analysis of Underwater Hyperspectral Imaging (UHI)
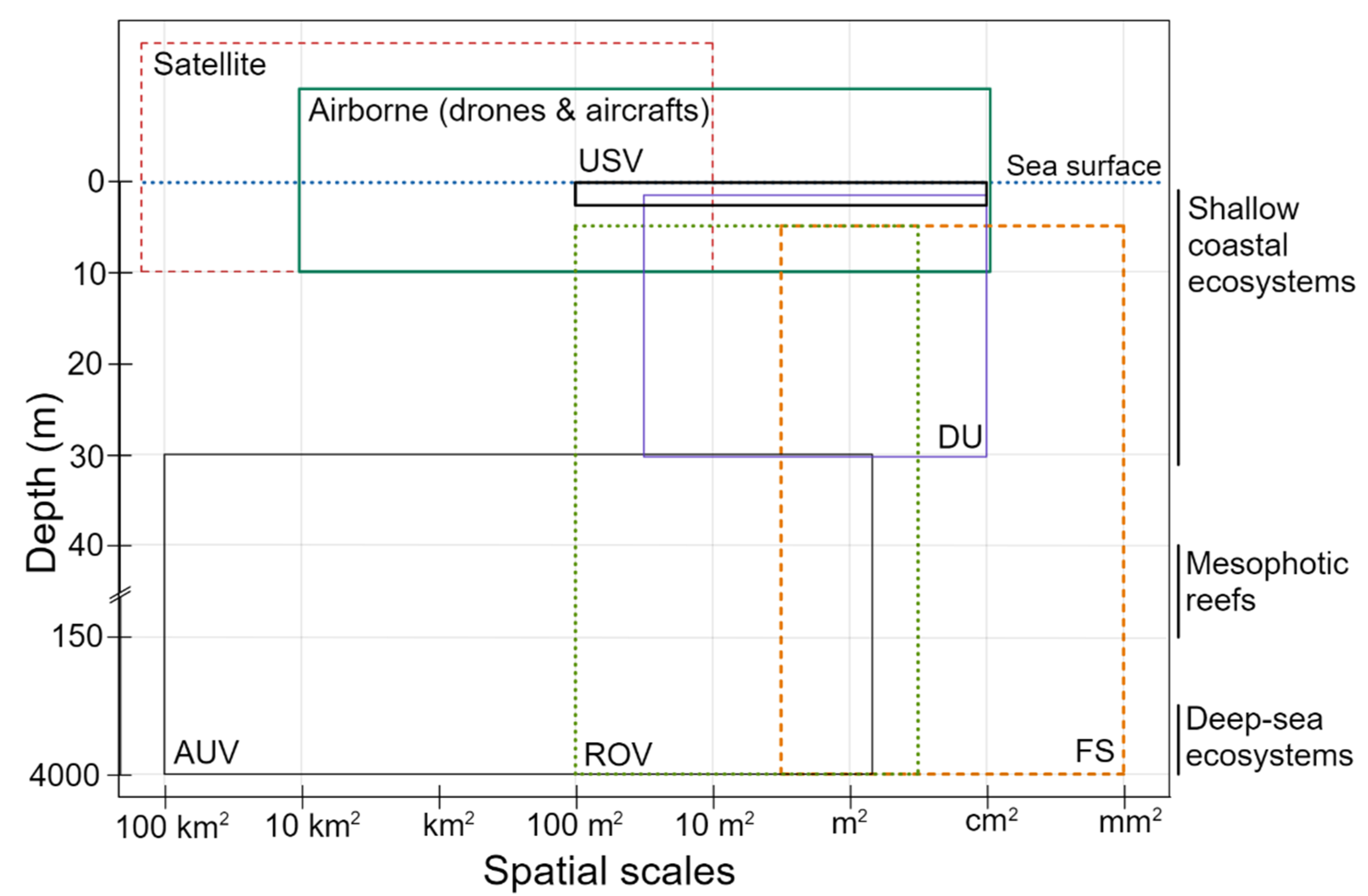
3.1. Applied UHI Systems and Sensor Architectures
3.1.1. Fixed Underwater Motorized Rails
3.1.2. Underwater Unmanned Vehicles (UUVs)
3.1.3. Unmanned Surface Vehicles (USVs)
3.1.4. Under-ice Sliding Platforms
3.1.5. Diver Operated Units (DU)
3.1.6. Fixed Stations & Networks
3.1.7. In Vitro and Ex Situ-Based Systems
4. Breakdown of Applications and the Importance of Pigments for UHI
4.1. Microphytobenthos and Sediment Phytodetritus
4.2. Coral Reefs
4.2.1. Warm-Water Corals
4.2.2. Cold-Water Corals
4.3. Coralline Algae
4.3.1. Non-Geniculate (crustose) Coralline Algae
4.3.2. Geniculate Coralline Algae
4.4. Sponges
4.5. Oyster Reefs
4.6. Sympagic Environments
4.7. Seafloor Areas with Mineral Resources
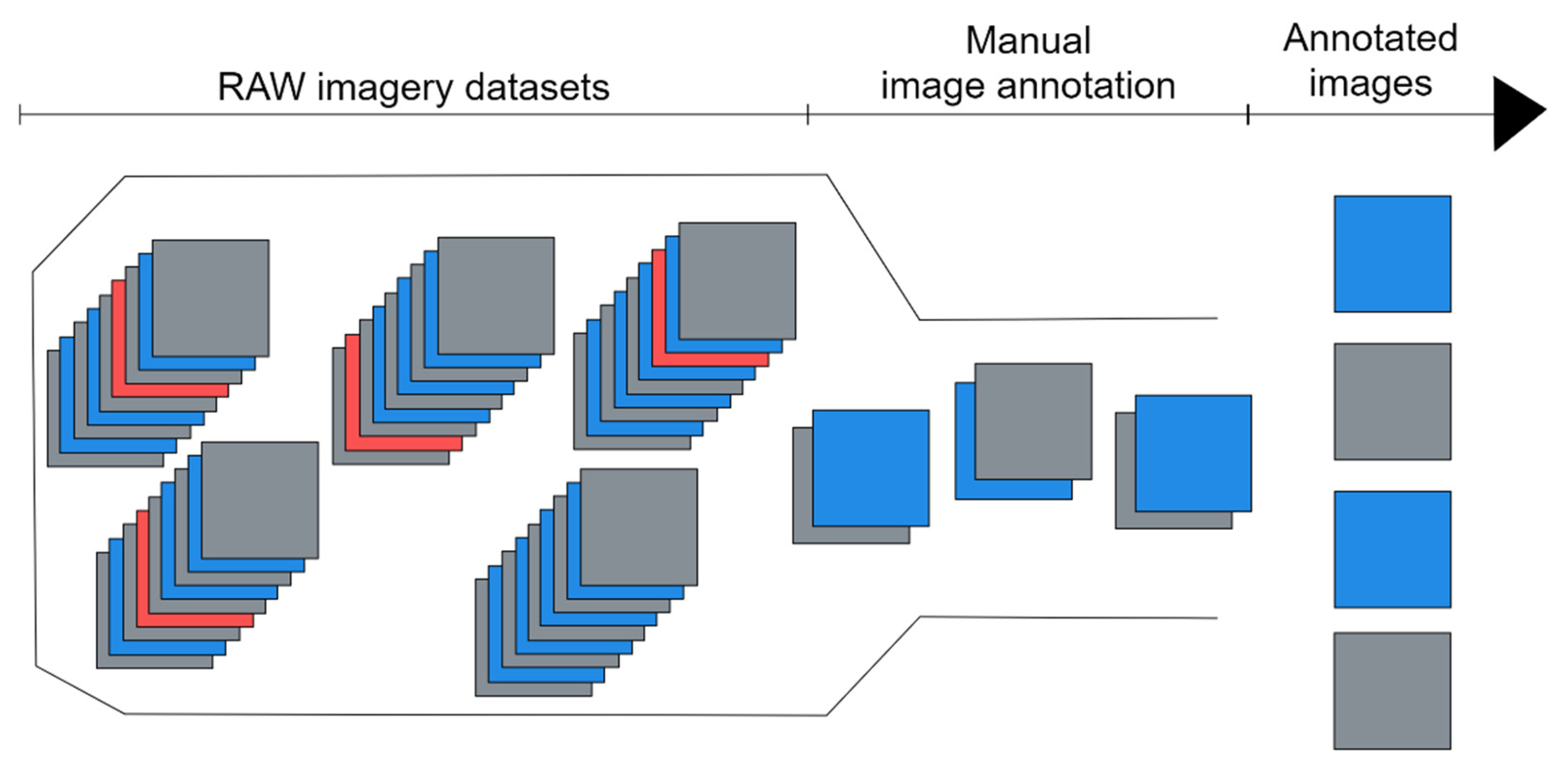
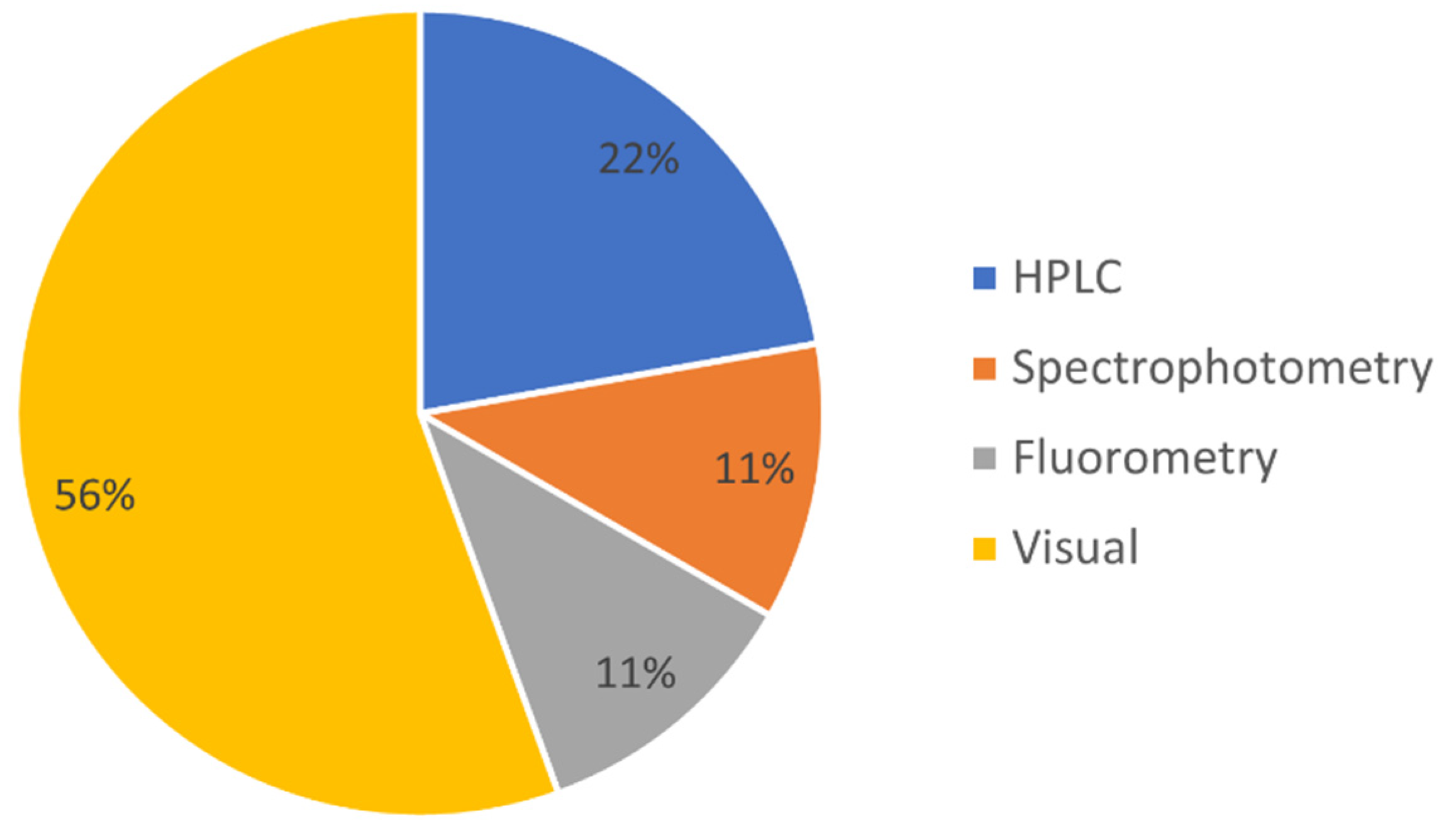
5. UHI Validation and Calibration: Pigment Extraction and Specimen Identification
6. Discussion of Technical Challenges for UHI Systems for Seafloor Observations
6.1. Variable Survey Altitude and Uneven Illumination Effects
6.2. Navigation, Georeferencing, and Survey Procedures
6.3. UHI Data Processing
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Costello, M.J.; Chaudhary, C. Marine Biodiversity, Biogeography, Deep-Sea Gradients, and Conservation. Curr. Biol. 2017, 27, R511–R527. [Google Scholar] [CrossRef]
- Chave, J. The problem of pattern and scale in ecology: What have we learned in 20 years? Ecol. Lett. 2013, 16, 4–16. [Google Scholar] [CrossRef]
- Halpern, B.S.; Frazier, M.; Afflerbach, J.; Lowndes, J.S.; Micheli, F.; O’Hara, C.; Scarborough, C.; Selkoe, K.A. Recent pace of change in human impact on the world’s ocean. Sci. Rep. 2019, 9, 11609. [Google Scholar] [CrossRef] [Green Version]
- McCauley, D.J.; Pinsky, M.L.; Palumbi, S.R.; Estes, J.A.; Joyce, F.H.; Warner, R.R. Marine defaunation: Animal loss in the global ocean. Science 2015, 347, 6219. [Google Scholar] [CrossRef] [Green Version]
- Borja, A.; Andersen, J.H.; Arvanitidis, C.D.; Basset, A.; Buhl-Mortensen, L.; Carvalho, S.; Dafforn, K.A.; Devlin, M.J.; Escobar-Briones, E.G.; Grenz, C.; et al. Past and Future Grand Challenges in Marine Ecosystem Ecology. Front. Mar. Sci. 2020, 7, 362. [Google Scholar] [CrossRef]
- Gamfeldt, L.; Lefcheck, J.S.; Byrnes, J.E.K.; Cardinale, B.J.; Duffy, J.E.; Griffin, J.N. Marine biodiversity and ecosystem functioning: What’s known and what’s next? Oikos 2015, 124, 252–265. [Google Scholar] [CrossRef] [Green Version]
- Kung, A.; Svobodova, K.; Lèbre, E.; Valenta, R.; Kemp, D.; Owen, J.R. Governing deep sea mining in the face of uncertainty. J. Environ. Manag. 2021, 279, 111593. [Google Scholar] [CrossRef]
- Payne, M.R.; Barange, M.; Cheung, W.W.L.; MacKenzie, B.R.; Batchelder, H.P.; Cormon, X.; Eddy, T.D.; Fernandes, J.A.; Hollowed, A.B.; Jones, M.C.; et al. Uncertainties in projecting climate-change impacts in marine ecosystems. ICES J. Mar. Sci. 2016, 73, 1272–1282. [Google Scholar] [CrossRef]
- Danovaro, R.; Aguzzi, J.; Fanelli, E.; Billett, D.; Gjerde, K.; Jamieson, A.; Ramirez-Llodra, E.; Smith, C.R.; Snelgrove, P.V.R.; Thomsen, L.; et al. An ecosystem-based deep-ocean strategy. Science 2017, 355, 452–454. [Google Scholar] [CrossRef]
- Kachelriess, D.; Wegmann, M.; Gollock, M.; Pettorelli, N. The application of remote sensing for marine protected area management. Ecol. Indic. 2014, 36, 169–177. [Google Scholar] [CrossRef]
- Muller-Karger, F.E.; Hestir, E.; Ade, C.; Turpie, K.; Roberts, D.A.; Siegel, D.; Miller, R.J.; Humm, D.; Izenberg, N.; Keller, M.; et al. Satellite sensor requirements for monitoring essential biodiversity variables of coastal ecosystems. Ecol. Appl. 2018, 28, 749–760. [Google Scholar] [CrossRef]
- Kuchler, D.A.; Jupp, D.L.B.; Daniel, D.B.; Bour, W. Coral Reef Remote Sensing; Goodman, J.A., Purkis, S.J., Phinn, S.R., Eds.; Springer: Dordrecht, The Netherlands, 2013; Volume 1, ISBN 978-90-481-9291-5. [Google Scholar]
- Lesser, M.P.; Mobley, C.D. Bathymetry, water optical properties, and benthic classification of coral reefs using hyperspectral remote sensing imagery. Coral Reefs 2007, 26, 819–829. [Google Scholar] [CrossRef]
- Bioucas-Dias, J.M.; Plaza, A.; Camps-Valls, G.; Scheunders, P.; Nasrabadi, N.M.; Chanussot, J. Hyperspectral remote sensing data analysis and future challenges. IEEE Geosci. Remote Sens. Mag. 2013, 1, 6–36. [Google Scholar] [CrossRef] [Green Version]
- Lodhi, V.; Chakravarty, D.; Mitra, P. Hyperspectral Imaging for Earth Observation: Platforms and Instruments. J. Indian Inst. Sci. 2018, 98, 429–443. [Google Scholar] [CrossRef]
- Kobryn, H.T.; Wouters, K.; Beckley, L.E.; Heege, T. Ningaloo Reef: Shallow Marine Habitats Mapped Using a Hyperspectral Sensor. PLoS ONE 2013, 8, e70105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tait, L.; Bind, J.; Charan-Dixon, H.; Hawes, I.; Pirker, J.; Schiel, D. Unmanned aerial vehicles (UAVs) for monitoring macroalgal biodiversity: Comparison of RGB and multispectral imaging sensors for biodiversity assessments. Remote Sens. 2019, 11, 2332. [Google Scholar] [CrossRef] [Green Version]
