
Synthetic Aperture Radar Monitoring of Snow in a Reindeer-Grazing Landscape



Abstract
:1. Introduction
2. Study Area and Datasets
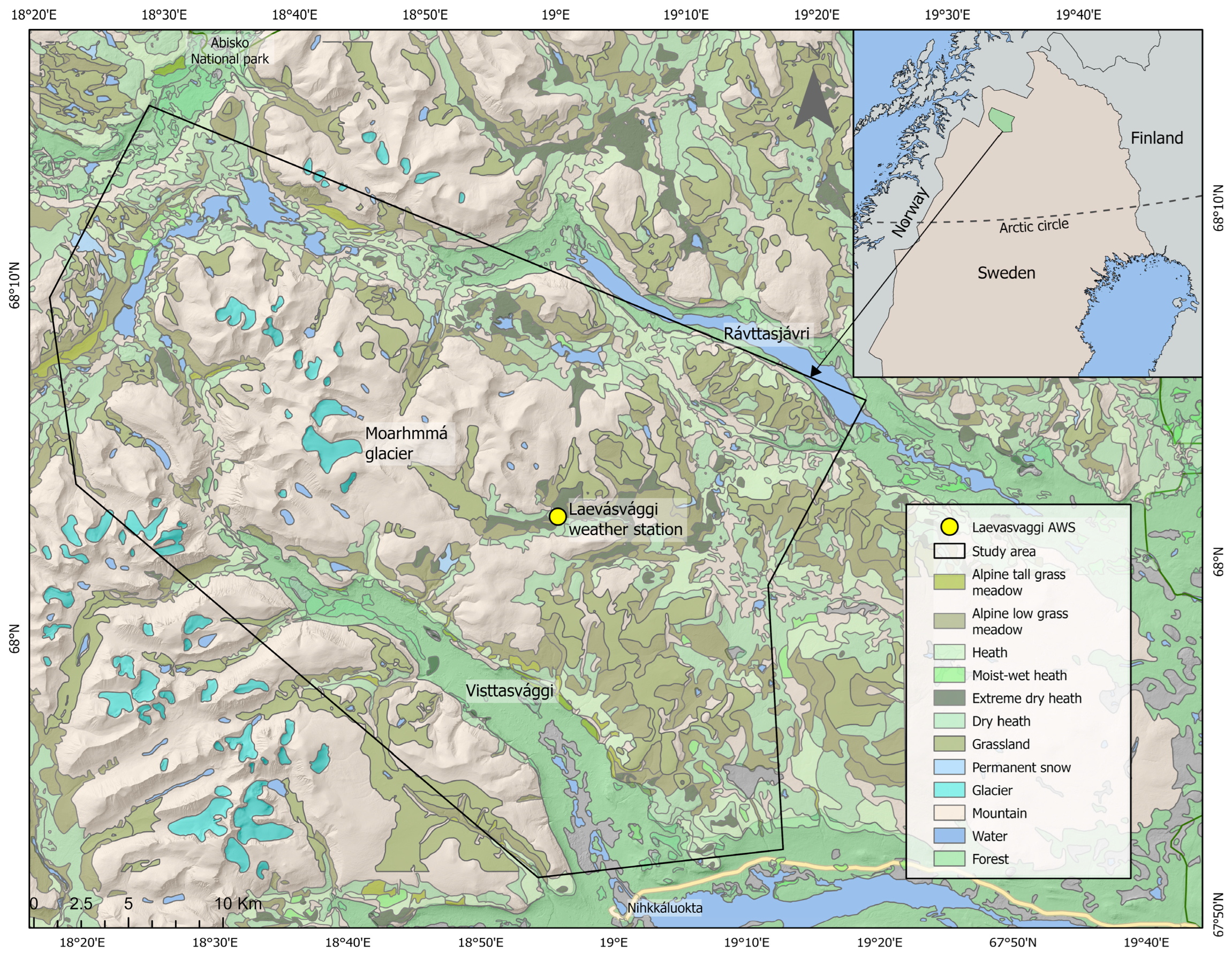
2.1. Study Area
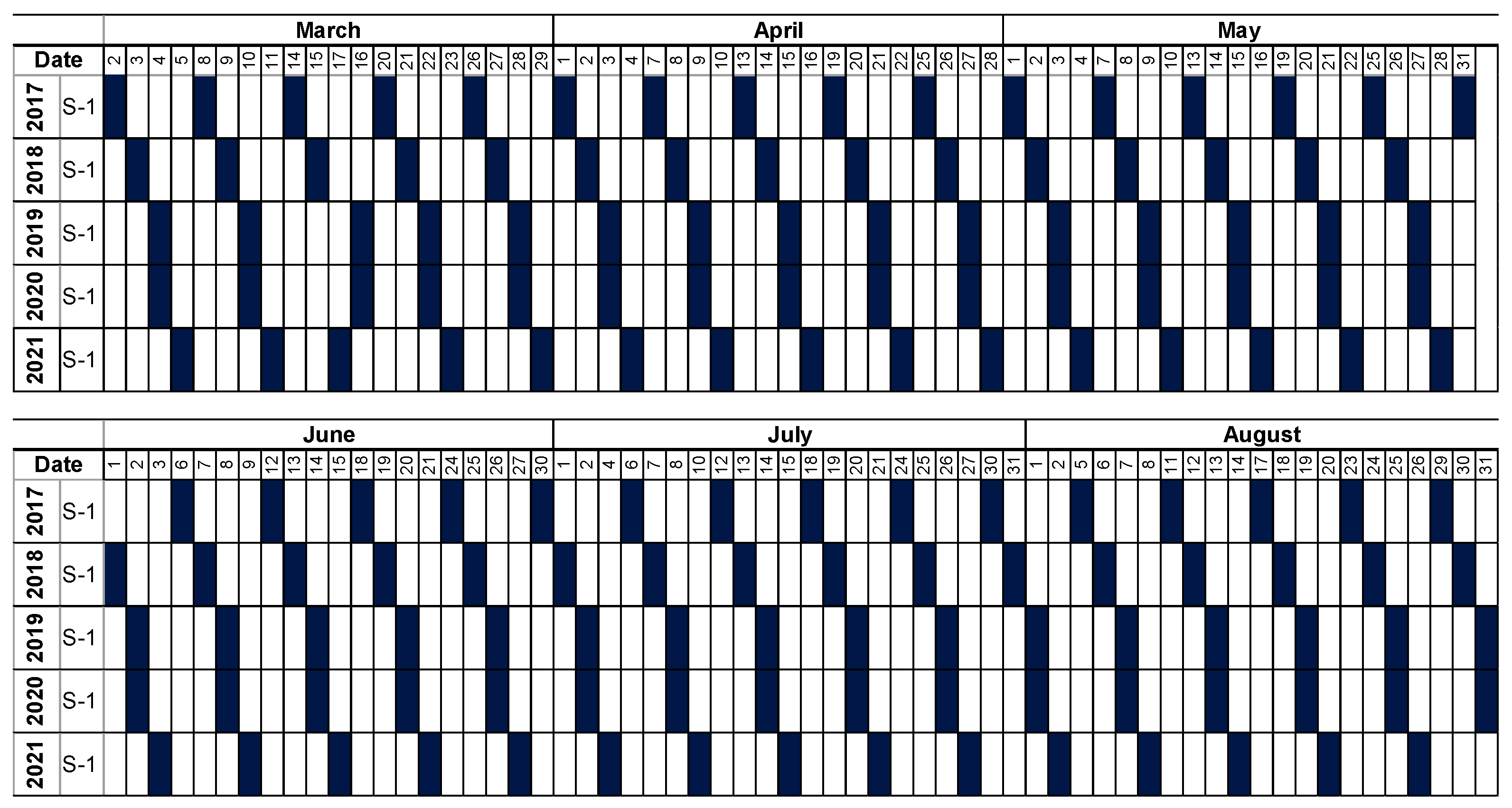
2.2. Datasets
3. Method
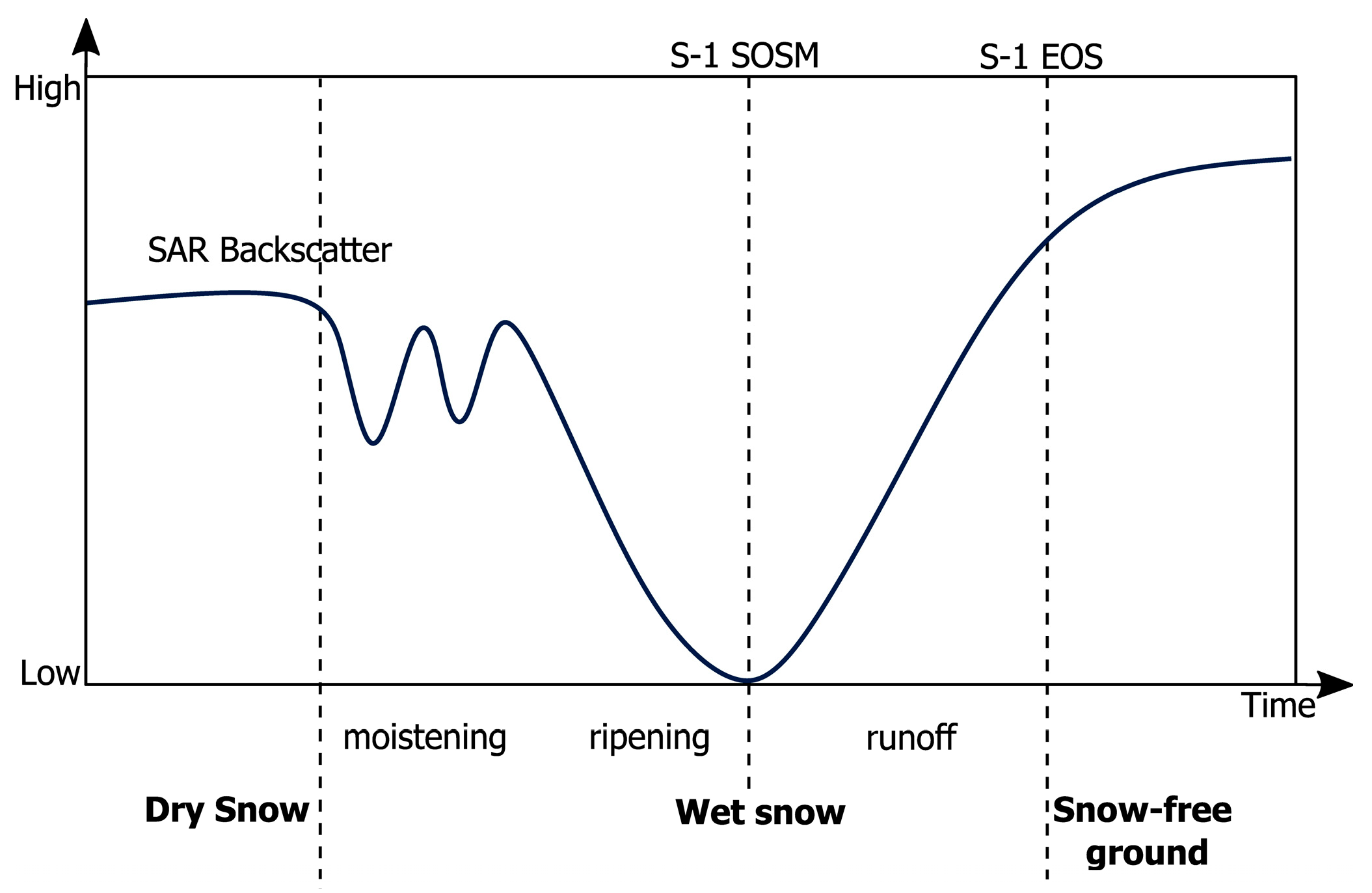
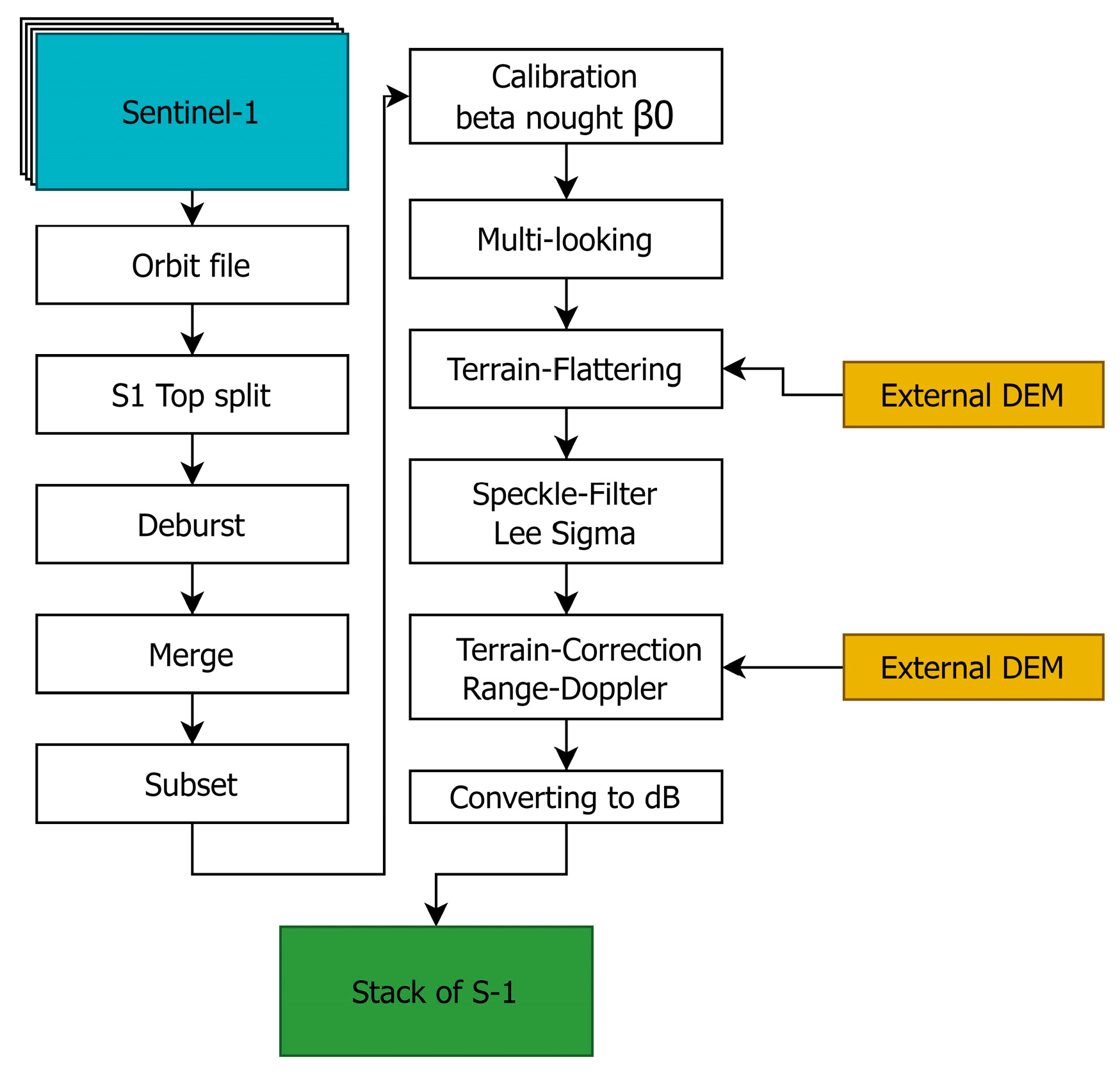
3.1. Pre-Processing of Sentinel-1 Data
3.2. Extracting SOSM and EOS Using Python
4. Results
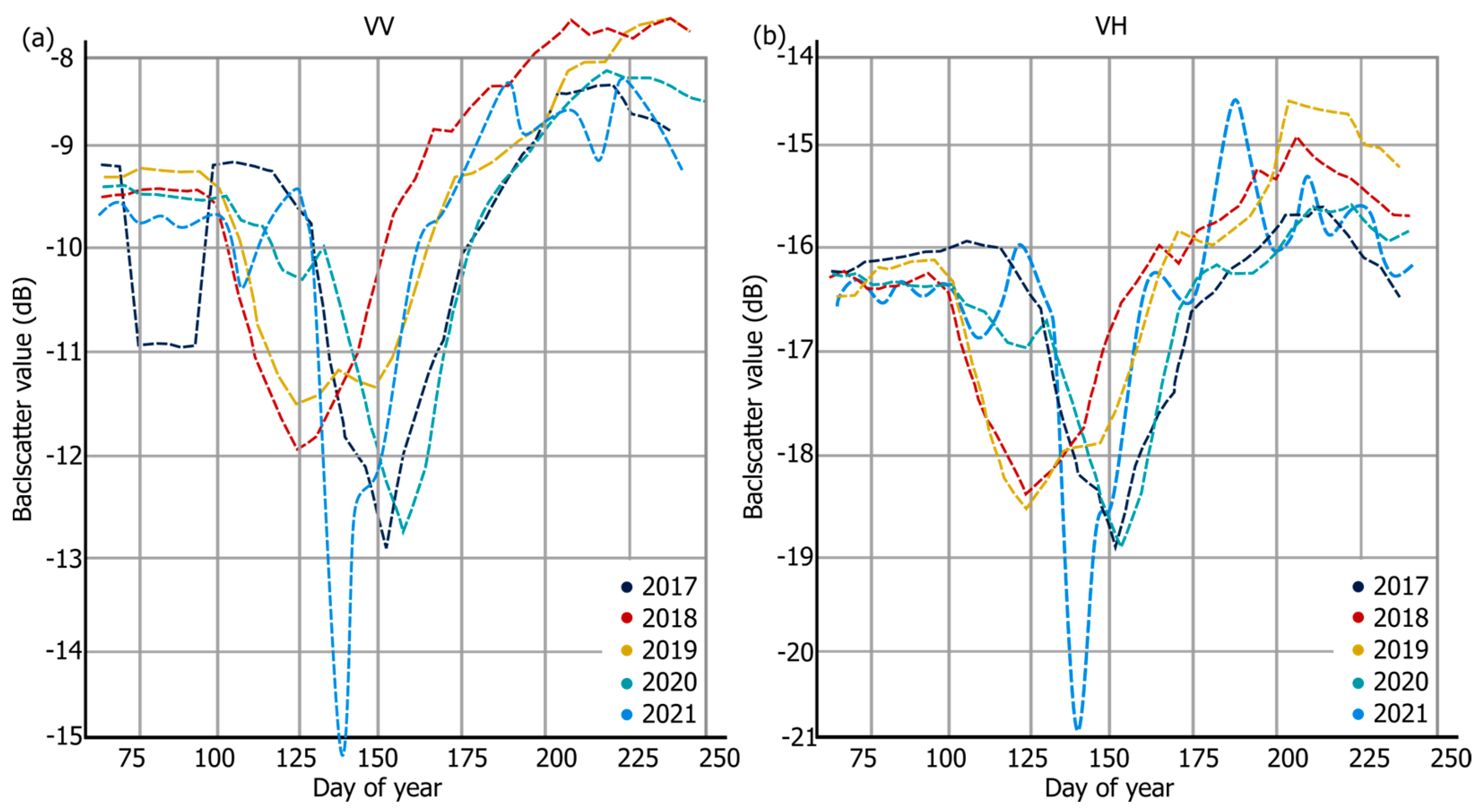
4.1. Backscatter Behaviour between the Different Polarisations and Years
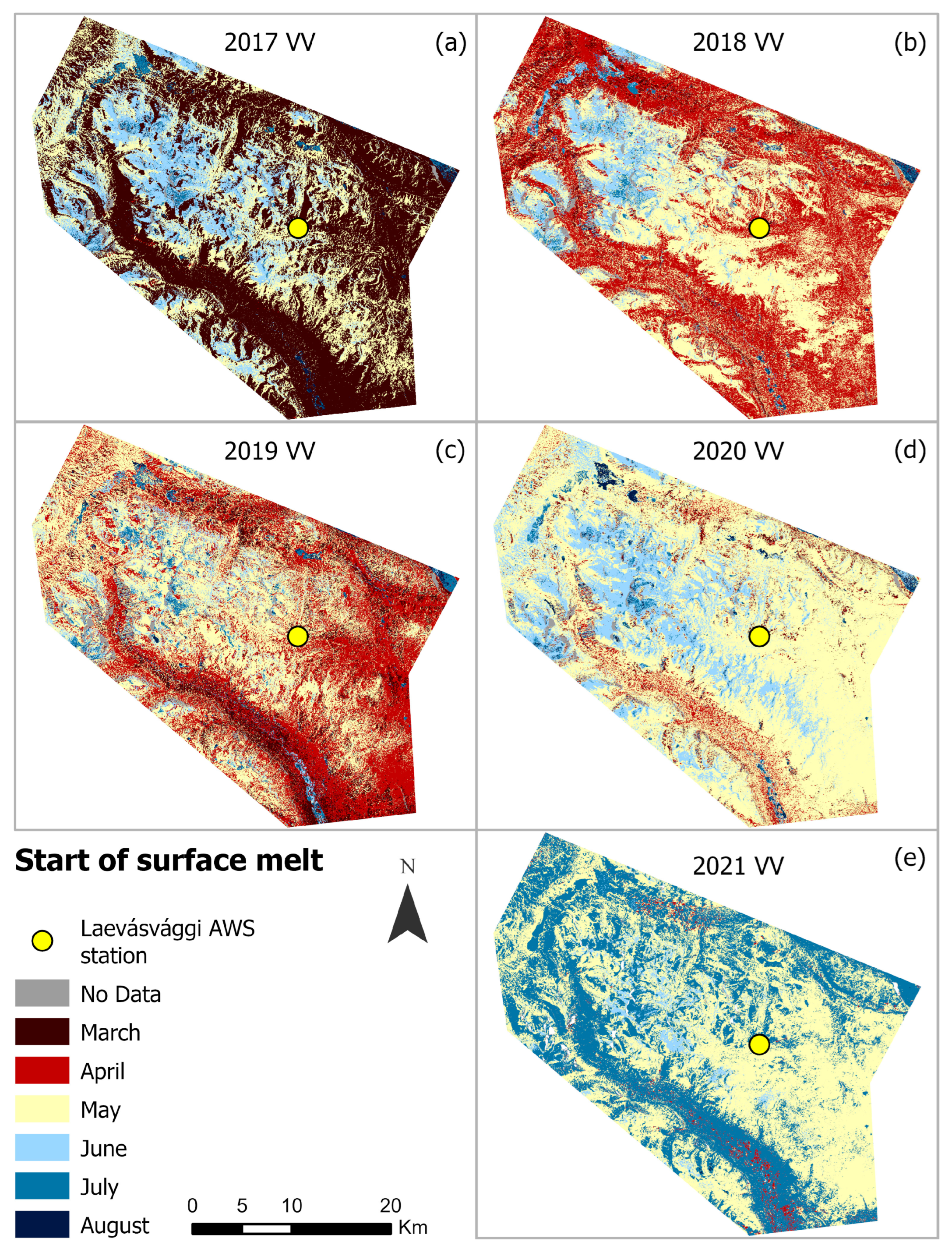
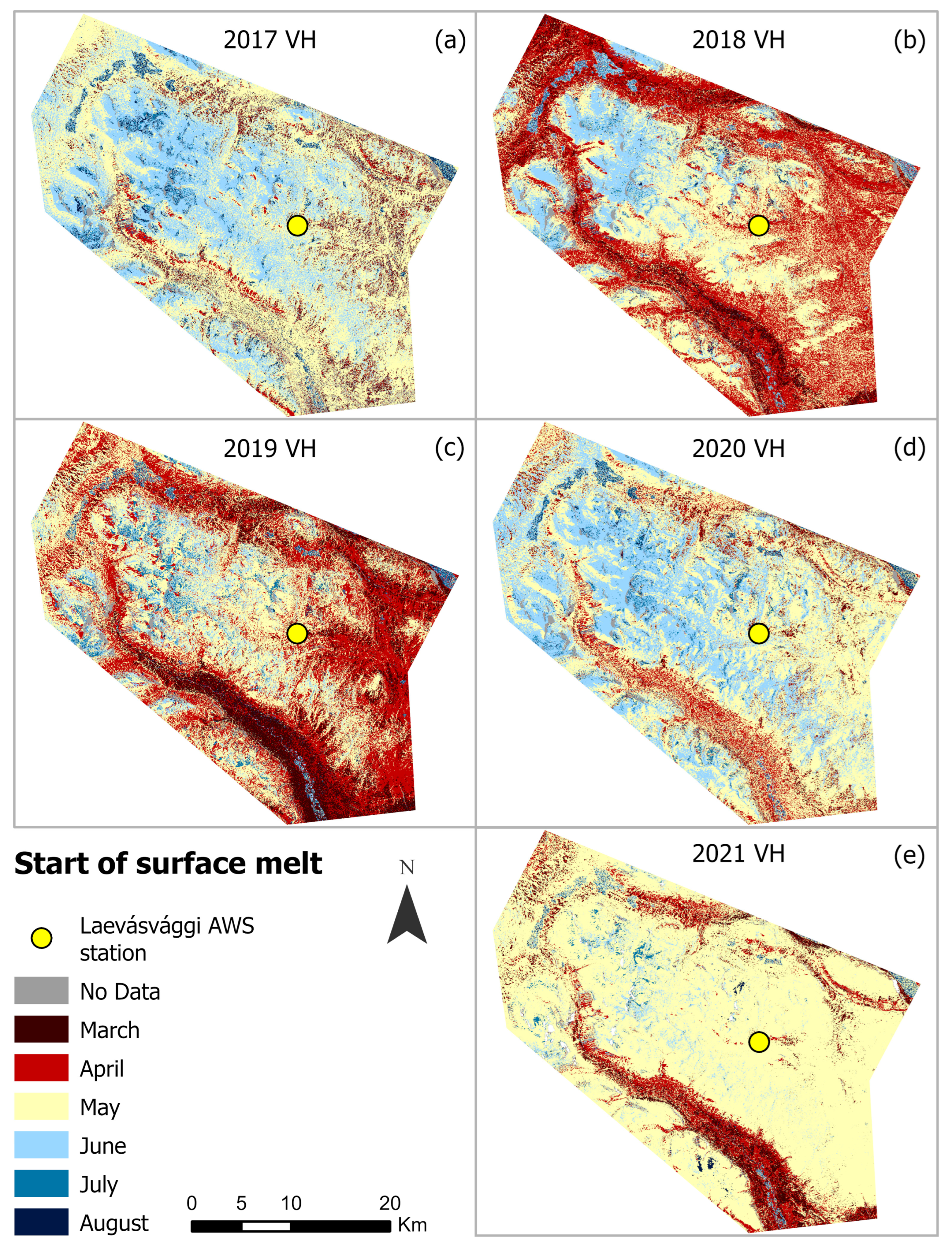
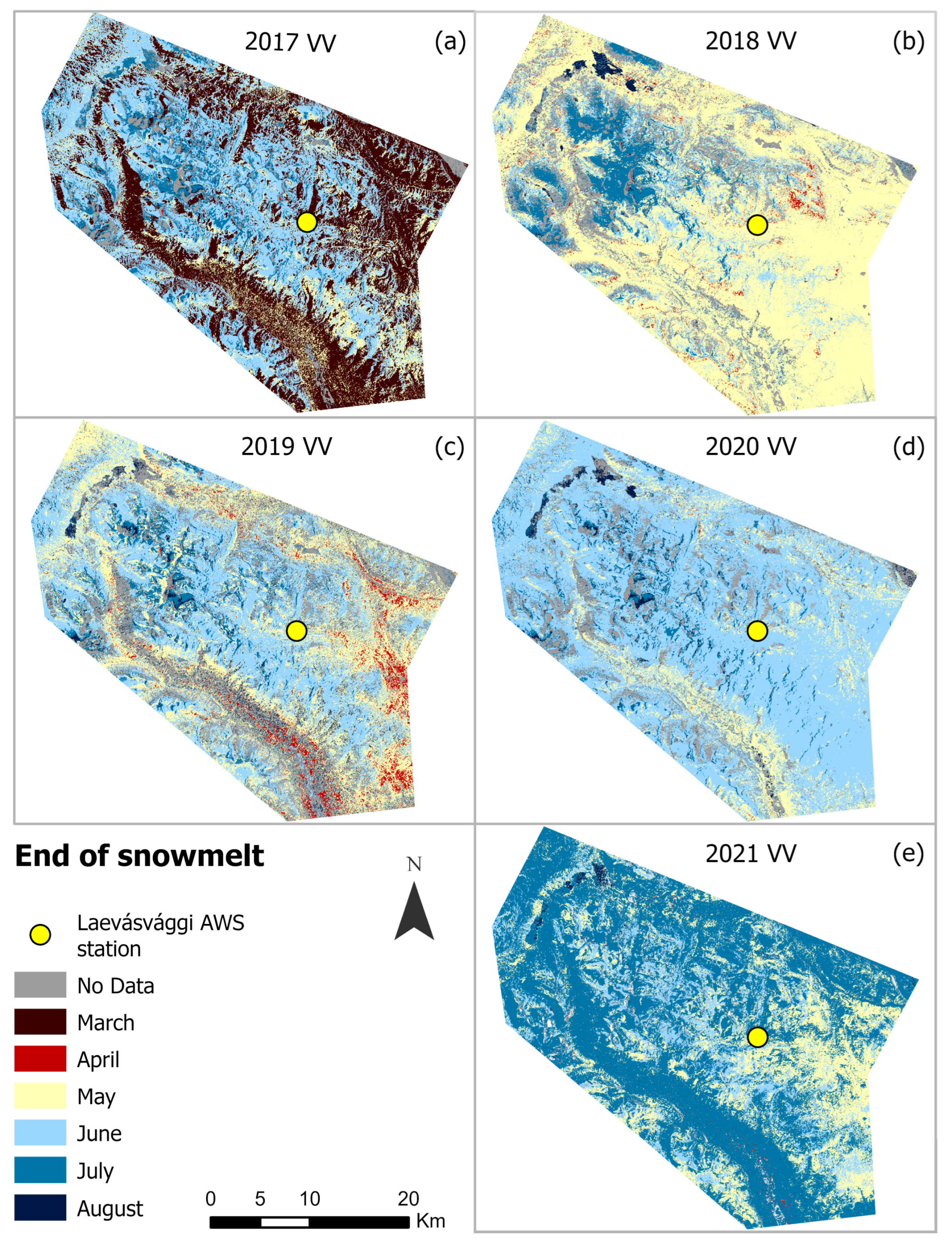
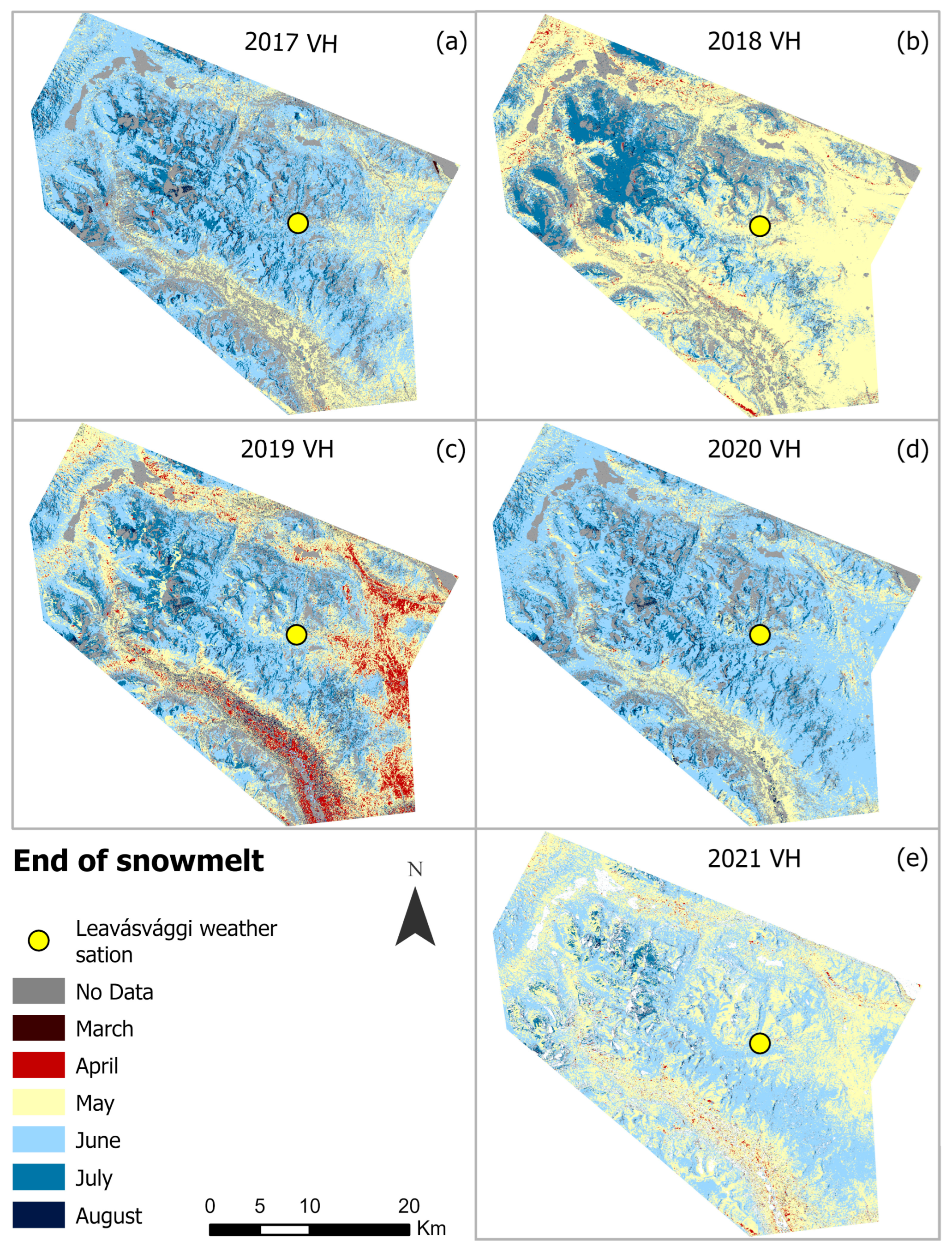
4.2. Seasonal Variations in SOSM and EOS from Sentinel-1 (2017–2021)
4.3. Sentinel-1 and In Situ Data from Laevásvággi AWS
4.4. Differences between Landcover Classes
5. Discussion
5.1. Variation in Backscattering and Spring Snowmelt
5.2. Impact of Variation in Spring Snowmelt on Vegetation and Reindeer
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- IPCC. Climate Change 2023: Synthesis Report. In A Report of the Intergovernmental Panel on Climate Change. Contribution of Working Groups I, II and III to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; IPCC: Geneva, Switzerland, 2023; pp. 1–85. [Google Scholar]
