
Estimating Wildlife Density as a Function of Environmental Heterogeneity Using Unmarked Data


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
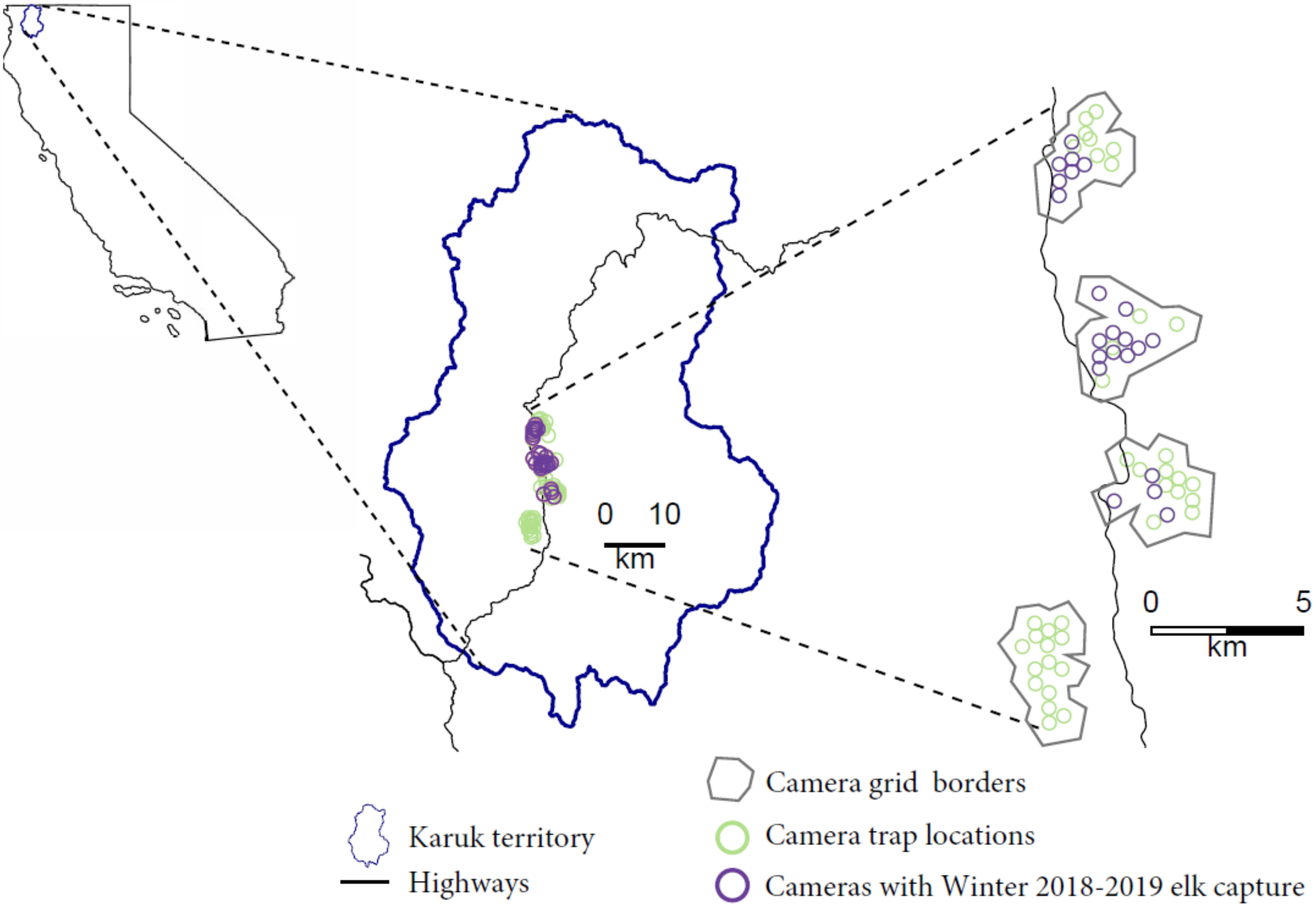
Study System
- Camera trapping
- 2.