- Vahtmäe, E.; Paavel, B.; Kutser, T. How much benthic information can be retrieved with hyperspectral sensor from the optically complex coastal waters? J. Appl. Remote Sens. 2020, 14, 1. [Google Scholar] [CrossRef]
- Tait, L.W.; Orchard, S.; Schiel, D.R. Missing the Forest and the Trees: Utility, Limits and Caveats for Drone Imaging of Coastal Marine Ecosystems. Remote Sens. 2021, 13, 3136. [Google Scholar] [CrossRef]
- Garcia, R.A.; Lee, Z.; Hochberg, E.J. Hyperspectral shallow-water remote sensing with an enhanced benthic classifier. Remote Sens. 2018, 10, 147. [Google Scholar] [CrossRef] [Green Version]
- Hoegh-Guldberg, O.; Poloczanska, E.S.; Skirving, W.; Dove, S. Coral reef ecosystems under climate change and ocean acidification. Front. Mar. Sci. 2017, 4, 158. [Google Scholar] [CrossRef] [Green Version]
- Ramirez-Llodra, E.; Brandt, A.; Danovaro, R.; De Mol, B.; Escobar, E.; German, C.R.; Levin, L.A.; Martinez Arbizu, P.; Menot, L.; Buhl-Mortensen, P.; et al. Deep, diverse and definitely different: Unique attributes of the world’s largest ecosystem. Biogeosciences 2010, 7, 2851–2899. [Google Scholar] [CrossRef] [Green Version]
- Kaiser, S.; Brandão, S.N.; Brix, S.; Barnes, D.K.A.; Bowden, D.A.; Ingels, J.; Leese, F.; Schiaparelli, S.; Arango, C.P.; Badhe, R.; et al. Patterns, processes and vulnerability of Southern Ocean benthos: A decadal leap in knowledge and understanding. Mar. Biol. 2013, 160, 2295–2317. [Google Scholar] [CrossRef]
- Brasier, M.J.; Barnes, D.; Bax, N.; Brandt, A.; Christianson, A.B.; Constable, A.J.; Downey, R.; Figuerola, B.; Griffiths, H.; Gutt, J.; et al. Responses of Southern Ocean Seafloor Habitats and Communities to Global and Local Drivers of Change. Front. Mar. Sci. 2021, 8, 109. [Google Scholar] [CrossRef]
- Aasen, H.; Honkavaara, E.; Lucieer, A.; Zarco-Tejada, P.J. Quantitative remote sensing at ultra-high resolution with UAV spectroscopy: A review of sensor technology, measurement procedures, and data correctionworkflows. Remote Sens. 2018, 10, 1091. [Google Scholar] [CrossRef] [Green Version]
- Johnsen, G.; Volent, Z.; Dierssen, H.; Pettersen, R.; Van Ardelan, M.; Søreide, F.; Fearns, P.; Ludvigsen, M.; Moline, M. Underwater Hyperspectral Imagery to Create Biogeochemical maps of Seafloor Properties. In Woodhead Publishing Series in Electronic and Optical Materials, Subsea Optics and Imaging; Woodhead Publishing: Cambridge, UK, 2013; ISBN 9780857093417. [Google Scholar]
- Chennu, A.; Färber, P.; Volkenborn, N.; Al-Najjar, M.A.A.; Janssen, F.; de Beer, D.; Polerecky, L. Hyperspectral imaging of the microscale distribution and dynamics of microphytobenthos in intertidal sediments. Limnol. Oceanogr. Methods 2013, 11, 511–528. [Google Scholar] [CrossRef] [Green Version]
- Aguzzi, J.; Chatzievangelou, D.; Marini, S.; Fanelli, E.; Danovaro, R.; Flögel, S.; Lebris, N.; Juanes, F.; De Leo, F.C.; Del Rio, J.; et al. New High-Tech Flexible Networks for the Monitoring of Deep-Sea Ecosystems. Environ. Sci. Technol. 2019, 53, 6616–6631. [Google Scholar] [CrossRef] [Green Version]
- Aguzzi, J.; Chatzievangelou, D.; Francescangeli, M.; Marini, S.; Bonofiglio, F.; Del Rio, J.; Danovaro, R. The hierarchic treatment of marine ecological information from spatial networks of benthic platforms. Sensors 2020, 20, 1751. [Google Scholar] [CrossRef] [Green Version]
- Eisenhauer, N.; Schielzeth, H.; Barnes, A.D.; Barry, K.E.; Bonn, A.; Brose, U.; Bruelheide, H.; Buchmann, N.; Buscot, F.; Ebeling, A.; et al. A multitrophic perspective on biodiversity–ecosystem functioning research. Adv. Ecol. Res. 2019, 61, 1–54. [Google Scholar]
- Bicknell, A.W.J.; Godley, B.J.; Sheehan, E.V.; Votier, S.C.; Witt, M.J. Camera technology for monitoring marine biodiversity and human impact. Front. Ecol. Environ. 2016, 14, 424–432. [Google Scholar] [CrossRef]
- Morris, K.J.; Bett, B.J.; Durden, J.M.; Huvenne, V.A.I.; Milligan, R.; Jones, D.O.B.; McPhail, S.; Robert, K.; Bailey, D.M.; Ruhl, H.A. A new method for ecological surveying of the abyss using autonomous underwater vehicle photography. Limnol. Oceanogr. Methods 2014, 12, 795–809. [Google Scholar] [CrossRef] [Green Version]
- Piazza, P.; Cummings, V.; Guzzi, A.; Hawes, I.; Lohrer, A.; Marini, S.; Marriott, P.; Menna, F.; Nocerino, E.; Peirano, A.; et al. Underwater photogrammetry in Antarctica: Long-term observations in benthic ecosystems and legacy data rescue. Polar Biol. 2019, 42, 1061–1079. [Google Scholar] [CrossRef] [Green Version]
- Ludvigsen, M.; Sørensen, A.J. Towards integrated autonomous underwater operations for ocean mapping and monitoring. Annu. Rev. Control 2016, 42, 145–157. [Google Scholar] [CrossRef]
- Monk, J.; Barrett, N.S.; Peel, D.; Lawrence, E.; Hill, N.A.; Lucieer, V.; Hayes, K.R. An evaluation of the error and uncertainty in epibenthos cover estimates from AUV images collected with an efficient, spatially-balanced design. PLoS ONE 2018, 13, e0203827. [Google Scholar] [CrossRef] [Green Version]
- Schoening, T.; Köser, K.; Greinert, J. An acquisition, curation and management workflow for sustainable, terabyte-scale marine image analysis. Sci. Data 2018, 5, 180181. [Google Scholar] [CrossRef]