- Coumou, D.; Di Capua, G.; Vavrus, S.; Wang, L.; Wang, S. The Influence of Arctic Amplification on Mid-Latitude Summer Circulation. Nat. Commun. 2018, 9, 2959. [Google Scholar] [CrossRef] [PubMed]
- Furberg, M.; Evengård, B.; Nilsson, M. Facing the Limit of Resilience: Perceptions of Climate Change among Reindeer Herding Sami in Sweden. Glob. Health Action 2011, 4, 8417. [Google Scholar] [CrossRef] [PubMed]
- Kivinen, S.; Rasmus, S. Observed Cold Season Changes in a Fennoscandian Fell Area over the Past Three Decades. AMBIO 2015, 44, 214–225. [Google Scholar] [CrossRef] [PubMed]
- Rosqvist, G.C.; Inga, N.; Eriksson, P. Impacts of Climate Warming on Reindeer Herding Require New Land-Use Strategies. AMBIO 2022, 51, 1247–1262. [Google Scholar] [CrossRef] [PubMed]
- Pascual, D.; Åkerman, J.; Becher, M.; Callaghan, T.V.; Christensen, T.R.; Dorrepaal, E.; Emanuelsson, U.; Giesler, R.; Hammarlund, D.; Hanna, E.; et al. The Missing Pieces for Better Future Predictions in Subarctic Ecosystems: A Torneträsk Case Study. AMBIO 2021, 50, 375–392. [Google Scholar] [CrossRef] [PubMed]
- Wipf, S.; Rixen, C. A Review of Snow Manipulation Experiments in Arctic and Alpine Tundra Ecosystems. Polar Res. 2010, 29, 95–109. [Google Scholar] [CrossRef]