- Elk spatial presence/counts
- 3.
- GPS collaring
- 4.
- Unmarked spatial capture recapture model
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Burton, A.C.; Neilson, E.; Moreira, D.; Ladle, A.; Steenweg, R.; Fisher, J.T.; Bayne, E.; Boutin, S. Wildlife camera trapping: A review and recommendations for linking surveys to ecological processes. J. Appl. Ecol. 2015, 52, 675–685. [Google Scholar] [CrossRef]
- De Bondi, N.; White, J.G.; Stevens, M.; Cooke, R. A comparison of the effectiveness of camera trapping and live trapping for sampling terrestrial small-mammal communities. Wildl. Res. 2010, 37, 456–465. [Google Scholar] [CrossRef]
- Royle, J.A.; Karanth, K.U.; Gopalaswamy, A.M.; Kumar, N.S. Bayesian inference in camera trapping studies for a class of spatial capture-recapture models. Ecology 2009, 90, 3233–3244. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, N.A.; Clare, J.D.; Stenglein, J.L.; Zuckerberg, B. Abundance estimation methods for unmarked animals with camera traps. Conserv. Biol. 2020, 35, 88–100. [Google Scholar] [CrossRef]
- Chandler, R.B.; Royle, J.A. Spatially explicit models for inference about density in unmarked or partially marked populations. Ann. Appl. Stat. 2013, 7, 936–954. [Google Scholar] [CrossRef]
- Efford, M.G.; Fewster, R.M. Estimating population size by spatially explicit capture-recapture. Oikos 2013, 122, 918–928. [Google Scholar] [CrossRef]
- Evans, M.J.; Rittenhouse, T.A.G. Evaluating spatially explicit density estimates of unmarked wildlife detected by remote cameras. J. Appl. Ecol. 2018, 55, 2565–2574. [Google Scholar] [CrossRef]
- Ramsey, D.S.L.; Caley, P.A.; Robley, A. Estimating Population Density From Presence-Absence Data Using a Spatially Explicit Model. J. Wildl. Manag. 2015, 79, 491–499. [Google Scholar] [CrossRef]
- KDNR. Eco-Cultural Resource Management Plan; KDNR: Orleans, CA, USA, 2011; pp. 69–70. [Google Scholar]
- Allison, B.L.; Creasy, M.; Ford, M.; Hacking, A.; Schaefer, R.; West, J.; Youngblood, Q. Elk Management Strategy Klamath National Forest; Klamath National Forest: Orleans, CA, USA, 2007. [Google Scholar]
- Taylor, A.H.; Skinner, C.N. Spatial patterns and controls on historical fire regimes and forest structure in the Klamath Mountains. Ecol. Appl. 2003, 13, 704–719. [Google Scholar] [CrossRef]
- Sawyer, J.O. Northwest California: A Natural History; University of California Press: Berkeley, CA, USA, 2006; pp. 1–250. [Google Scholar]
- Norgaard, K. The Politics of Fire and the Social Impacts of Fire Exclusion on the Klamath. Humboldt J. Soc. Relat. 2014, 36, 77–101. [Google Scholar]
- Harper, J.A.; Harn, J.H.; Bentley, W.W.; Yocom, C.F. The status and ecology of the Roosevelt elk in California. Wildl. Monogr. 1967, 16, 3–49. [Google Scholar]
- USFS. Somes Bar Integrated Fire Management Project: Environmental Assessment; USFS: Orleans, CA, USA, 2018. [Google Scholar]
- Niedballa, J.; Sollmann, R.; Courtiol, A.; Wilting, A. camtrapR: An R package for efficient camera trap data management. Methods Ecol. Evol. 2016, 7, 1457–1462. [Google Scholar] [CrossRef]
- Calenge, C. The package “adehabitat” for the R software: A tool for the analysis of space and habitat use by animals. Ecol. Model. 2006, 197, 516–519. [Google Scholar] [CrossRef]
- Efford, M. Secr 3.2-Spatially Explicit Capture–Recapture in R; Dunedin, NZ, USA. 2019. Available online: https://cran.r-project.org/web/packages/secr/index.html (accessed on 4 January 2022).