- Balazy, P.; Kuklinski, P.; Berge, J. Diver deployed autonomous time-lapse camera systems for ecological studies. J. Mar. Eng. Technol. 2018, 17, 137–142. [Google Scholar] [CrossRef]
- Schoening, T.; Osterloff, J.; Nattkemper, T.W. RecoMIA-recommendations for marine image annotation: Lessons learned and future directions. Front. Mar. Sci. 2016, 3, 59. [Google Scholar] [CrossRef]
- Beijbom, O.; Edmunds, P.J.; Kline, D.I.; Mitchell, B.G.; Kriegman, D. Automated annotation of coral reef survey images. In Proceedings of the 2012 IEEE Conference on Computer Vision and Pattern Recognition, Providence, RI, USA, 16–21 June 2012; pp. 1170–1177. [Google Scholar] [CrossRef] [Green Version]
- Beijbom, O.; Edmunds, P.J.; Roelfsema, C.; Smith, J.; Kline, D.I.; Neal, B.P.; Dunlap, M.J.; Moriarty, V.; Fan, T.Y.; Tan, C.J.; et al. Towards automated annotation of benthic survey images: Variability of human experts and operational modes of automation. PLoS ONE 2015, 10, e0130312. [Google Scholar] [CrossRef]
- Durden, J.M.; Bett, B.J.; Schoening, T.; Morris, K.J.; Nattkemper, T.W.; Ruhl, H.A. Comparison of image annotation data generated by multiple investigators for benthic ecology. Mar. Ecol. Prog. Ser. 2016, 552, 61–70. [Google Scholar] [CrossRef] [Green Version]
- Althaus, F.; Hill, N.; Ferrari, R.; Edwards, L.; Przeslawski, R.; Schönberg, C.H.L.; Stuart-Smith, R.; Barrett, N.; Edgar, G.; Colquhoun, J.; et al. A standardised vocabulary for identifying benthic biota and substrata from underwater imagery: The CATAMI classification scheme. PLoS ONE 2015, 10, e0141039. [Google Scholar] [CrossRef] [PubMed]
- Langenkämper, D.; Zurowietz, M.; Schoening, T.; Nattkemper, T.W. BIIGLE 2.0—Browsing and annotating large marine image collections. Front. Mar. Sci. 2017, 4, 83. [Google Scholar] [CrossRef] [Green Version]
- Williams, I.D.; Couch, C.; Beijbom, O.; Oliver, T.; Vargas-Angel, B.; Schumacher, B.; Brainard, R. Leveraging automated image analysis tools to transform our capacity to assess status and trends on coral reefs. Front. Mar. Sci. 2019, 6, 222. [Google Scholar] [CrossRef] [Green Version]
- Piechaud, N.; Hunt, C.; Culverhouse, P.F.; Foster, N.L.; Howell, K.L. Automated identification of benthic epifauna with computer vision. Mar. Ecol. Prog. Ser. 2019, 615, 15–30. [Google Scholar] [CrossRef]
- Conti, L.A.; Lim, A.; Wheeler, A.J. High resolution mapping of a cold water coral mound. Sci. Rep. 2019, 9, 1016. [Google Scholar] [CrossRef]
- Ventura, D.; Bonifazi, A.; Gravina, M.F.; Belluscio, A.; Ardizzone, G. Mapping and classification of ecologically sensitive marine habitats using unmanned aerial vehicle (UAV) imagery and Object-Based Image Analysis (OBIA). Remote Sens. 2018, 10, 1331. [Google Scholar] [CrossRef] [Green Version]
- Pavoni, G.; Corsini, M.; Callieri, M.; Palma, M.; Scopigno, R. Semantic segmentation of benthic communities from ortho-mosaic maps. ISPRS Ann. Photogramm. Remote Sens. Spat. Inf. Sci. 2019, 42, 151–158. [Google Scholar] [CrossRef] [Green Version]
- Brandt, A.; Gutt, J.; Hildebrandt, M.; Pawlowski, J.; Schwendner, J.; Soltwedel, T.; Thomsen, L. Cutting the umbilical: New technological perspectives in benthic deep-sea research. J. Mar. Sci. Eng. 2016, 4, 36. [Google Scholar] [CrossRef] [Green Version]
- Witte, U.; Wenzhöfer, F.; Sommer, S.; Boetius, A.; Heinz, P.; Aberle, N.; Sand, M.; Cremer, A.; Abraham, W.R.; Jørgensen, B.B.; et al. In situ experimental evidence of the fate of a phytodetritus pulse at the abyssal sea floor. Nature 2003, 424, 763–766. [Google Scholar] [CrossRef] [Green Version]
- Danovaro, R.; Snelgrove, P.V.R.; Tyler, P. Challenging the paradigms of deep-sea ecology. Trends Ecol. Evol. 2014, 29, 465–475. [Google Scholar] [CrossRef]
- Ziegler, A.F.; Cape, M.; Lundesgaard, Ø.; Smith, C.R. Intense deposition and rapid processing of seafloor phytodetritus in a glaciomarine fjord, Andvord Bay (Antarctica). Prog. Oceanogr. 2020, 187, 102413. [Google Scholar] [CrossRef]
- Chapman, M.G.; Tolhurst, T.J.; Murphy, R.J.; Underwood, A.J. Complex and inconsistent patterns of variation in benthos, micro-algae and sediment over multiple spatial scales. Mar. Ecol. Prog. Ser. 2009, 398, 33–47. [Google Scholar] [CrossRef] [Green Version]
- Quijón, P.A.; Kelly, M.C.; Snelgrove, P.V.R. The role of sinking phytodetritus in structuring shallow-water benthic communities. J. Exp. Mar. Bio. Ecol. 2008, 366, 134–145. [Google Scholar] [CrossRef]
- Grémare, A.; Amouroux, J.M.; Charles, F.; Dinet, A.; Riaux-Gobin, C.; Baudart, J.; Medernach, L.; Bodiou, J.Y.; Vétion, G.; Colomines, J.C.; et al. Temporal changes in the biochemical composition and nutritional value of the particulate organic matter available to surface deposit-feeders: A two year study. Mar. Ecol. Prog. Ser. 1997, 150, 195–206. [Google Scholar] [CrossRef] [Green Version]
- Schneider, F.D.; Morsdorf, F.; Schmid, B.; Petchey, O.L.; Hueni, A.; Schimel, D.S.; Schaepman, M.E. Mapping functional diversity from remotely sensed morphological and physiological forest traits. Nat. Commun. 2017, 8, 1441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hennessy, A.; Clarke, K.; Lewis, M. Hyperspectral Classification of Plants: A Review of Waveband Selection Generalisability. Remote Sens. 2020, 12, 113. [Google Scholar] [CrossRef] [Green Version]