- Kater, I.; Baxter, R. Abundance and Accessibility of Forage for Reindeer in Forests of Northern Sweden: Impacts of Landscape and Winter Climate Regime. Ecol. Evol. 2022, 12, e8820. [Google Scholar] [CrossRef] [PubMed]
- Wipf, S.; Sommerkorn, M.; Stutter, M.I.; Wubs, E.R.J.; Van Der Wal, R. Snow Cover, Freeze-thaw, and the Retention of Nutrients in an Oceanic Mountain Ecosystem. Ecosphere 2015, 6, 1–16. [Google Scholar] [CrossRef]
- Rixen, C.; Høye, T.T.; Macek, P.; Aerts, R.; Alatalo, J.M.; Anderson, J.T.; Arnold, P.A.; Barrio, I.C.; Bjerke, J.W.; Björkman, M.P.; et al. Winters Are Changing: Snow Effects on Arctic and Alpine Tundra Ecosystems. Arct. Sci. 2022, 8, 572–608. [Google Scholar] [CrossRef]
- Bokhorst, S.; Pedersen, S.H.; Brucker, L.; Anisimov, O.; Bjerke, J.W.; Brown, R.D.; Ehrich, D.; Essery, R.L.H.; Heilig, A.; Ingvander, S.; et al. Changing Arctic Snow Cover: A Review of Recent Developments and Assessment of Future Needs for Observations, Modelling, and Impacts. AMBIO 2016, 45, 516–537. [Google Scholar] [CrossRef]
- Bosiö, J.; Stiegler, C.; Johansson, M.; Mbufong, H.N.; Christensen, T.R. Increased Photosynthesis Compensates for Shorter Growing Season in Subarctic Tundra—8 Years of Snow Accumulation Manipulations. Clim. Chang. 2014, 127, 321–334. [Google Scholar] [CrossRef]