- Royle, J.A.; Dorazio, R.M. Parameter-expanded data augmentation for Bayesian analysis of capture-recapture models. J. Ornithol. 2012, 152, S521–S537. [Google Scholar] [CrossRef]
- Royle, J.A.; Young, K.V. A hierarchical model for spatial capture-recapture data. Ecology 2008, 89, 2281–2289. [Google Scholar] [CrossRef]
- Jones, M.O.; Allred, B.W.; Naugle, D.E.; Maestas, J.D.; Donnelly, P.; Metz, L.J.; Karl, J.; Smith, R.; Bestelmeyer, B.; Boyd, C.; et al. Innovation in rangeland monitoring: Annual, 30 m, plant functional type percent cover maps for US rangelands, 1984–2017. Ecosphere 2018, 9, e02430. [Google Scholar] [CrossRef]
- De Valpine, P.; Turek, D.; Paciorek, C.J.; Anderson-Bergman, C.; Lang, D.T.; Bodik, R. Programming with Models: Writing Statistical Algorithms for General Model Structures with NIMBLE. J. Comput. Graph. Stat. 2017, 26, 403–413. [Google Scholar] [CrossRef] [Green Version]
- R Core Development Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Hooten, M.B.; Hobbs, N.T. A guide to Bayesian model selection for ecologists. Ecol. Monogr. 2015, 85, 3–28. [Google Scholar] [CrossRef]
- Lambin, E.F. Modelling and monitoring land-cover change processes in tropical regions. Prog. Phys. Geogr. 1997, 21, 375–393. [Google Scholar] [CrossRef]
- Cushman, S.A.; Landguth, E.L. Scale dependent inference in landscape genetics. Landsc. Ecol. 2010, 25, 967–979. [Google Scholar] [CrossRef]
- Houlahan, J.E.; McKinney, S.T.; Anderson, T.M.; McGill, B.J. The priority of prediction in ecological understanding. Oikos 2017, 126, 1–7. [Google Scholar] [CrossRef]
- Horncastle, V.J.; Yarborough, R.F.; Dickson, B.G.; Rosenstock, S.S. Summer Habitat Use by Adult Female Mule Deer in a Restoration-Treated Ponderosa Pine Forest. Wildl. Soc. Bull. 2013, 37, 707–713. [Google Scholar] [CrossRef]
- Porter, W.P.; Sabo, J.L.; Tracy, C.R.; Reichman, O.J.; Ramankutty, N. Physiology on a landscape scale: Plant-animal interactions. Integr. Comp. Biol. 2002, 42, 431–453. [Google Scholar] [CrossRef] [PubMed]
- Parker, K.L.; Gillingham, M.P. Estimates of critical thermal environments for mule deer. J. Range Manag. 1990, 43, 73–81. [Google Scholar] [CrossRef]
- Cook, J.G.; Irwin, L.L.; Bryant, L.D.; Riggs, R.A.; Thomas, J.W. Relations of forest cover and condition of elk: A test of the thermal cover hypothesis in summer and winter. Wildl. Monogr. 1998, 141, 5–61. [Google Scholar]
- Dey, S.; Delampady, M.; Gopalaswamy, A.M. Bayesian model selection for spatial capture-recapture models. Ecol. Evol. 2019, 9, 11569–11583. [Google Scholar] [CrossRef]
- Gardner, B.; Royle, J.A.; Wegan, M.T.; Rainbolt, R.E.; Curtis, P.D. Estimating Black Bear Density Using DNA Data From Hair Snares. J. Wildl. Manag. 2010, 74, 318–325. [Google Scholar] [CrossRef]
- Sutherland, C.; Royle, J.A.; Linden, D.W. oSCR: A spatial capture-recapture R package for inference about spatial ecological processes. Ecography 2019, 42, 1459–1469. [Google Scholar] [CrossRef] [Green Version]