- Dumke, I.; Nornes, S.M.; Purser, A.; Marcon, Y.; Ludvigsen, M.; Ellefmo, S.L.; Johnsen, G.; Søreide, F. First hyperspectral imaging survey of the deep seafloor: High-resolution mapping of manganese nodules. Remote Sens. Environ. 2018, 209, 19–30. [Google Scholar] [CrossRef]
- Sture, Ø.; Snook, B.; Ludvigsen, M. Obtaining hyperspectral signatures for seafloor massive sulphide exploration. Minerals 2019, 9, 694. [Google Scholar] [CrossRef] [Green Version]
- Mogstad, A.A.; Ødegård, Ø.; Nornes, S.M.; Ludvigsen, M.; Johnsen, G.; Sørensen, A.J.; Berge, J. Mapping the historical shipwreck Figaro in the high arctic using underwater sensor-carrying robots. Remote Sens. 2020, 12, 997. [Google Scholar] [CrossRef] [Green Version]
- Pettersen, R.; Lein Braa, H.; Gawel, B.A.; Letnes, P.A.; Sæther, K.; Aas, L.M.S. Detection and classification of Lepeophterius salmonis (Krøyer, 1837) using underwater hyperspectral imaging. Aquac. Eng. 2019, 87, 102025. [Google Scholar] [CrossRef]
- Kirk, J.T.O. Light and Photosynthesis in Aquatic Ecosystems, 3rd ed.; Cambridge University Press: Cambridge, UK, 2010; ISBN 9781139168212. [Google Scholar]
- Johnsen, G.; Ludvigsen, M.; Sørensen, A.; Sandvik Aas, L.M. The use of underwater hyperspectral imaging deployed on remotely operated vehicles—Methods and applications. IFAC-PapersOnLine 2016, 49, 476–481. [Google Scholar] [CrossRef]
- Pettersen, R.; Johnsen, G.; Bruheim, P.; Andreassen, T. Development of hyperspectral imaging as a bio-optical taxonomic tool for pigmented marine organisms. Org. Divers. Evol. 2014, 14, 237–246. [Google Scholar] [CrossRef]
- Volent, Z.; Johnsen, G.; Sigernes, F. Microscopic hyperspectral imaging used as a bio-optical taxonomic tool for micro- and macroalgae. Appl. Opt. 2009, 48, 4170–4176. [Google Scholar] [CrossRef]
- Cimoli, E.; Lucieer, A.; Meiners, K.M.; Lund-Hansen, L.C.; Kennedy, F.; Martin, A.; McMinn, A.; Lucieer, V. Towards improved estimates of sea-ice algal biomass: Experimental assessment of hyperspectral imaging cameras for under-ice studies. Ann. Glaciol. 2017, 58, 68–77. [Google Scholar] [CrossRef] [Green Version]
- Mogstad, A.A.; Johnsen, G.; Ludvigsen, M. Shallow-Water Habitat Mapping using Underwater Hyperspectral Imaging from an Unmanned Surface Vehicle: A Pilot Study. Remote Sens. 2019, 11, 685. [Google Scholar] [CrossRef]
- Cimoli, E.; Meiners, K.M.; Lucieer, A.; Lucieer, V. An Under-Ice Hyperspectral and RGB Imaging System to Capture Fine-Scale Biophysical Properties of Sea Ice. Remote Sens. 2019, 11, 2860. [Google Scholar] [CrossRef] [Green Version]
- Cimoli, E.; Meiners, K.; Lund-hansen, L.; Lucieer, V. Spatial variability in sea-ice algal biomass: An under-ice remote sensing perspective. Adv. Polar Sci. 2017, 28, 268–296. [Google Scholar] [CrossRef]
- Sture, O.; Ludvigsen, M.; Soreide, F.; Aas, L.M.S. Autonomous underwater vehicles as a platform for underwater hyperspectral imaging. In Proceedings of the Ocean, Aberdeen, UK, 19–22 June 2017; pp. 1–8. [Google Scholar] [CrossRef]
- Dumke, I.; Purser, A.; Marcon, Y.; Nornes, S.M.; Johnsen, G.; Ludvigsen, M.; Søreide, F. Underwater hyperspectral imaging as an in situ taxonomic tool for deep-sea megafauna. Sci. Rep. 2018, 8, 12860. [Google Scholar] [CrossRef] [Green Version]
- Mogstad, A.A.; Johnsen, G. Spectral characteristics of coralline algae: A multi-instrumental approach, with emphasis on underwater hyperspectral imaging. Appl. Opt. 2017, 56, 9957. [Google Scholar] [CrossRef]
- Foglini, F.; Grande, V.; Marchese, F.; Bracchi, V.A.; Prampolini, M.; Angeletti, L.; Castellan, G.; Chimienti, G.; Hansen, I.M.; Gudmundsen, M.; et al. Application of Hyperspectral Imaging to Underwater Habitat Mapping, Southern Adriatic Sea. Sensors 2019, 19, 2261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chennu, A.; Färber, P.; De’ath, G.; De Beer, D.; Fabricius, K.E. A diver-operated hyperspectral imaging and topographic surveying system for automated mapping of benthic habitats. Sci. Rep. 2017, 7, 7122. [Google Scholar] [CrossRef]
- Dumke, I.; Ludvigsen, M.; Ellefmo, S.L.; Søreide, F.; Johnsen, G.; Murton, B.J. Underwater Hyperspectral Imaging Using a Stationary Platform in the Trans-Atlantic Geotraverse Hydrothermal Field. IEEE Trans. Geosci. Remote Sens. 2019, 57, 2947–2962. [Google Scholar] [CrossRef] [Green Version]
- Cimoli, E.; Lucieer, V.; Meiners, K.M.; Chennu, A.; Castrisios, K.; Ryan, K.G.; Lund-Hansen, L.C.; Martin, A.; Kennedy, F.; Lucieer, A. Mapping the in situ microspatial distribution of ice algal biomass through hyperspectral imaging of sea-ice cores. Sci. Rep. 2020, 10, 21848. [Google Scholar] [CrossRef] [PubMed]
- Letnes, P.A.; Hansen, I.M.; Aas, L.M.S.; Eide, I.; Pettersen, R.; Tassara, L.; Receveur, J.; le Floch, S.; Guyomarch, J.; Camus, L.; et al. Underwater hyperspectral classification of deep sea corals exposed to 2-methylnaphthalene. PLoS ONE 2019, 14, e0209960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Angel, Y.; Turner, D.; Parkes, S.; Malbeteau, Y.; Lucieer, A.; McCabe, M.F. Automated georectification and mosaicking of UAV-based hyperspectral imagery from push-Broom sensors. Remote Sens. 2020, 12, 34. [Google Scholar] [CrossRef] [Green Version]