- Kelsey, K.C.; Pedersen, S.H.; Leffler, A.J.; Sexton, J.O.; Feng, M.; Welker, J.M. Winter Snow and Spring Temperature Have Differential Effects on Vegetation Phenology and Productivity across Arctic Plant Communities. Glob. Chang. Biol. 2021, 27, 1572–1586. [Google Scholar] [CrossRef] [PubMed]
- Walker, M.D.; Walker, D.A.; Welker, J.M.; Arft, A.M.; Bardsley, T.; Brooks, P.D.; Fahnestock, J.T.; Jones, M.H.; Losleben, M.; Parsons, A.N.; et al. Long-Term Experimental Manipulation of Winter Snow Regime and Summer Temperature in Arctic and Alpine Tundra. Hydrol. Process. 1999, 13, 2315–2330. [Google Scholar] [CrossRef]
- Gamon, J.A.; Huemmrich, K.F.; Stone, R.S.; Tweedie, C.E. Spatial and Temporal Variation in Primary Productivity (NDVI) of Coastal Alaskan Tundra: Decreased Vegetation Growth Following Earlier Snowmelt. Remote Sens. Environ. 2013, 129, 144–153. [Google Scholar] [CrossRef]
- Bilbrough, C.J.; Welker, J.M.; Bowman, W.D. Early Spring Nitrogen Uptake by Snow-Covered Plants: A Comparison of Arctic and Alpine Plant Function under the Snowpack. Arct. Antarct. Alp. Res. 2000, 32, 404–411. [Google Scholar] [CrossRef]
- Li, Y.; He, H.S.; Zong, S.; Sun, H. Spring Snowmelt Affects Changes of Alpine Tundra Vegetation in Changbai Mountains. Ecohydrology 2022, 15, e2447. [Google Scholar] [CrossRef]