- Bailey, L.L.; Simons, T.R.; Pollock, K.H. Spatial and temporal variation in detection probability of plethodon salamanders using the robust capture-recapture design. J. Wildl. Manag. 2004, 68, 14–24. [Google Scholar] [CrossRef]
- Furnas, B.J.; Landers, R.H.; Hill, S.; Itoga, S.S.; Sacks, B.N. Integrated modeling to estimate population size and composition of mule deer. J. Wildl. Manag. 2018, 82, 1429–1441. [Google Scholar] [CrossRef]
- Royle, J.A.; Nichols, J.D. Estimating abundance from repeated presence-absence data or point counts. Ecology 2003, 84, 777–790. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Parameter | Mean (Probability of Inclusion in Parentheses, If Applicable *) | Standard Deviation | 95% Lower CI | 95% Upper CI | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Independent Detection Count | Total Individual Count | Presence-Absence | Independent Detection Count | Total Individual Count | Presence-Absence | Independent Detection Count | Total Individual Count | Presence-Absence | Independent Detection Count | Total Individual Count | Presence-Absence | |
| g0 | 0.100 | 0.340 | 0.050 | 0.023 | 0.099 | 0.028 | 0.062 | 0.142 | 0.023 | 0.149 | 0.534 | 0.125 |
| sigma | 0.264 | 0.225 | 1.09 | 0.024 | 0.017 | 0.682 | 0.219 | 0.201 | 0.227 | 0.313 | 0.269 | 3.115 |
| Density | 0.463 | 0.566 | 0.109 | 0.167 | 0.157 | 0.191 | 0.206 | 0.301 | 0.003 | 0.858 | 0.907 | 0.838 |
| FG cover | 0.052 (0.73) | 0.076 (0.908) | −0.035 (0.529) | 0.045 | 0.027 | 0.047 | −0.026 | 0.015 | −0.121 | 0.130 | 0.125 | 0.066 |
| Percent tree cover | −0.081 (0.81) | −0.116 (0.991) | −0.065 (0.712) | 0.066 | 0.046 | 0.053 | −0.199 | −0.194 | −0.155 | 0.043 | −0.017 | 0.070 |
| Elevation | −0.004 (0.17) | 1.2 × 10−4 (0.003) | −4.9 × 10−4 (0.010) | 0.003 | 3.0 × 10−4 | 7.2 × 10−4 | −0.011 | −0.001 | −0.002 | 0.001 | 3.8 × 10−4 | 0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Connor, T.; Division, W.; Tripp, E.; Bean, W.T.; Saxon, B.J.; Camarena, J.; Donahue, A.; Sarna-Wojcicki, D.; Macaulay, L.; Tripp, W.; et al. Estimating Wildlife Density as a Function of Environmental Heterogeneity Using Unmarked Data. Remote Sens. 2022, 14, 1087. https://doi.org/10.3390/rs14051087
Connor T, Division W, Tripp E, Bean WT, Saxon BJ, Camarena J, Donahue A, Sarna-Wojcicki D, Macaulay L, Tripp W, et al. Estimating Wildlife Density as a Function of Environmental Heterogeneity Using Unmarked Data. Remote Sensing. 2022; 14(5):1087. https://doi.org/10.3390/rs14051087
Chicago/Turabian StyleConnor, Thomas, Wildlife Division, Emilio Tripp, William T. Bean, B. J. Saxon, Jessica Camarena, Asa Donahue, Daniel Sarna-Wojcicki, Luke Macaulay, William Tripp, and et al. 2022. "Estimating Wildlife Density as a Function of Environmental Heterogeneity Using Unmarked Data" Remote Sensing 14, no. 5: 1087. https://doi.org/10.3390/rs14051087