- Bongiorno, D.L.; Bryson, M.; Bridge, T.C.L.; Dansereau, D.G.; Williams, S.B. Coregistered Hyperspectral and Stereo Image Seafloor Mapping from an Autonomous Underwater Vehicle. J. Field Robot. 2018, 35, 312–329. [Google Scholar] [CrossRef]
- Androuin, T.; Polerecky, L.; Decottignies, P.; Dubois, S.F.; Dupuy, C.; Hubas, C.; Jesus, B.; Le Gall, E.; Marzloff, M.P.; Carlier, A. Subtidal microphytobenthos: A secret garden stimulated by the engineer species Crepidula fornicata. Front. Mar. Sci. 2018, 5, 475. [Google Scholar] [CrossRef]
- Heard, S.B.; Hauser, D.L. Key evolutionary innovations and their ecological mechanisms. Hist. Biol. 1995, 10, 151–173. [Google Scholar] [CrossRef]
- Pearcy, R.W.; Krall, J.P.; Sassenrath-Cole, G.F. Photosynthesis in Fluctuating Light Environments. In Photosynthesis and the Environment; Baker, N.R., Ed.; Springer: Dordrecht, The Netherlands, 1996; pp. 321–346. ISBN 978-0-306-48135-2. [Google Scholar]
- Nishio, J.N. Why are higher plants green? Evolution of the higher plant photosynthetic pigment complement. Plant Cell Environ. 2000, 23, 539–548. [Google Scholar] [CrossRef]
- Hull, P.M. Emergence of modern marine ecosystems. Curr. Biol. 2017, 27, R466–R469. [Google Scholar] [CrossRef] [Green Version]
- Greenwold, M.J.; Cunningham, B.R.; Lachenmyer, E.M.; Pullman, J.M.; Richardson, T.L.; Dudycha, J.L. Diversification of light capture ability was accompanied by the evolution of phycobiliproteins in cryptophyte algae. Proc. R. Soc. B Biol. Sci. 2019, 286, 20190655. [Google Scholar] [CrossRef] [Green Version]
- Horning, N.; Robinson, J.a; Sterling, E.J.; Turner, W.; Spector, S. Remote Sensing for Ecology and Conservation: A Handbook of Techniques; Oxford University Press: Oxford, UK, 2010; Volume 467. [Google Scholar]
- Green, E.; Mumby, P.; Edwards, A.; Clark, C. Remote Sensing Handbook for Tropical Coastal Management; UNESCO Publishing: Paris, France, 2000; ISBN 92-3-103736-6. [Google Scholar]
- Bandaranayake, W.M. The nature and role of pigments of marine invertebrates. Nat. Prod. Rep. 2006, 23, 223–255. [Google Scholar] [CrossRef]
- Marshall, J.; Oberwinkler, J. The colourful world of the mantis shrimp. Nature 1999, 401, 873–874. [Google Scholar] [CrossRef]
- MacColl, R.; Eisele, L.E.; Williams, E.C.; Bowser, S.S. The discovery of a novel R-phycoerythrin from an antarctic red alga. J. Biol. Chem. 1996, 271, 17157–17160. [Google Scholar] [CrossRef] [Green Version]
- Gaitan-Espitia, J.D.; Schmid, M. Antarctic Seaweeds; Gómez, I., Huovinen, P., Eds.; Springer International Publishing: Cham, Switzerland, 2020; ISBN 978-3-030-39447-9. [Google Scholar]
- Michel, C.; Legendre, L.; Demers, S.; Therriault, J.-C. Photoadaptation of sea-ice microalgae in springtime: Photosynthesis and carboxylating enzymes. Mar. Ecol. Prog. Ser. 1988, 50, 177–185. [Google Scholar] [CrossRef]
- Tegdan, J.; Ekehaug, S.; Hansen, I.M.; Aas, L.M.S.; Steen, K.J.; Pettersen, R.; Beuchel, F.; Camus, L. Underwater hyperspectral imaging for environmental mapping and monitoring of seabed habitats. In Proceedings of the OCEANS 2015—Genova, Genova, Italy, 18–21 May 2015; pp. 1–6. [Google Scholar] [CrossRef]
- Verrelst, J.; Malenovský, Z.; Van der Tol, C.; Camps-Valls, G.; Gastellu-Etchegorry, J.P.; Lewis, P.; North, P.; Moreno, J. Quantifying Vegetation Biophysical Variables from Imaging Spectroscopy Data: A Review on Retrieval Methods. Surv. Geophys. 2019, 40, 589–629. [Google Scholar] [CrossRef] [Green Version]
- MacIntyre, H.L.; Geider, R.J.; Miller, D.C. Microphytobenthos: The Ecological Role of the “Secret Garden” of Unvegetated, Shallow-Water Marine Habitats. I. Distribution, Abundance and Primary Production. Estuaries 1996, 19, 186. [Google Scholar] [CrossRef]
- Muscatine, L.; Porter, J.W. Reef Corals: Mutualistic Symbioses Adapted to Nutrient-Poor Environments. Bioscience 1977, 27, 454–460. [Google Scholar] [CrossRef]
- Hochberg, E.J.; Atkinson, M.J.; Andréfouët, S. Spectral reflectance of coral reef bottom-types worldwide and implications for coral reef remote sensing. Remote Sens. Environ. 2003, 85, 159–173. [Google Scholar] [CrossRef]
- Holden, H.; Ledrew, E. Hyperspectral identification of coral reef features. Int. J. Remote Sens. 1999, 20, 2545–2563. [Google Scholar] [CrossRef]
- Mehrubeoglu, M.; Smith, D.K.; Smith, S.W.; Strychar, K.B.; McLauchlan, L. Investigating coral hyperspectral properties across coral species and coral state using hyperspectral imaging. Imaging Spectrom. XVIII 2013, 8870, 88700M. [Google Scholar] [CrossRef]
- Barott, K.; Smith, J.; Dinsdale, E.; Hatay, M.; Sandin, S.; Rohwer, F. Hyperspectral and Physiological Analyses of Coral-Algal Interactions. PLoS ONE 2009, 4, e8043. [Google Scholar] [CrossRef]
- Teague, J.; Willans, J.; Allen, M.; Scott, T.; Day, J. Hyperspectral imaging as a tool for assessing coral health utilising natural fluorescence. J. Spectr. Imaging 2019, 8, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Turley, C.M.; Roberts, J.M.; Guinotte, J.M. Corals in deep-water: Will the unseen hand of ocean acidification destroy cold-water ecosystems? Coral Reefs 2007, 26, 445–448. [Google Scholar] [CrossRef]