- Semenchuk, P.R.; Gillespie, M.A.K.; Rumpf, S.B.; Baggesen, N.; Elberling, B.; Cooper, E.J. High Arctic Plant Phenology Is Determined by Snowmelt Patterns but Duration of Phenological Periods Is Fixed: An Example of Periodicity. Environ. Res. Lett. 2016, 11, 125006. [Google Scholar] [CrossRef]
- Testolin, R.; Carmona, C.P.; Attorre, F.; Borchardt, P.; Bruelheide, H.; Dolezal, J.; Finckh, M.; Haider, S.; Hemp, A.; Jandt, U.; et al. Global Functional Variation in Alpine Vegetation. J. Veg. Sci. 2021, 32, e13000. [Google Scholar] [CrossRef]
- Lind, P.; Belušić, D.; Médus, E.; Dobler, A.; Pedersen, R.A.; Wang, F.; Matte, D.; Kjellström, E.; Landgren, O.; Lindstedt, D.; et al. Climate Change Information over Fenno-Scandinavia Produced with a Convection-Permitting Climate Model. Clim. Dyn. 2023, 61, 519–541. [Google Scholar] [CrossRef]
- Forbes, B.C.; Kumpula, T. The Ecological Role and Geography of Reindeer (Rangifer Tarandus) in Northern Eurasia: Ecology/Geography of Eurasian Reindeer. Geogr. Compass 2009, 3, 1356–1380. [Google Scholar] [CrossRef]
- Stark, S.; Horstkotte, T.; Kumpula, J.; Olofsson, J.; Tømmervik, H.; Turunen, M. The Ecosystem Effects of Reindeer (Rangifer Tarandus) in Northern Fennoscandia: Past, Present and Future. Perspect. Plant Ecol. Evol. Syst. 2023, 58, 125716. [Google Scholar] [CrossRef]
- Austrheim, G.; Eriksson, O. Plant Species Diversity and Grazing in the Scandinavian Mountains - Patterns and Processes at Different Spatial Scales. Ecography 2008, 24, 683–695. [Google Scholar] [CrossRef]