- Steneck, R.S. The ecology of coralline algal crusts: Convergent patterns and adaptative strategies. Annaul Rev. Ecol. Syst. 1986, 17, 273–303. [Google Scholar] [CrossRef]
- Hind, K.R.; Starko, S.; Burt, J.M.; Lemay, M.A.; Salomon, A.K.; Martone, P.T. Trophic control of cryptic coralline algal diversity. Proc. Natl. Acad. Sci. USA 2019, 116, 15080–15085. [Google Scholar] [CrossRef] [Green Version]
- Harvey, A.S.; Harvey, R.M.; Merton, E. The distribution, significance and vulnerability of Australian rhodolith beds: A review. Mar. Freshw. Res. 2017, 68, 411–428. [Google Scholar] [CrossRef]
- Jørgensbye, H.I.Ø.; Halfar, J. Overview of coralline red algal crusts and rhodolith beds (Corallinales, Rhodophyta) and their possible ecological importance in Greenland. Polar Biol. 2017, 40, 517–531. [Google Scholar] [CrossRef]
- Vásquez-Elizondo, R.M.; Enríquez, S. Light absorption in coralline algae (Rhodophyta): A morphological and functional approach to understanding species distribution in a coral reef lagoon. Front. Mar. Sci. 2017, 4, 297. [Google Scholar] [CrossRef] [Green Version]
- Perkins, R.G.; Williamson, C.J.; Brodie, J.; Barillé, L.; Launeau, P.; Lavaud, J.; Yallop, M.L.; Jesus, B. Microspatial variability in community structure and photophysiology of calcified macroalgal microbiomes revealed by coupling of hyperspectral and high-resolution fluorescence imaging. Sci. Rep. 2016, 6, 22343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maldonado, M.; Aguilar, R.; Bannister, R.J.; Bell, J.J.; Conway, K.W.; Dayton, P.K.; Díaz, C.; Gutt, J.; Kelly, M.; Kenchington, E.L.R.; et al. Sponge Grounds as Key Marine Habitats: A Synthetic Review of Types, Structure, Functional Roles, and Conservation Concerns. In Marine Animal Forests; Springer International Publishing: Cham, Switzerland, 2015; pp. 1–39. ISBN 9783319170015. [Google Scholar]
- Gutt, J.; Cummings, V.; Dayton, P.; Isla, E.; Jentsch, A.; Schiaparelli, S. Antarctic Marine Animal Forests: Three-Dimensional Communities in Southern Ocean Ecosystems. In Marine Animal Forests: The Ecology of Benthic Biodiversity Hotspots; Rossi, S., Bramanti, L., Gori, A., del Valle, C., Eds.; Springer International Publishing: Cham, Switzerland, 2015; pp. 1–30. ISBN 978-3-319-17001-5. [Google Scholar]
- Du, H.; Jiao, N.; Hu, Y.; Zeng, Y. Diversity and distribution of pigmented heterotrophic bacteria in marine environments. FEMS Microbiol. Ecol. 2006, 57, 92–105. [Google Scholar] [CrossRef] [Green Version]
- Vaughn, C.C.; Hoellein, T.J. Bivalve impacts in freshwater and marine ecosystems. Annu. Rev. Ecol. Evol. Syst. 2018, 49, 183–208. [Google Scholar] [CrossRef] [Green Version]
- Barillé, L.; Le Bris, A.; Méléder, V.; Launeau, P.; Robin, M.; Louvrou, I.; Ribeiro, L. Photosynthetic epibionts and endobionts of Pacific oyster shells from oyster reefs in rocky versus mudflat shores. PLoS ONE 2017, 12, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Lizotte, M.P. The Contributions of Sea Ice Algae to Antarctic Marine Primary Production. Am. Zool. 2001, 41, 57–73. [Google Scholar] [CrossRef]
- Niner, H.J.; Ardron, J.A.; Escobar, E.G.; Gianni, M.; Jaeckel, A.; Jones, D.O.B.; Levin, L.A.; Smith, C.R.; Thiele, T.; Turner, P.J.; et al. Deep-sea mining with no net loss of biodiversity-an impossible aim. Front. Mar. Sci. 2018, 5, 53. [Google Scholar] [CrossRef] [Green Version]
- Da Ros, Z.; Dell’Anno, A.; Morato, T.; Sweetman, A.K.; Carreiro-Silva, M.; Smith, C.J.; Papadopoulou, N.; Corinaldesi, C.; Bianchelli, S.; Gambi, C.; et al. The deep sea: The new frontier for ecological restoration. Mar. Policy 2019, 108, 103642. [Google Scholar] [CrossRef]
- Simon-Lledó, E.; Bett, B.J.; Huvenne, V.A.I.; Köser, K.; Schoening, T.; Greinert, J.; Jones, D.O.B. Biological effects 26 years after simulated deep-sea mining. Sci. Rep. 2019, 9, 8040. [Google Scholar] [CrossRef]
- Dunn, D.C.; Van Dover, C.L.; Etter, R.J.; Smith, C.R.; Levin, L.A.; Morato, T.; Colaço, A.; Dale, A.C.; Gebruk, A.V.; Gjerde, K.M.; et al. A strategy for the conservation of biodiversity on mid-ocean ridges from deep-sea mining. Sci. Adv. 2018, 4, eaar4313. [Google Scholar] [CrossRef] [Green Version]
- Cochrane, S.K.J.; Ekehaug, S.; Pettersen, R.; Refit, E.C.; Hansen, I.M.; Aas, L.M.S. Detection of deposited drill cuttings on the sea floor—A comparison between underwater hyperspectral imagery and the human eye. Mar. Pollut. Bull. 2019, 145, 67–80. [Google Scholar] [CrossRef] [PubMed]
- Dierssen, H.; Bracher, A.; Brando, V.; Loisel, H.; Ruddick, K. Data needs for hyperspectral detection of algal diversity across the globe. Oceanography 2020, 33, 74–79. [Google Scholar] [CrossRef]
- Mendes, C.R.; Cartaxana, P.; Brotas, V. HPLC determination of phytoplankton and microphytobenthos pigments: Comparing resolution and sensitivity of a C18 and a C8 method. Limnol. Oceanogr. Methods 2007, 5, 363–370. [Google Scholar] [CrossRef]
- Jodłowska, S.; Latała, A. The Comparison of Spectrophotometric Method and High-Performance Liquid Chromatography in Photosynthetic Pigments Analysis. Online J. Biol. Sci. 2011, 11, 63–69. [Google Scholar]