- Egelkraut, D.; Aronsson, K.-Å.; Allard, A.; Åkerholm, M.; Stark, S.; Olofsson, J. Multiple Feedbacks Contribute to a Centennial Legacy of Reindeer on Tundra Vegetation. Ecosystems 2018, 21, 1545–1563. [Google Scholar] [CrossRef]
- Cohen, J.; Pulliainen, J.; Ménard, C.B.; Johansen, B.; Oksanen, L.; Luojus, K.; Ikonen, J. Effect of Reindeer Grazing on Snowmelt, Albedo and Energy Balance Based on Satellite Data Analyses. Remote Sens. Environ. 2013, 135, 107–117. [Google Scholar] [CrossRef]
- Skogland, T. Comparative Summer Feeding Strategies of Arctic and Alpine Rangifer. J. Anim. Ecol. 1980, 49, 81. [Google Scholar] [CrossRef]
- Mårell, A.; Hofgaard, A.; Danell, K. Nutrient Dynamics of Reindeer Forage Species along Snowmelt Gradients at Different Ecological Scales. Basic. Appl. Ecol. 2006, 7, 13–30. [Google Scholar] [CrossRef]
- Borner, A.P.; Kielland, K.; Walker, M.D. Effects of Simulated Climate Change on Plant Phenology and Nitrogen Mineralization in Alaskan Arctic Tundra. Arct. Antarct. Alp. Res. 2008, 40, 27–38. [Google Scholar] [CrossRef]
- Klein, D.R. Variation in Quality of Caribou and Reindeer Forage Plants Associated with Season, Plant Part, and Phenology. Ran 1990, 10, 123. [Google Scholar] [CrossRef]