- Thrane, J.E.; Kyle, M.; Striebel, M.; Haande, S.; Grung, M.; Rohrlack, T.; Andersen, T. Spectrophotometric analysis of pigments: A critical assessment of a high-throughput method for analysis of algal pigment mixtures by spectral deconvolution. PLoS ONE 2015, 10, e0137645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nelson, J.R.; Robertson, C.Y. Detrital spectral absorption: Laboratory studies of visible light effects on phytodetritus absorption, bacterial spectral signal, and comparison to field measurements. J. Mar. Res. 1993, 51, 181–207. [Google Scholar] [CrossRef]
- Rashid, A.R.; Chennu, A. A trillion coral reef colors: Deeply annotated underwater hyperspectral images for automated classification and habitat mapping. Data 2020, 5, 19. [Google Scholar] [CrossRef] [Green Version]
- Hedley, J.D.; Mirhakak, M.; Wentworth, A.; Dierssen, H.M. Influence of three-dimensional coral structures on hyperspectral benthic reflectance and water-leaving reflectance. Appl. Sci. 2018, 8, 2688. [Google Scholar] [CrossRef] [Green Version]
- Bao, J.; Li, D.; Qiao, X.; Rauschenbach, T. Integrated navigation for autonomous underwater vehicles in aquaculture: A review. Inf. Process. Agric. 2020, 7, 139–151. [Google Scholar] [CrossRef]
- Barker, L.D.L.; Jakuba, M.V.; Bowen, A.D.; German, C.R.; Maksym, T.; Mayer, L.; Boetius, A.; Dutrieux, P.; Whitcomb, L.L. Scientific challenges and present capabilities in underwater robotic vehicle design and navigation for oceanographic exploration under-ice. Remote Sens. 2020, 12, 2588. [Google Scholar] [CrossRef]
- Price, D.M.; Robert, K.; Callaway, A.; Lo lacono, C.; Hall, R.A.; Huvenne, V.A.I. Using 3D photogrammetry from ROV video to quantify cold-water coral reef structural complexity and investigate its influence on biodiversity and community assemblage. Coral Reefs 2019, 38, 1007–1021. [Google Scholar] [CrossRef] [Green Version]
- Savini, A.; Vertino, A.; Marchese, F.; Beuck, L.; Freiwald, A. Mapping cold-water coral habitats at different scales within the Northern Ionian Sea (central Mediterranean): An assessment of coral coverage and associated vulnerability. PLoS ONE 2014, 9, e87108. [Google Scholar] [CrossRef] [Green Version]
- Zwolak, K.; Wigley, R.; Bohan, A.; Zarayskaya, Y.; Bazhenova, E.; Dorshow, W.; Sumiyoshi, M.; Sattiabaruth, S.; Roperez, J.; Proctor, A.; et al. The autonomous underwater vehicle integrated with the unmanned surface vessel mapping the southern ionian sea. The winning technology solution of the shell ocean discovery XPRIZE. Remote Sens. 2020, 12, 1344. [Google Scholar] [CrossRef] [Green Version]
- Nocerino, E.; Menna, F.; Gruen, A.; Troyer, M.; Capra, A.; Castagnetti, C.; Rossi, P.; Brooks, A.J.; Schmitt, R.J.; Holbrook, S.J. Coral Reef Monitoring by Scuba Divers Using Underwater Photogrammetry and Geodetic Surveying. Remote Sens. 2020, 12, 3036. [Google Scholar] [CrossRef]
- Paoletti, M.E.; Haut, J.M.; Plaza, J.; Plaza, A. Deep learning classifiers for hyperspectral imaging: A review. ISPRS J. Photogramm. Remote Sens. 2019, 158, 279–317. [Google Scholar] [CrossRef]
- Schweiger, A.K.; Schütz, M.; Risch, A.C.; Kneubühler, M.; Haller, R.; Schaepman, M.E. How to predict plant functional types using imaging spectroscopy: Linking vegetation community traits, plant functional types and spectral response. Methods Ecol. Evol. 2017, 8, 86–95. [Google Scholar] [CrossRef]
- Laliberté, E.; Schweiger, A.K.; Legendre, P. Partitioning plant spectral diversity into alpha and beta components. Ecol. Lett. 2020, 23, 370–380. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| System/Platform | Achieved Transect Length (m) | Possible Survey Area (~m2) per Deployment | Spatial Resolution Achieved (cm/pix) | Distance to Target (m) | Deployment Depth (m) | Operation Mode | Reference |
|---|---|---|---|---|---|---|---|
| Underwater rail | 1–5 | 10 | 0.1 | 1 | 5–20 | MO from surface, A capacity | [26,27] |
| AUV | Not defined | 1 × 109 | 0.6 | 8.5 | 2300 | A | [26,70] |
| ROV | 1–20 | <500 | 0.1 | 1 | 30–4000 | MO from boat | [71,72,73] |
| USV | 1–20 | <500 | 0.5 | 1.5 | Surface | A | [67] |
| Under-ice slider | 10–30 | <40 | 0.1 | 1.2 | 1.5 | MO above ice | [68] |
| Diver units | 50 | 500–650 | 0.4 | 1 | 30 | MO underwater | [74] |
| Fixed stations | 1 | <2 | 0.1 | 1 | ~3500 | MO from boat, A capacity | [75] |
| Lab systems | 0.01 to 1 | N/A | 0.05 | <1 | - | MO or A | [27,72,76,77] |
| Study Focus(Marine Organism, Environment) | UHI Application | Platform | Wavelengths/Resolution | Data AnalysisMethod | Validation | Calibration | Ref |
|---|---|---|---|---|---|---|---|
| Microphytobenthos | Spectral Index for Chl a | Electric rail | 400–900 nm @ 1 nm | Regression: spectral index | Pigment extraction | Chl a spectrophotometry | [27] |
| Spectral Index for phycoerythrin | Diver-unit | 400–900 nm @ 1.5 nm | Regression: δδ (605 nm) | Visual ROI annotation | Uncalibrated | [74] | |
| Photosynthetic cell biomass | Electric rail | 400–1000 nm @ 1.3 nm | Regression: 3rd end-member spectrum | Visual ROI annotation | Uncalibrated | [80] | |