- Åhman, B.; White, R.G. Rangifer Diet and Nutritional Needs. In Reindeer and Caribou; Tryland, M., Kutz, S.J., Eds.; CRC Press: Boca Raton, FL, USA; Taylor & Francis: Abingdon, UK, 2018; pp. 107–134. [Google Scholar] [CrossRef]
- Skarin, A.; Danell, Ö.; Bergström, R.; Moen, J. Reindeer Movement Patterns in Alpine Summer Ranges. Polar Biol. 2010, 33, 1263–1275. [Google Scholar] [CrossRef]
- Niittynen, P.; Heikkinen, R.K.; Luoto, M. Decreasing Snow Cover Alters Functional Composition and Diversity of Arctic Tundra. Proc. Natl. Acad. Sci. USA 2020, 117, 21480–21487. [Google Scholar] [CrossRef]
- Mioduszewski, J.R.; Rennermalm, A.K.; Robinson, D.A.; Wang, L. Controls on Spatial and Temporal Variability in Northern Hemisphere Terrestrial Snow Melt Timing, 1979–2012. J. Clim. 2015, 28, 2136–2153. [Google Scholar] [CrossRef]
- Bourbigot, M.; Hajduch, G.; Johnsen, H.; Piantanida, R. Sentinel-1 Product Specification, 2023. European Space Agency, Frascati, Italy. Available online: https://sentinels.copernicus.eu/documents/247904/0/S1-RS-MDA-52-7441_3_14_1_Sentinel-1ProductSpecification/c107bebf-37dc-7db4-4582-fd79b15f3ea2 (accessed on 19 September 2022).
- Nagler, T.; Rott, H.; Ripper, E.; Bippus, G.; Hetzenecker, M. Advancements for Snowmelt Monitoring by Means of Sentinel-1 SAR. Remote Sens. 2016, 8, 348. [Google Scholar] [CrossRef]