| Warm water corals | Benthic classification | Diver-unit | 400–900 nm @ 1.5 nm | Regression: δδ (580, 675 nm) | Visual RGB | Uncalibrated | [74] |
| Colony bleaching assessment | In vitro rotational stage | 400–1000 nm @ 2.2 nm | Object fluorescence emission spectra | Visual RGB | Spectrometer | [101] | |
| Physiological interactions | In vitro electric rail | 450–900 nm @ not specified | GOC515-575-685 CIR550-650-860 NDVI800-680 ARVI800-680-450 | Visual RGB and oxygen profiling | Uncalibrated | [100] | |
| Cold water corals | Polyp mortality classification | In vitro electric rail | 381–846 nm @ 1 nm | Classification: v-SVM | Visual RGB | Visual inspection | [77] |
| Benthic classification | ROV | 380–800 nm @ 15 nm | Classification: SAM | Visual RGB | N/A | [73] | |
| Coralline algae | Bio-optical taxonomic tool | In vitro electric rail | 400–700 nm @ 2 nm | Classification: SAM | Pigment extraction | Spectrophotometry and HPLC | [72] |
| Benthic classification | ROV | 400–700 nm @ 2 nm | Classification: SAM, MD, BE, SID, Pp | Visual ROI annotation | Spectrophotometry and HPLC | [72] | |
| Benthic classification | ROV | 380–800 nm @ 15 nm | Classification: SAM | Visual ROI annotation | N/A | [73] | |
| Classification of photo-epibionts | In vitro tripod system | 400–1000 nm @ 4.5 nm | Regression: δδ (546, 568, 648, 677 nm) | Pigment extraction | HPLC | [108] | |
| Sponges | Bio-optical taxonomic tool | In vitro electric rail | 420–680 nm @ 1 nm | Classification: object reflectance spectra | Pigment extraction | HPLC & mass spectrophotometry | [64] |
| Benthic classification | ROV | 378–805 nm @ 4 nm | Classification: SVM | Visual ROI annotation | N/A | [71] | |
| Benthic classification | ROV | 380–800 nm @ 15 nm | Classification: SAM | Visual ROI annotation | N/A | [73] | |
| Oyster reefs | Classification of photo-bionts | In vitro electric rail | 400–950 nm @ 4.5 nm | Regression: NDVI750-673, δδ (462, 524, 571, 647 nm) | Pigment extraction | HPLC | [113] |
| Sympagic environments | Proxy of ice-algae biomass distribution | In vitro electric rail | 400–1000 nm @ 1.7, 3.4, 6.8 nm | PCA | Pigment extraction | Uncalibrated | [66] |
| Proxies of ice-algae biomass distribution | In situ under-ice sled | 400–1000 nm @ 3.5 nm | PCA NDI441-426, NDI648-567, ANMB650-700 | Visual ROI verification | Uncalibrated | [68] | |
| Quantitative estimates of biomass via spectral indices for Chl a | In situ under-ice sled and ex situ electric rail | 400–1000 nm @ 1.7 nm | Regression: NDI, AUC650-700, ANCB650-700, ANMB650-700, LAUC650-700 | Pigment extraction | Fluorometer | [76] | |
| Mineral resource assessment areas | Bio-optical taxonomic tool | ROV | 400–710 nm @ 4 nm | Classification: SVM | Visual ROI annotation | N/A | [71] |
| Sediment deposition homogeneity | ROV | 400–700 nm @ 5 nm | Regression: PCA and singular-value decomposition | van-Veen grab | N/A | [119] | |
| Benthic classification | Stationary platform | 400–730 nm @ 2 nm | Classification: SVM | Visual ROI annotation | N/A | [75] |
| Application | Microphytobenthos | Warm-Water Corals | Cold-Water Corals | Coralline Algae | Sponges | Oyster Reefs | Sympagic Environments | Mineral Resource Areas |
|---|---|---|---|---|---|---|---|---|
| Photosynthetic pigment content | Demonstrated | Lacking validation | N/A | Demonstrated | N/A | N/A | Demonstrated | Lacking validation |
| Species identification | Unproven | Demonstrated | Demonstrated | Unproven | Demonstrated | Unproven | Unproven | Demonstrated |
| Physiological assessments | N/A | Demonstrated | Demonstrated | Unproven | Unproven | N/A | Unproven | Unproven |
| In situ abundance | Demonstrated | Demonstrated | Demonstrated | Demonstrated | Demonstrated | Unproven | Demonstrated | Demonstrated |
| Epiphyte composition | N/A | Unproven | N/A | Demonstrated | Unproven | Demonstrated | N/A | N/A |
| Nutrient cycling | Demonstrated | Unproven | Unproven | Unproven | Unproven | Unproven | Unproven | Unproven |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Montes-Herrera, J.C.; Cimoli, E.; Cummings, V.; Hill, N.; Lucieer, A.; Lucieer, V. Underwater Hyperspectral Imaging (UHI): A Review of Systems and Applications for Proximal Seafloor Ecosystem Studies. Remote Sens. 2021, 13, 3451. https://doi.org/10.3390/rs13173451
Montes-Herrera JC, Cimoli E, Cummings V, Hill N, Lucieer A, Lucieer V. Underwater Hyperspectral Imaging (UHI): A Review of Systems and Applications for Proximal Seafloor Ecosystem Studies. Remote Sensing. 2021; 13(17):3451. https://doi.org/10.3390/rs13173451
Chicago/Turabian StyleMontes-Herrera, Juan C., Emiliano Cimoli, Vonda Cummings, Nicole Hill, Arko Lucieer, and Vanessa Lucieer. 2021. "Underwater Hyperspectral Imaging (UHI): A Review of Systems and Applications for Proximal Seafloor Ecosystem Studies" Remote Sensing 13, no. 17: 3451. https://doi.org/10.3390/rs13173451
APA StyleMontes-Herrera, J. C., Cimoli, E., Cummings, V., Hill, N., Lucieer, A., & Lucieer, V. (2021). Underwater Hyperspectral Imaging (UHI): A Review of Systems and Applications for Proximal Seafloor Ecosystem Studies. Remote Sensing, 13(17), 3451. https://doi.org/10.3390/rs13173451