- Buchelt, S.; Skov, K.; Rasmussen, K.K.; Ullmann, T. Sentinel-1 Time Series for Mapping Snow Cover Depletion and Timing of Snowmelt in Arctic Periglacial Environments: Case Study from Zackenberg and Kobbefjord, Greenland. Cryosphere 2022, 16, 625–646. [Google Scholar] [CrossRef]
- Vickers, H.; Malnes, E.; Eckerstorfer, M. A Synthetic Aperture Radar Based Method for Long Term Monitoring of Seasonal Snowmelt and Wintertime Rain-On-Snow Events in Svalbard. Front. Earth Sci. 2022, 10, 868945. [Google Scholar] [CrossRef]
- Awasthi, S.; Varade, D.; Kumar Thakur, P.; Kumar, A.; Singh, H.; Jain, K.; Snehmani. Development of a Novel Approach for Snow Wetness Estimation Using Hybrid Polarimetric RISAT-1 SAR Datasets in North-Western Himalayan Region. J. Hydrol. 2022, 612, 128252. [Google Scholar] [CrossRef]
- Dean, A.M.; Brown, I.A.; Huntley, B.; Thomas, C.J. Monitoring Snowmelt across the Arctic Forest–Tundra Ecotone Using Synthetic Aperture Radar. Int. J. Remote Sens. 2006, 27, 4347–4370. [Google Scholar] [CrossRef]
- Swedish Infrastructure for Ecosystem Science. Available online: https://data.fieldsites.se/portal/ (accessed on 28 April 2023).
- Fohringer, C.; Rosqvist, G.; Inga, N.; Singh, N.J. Reindeer Husbandry in Peril?—How Extractive Industries Exert Multiple Pressures on an Arctic Pastoral Ecosystem. People Nat. 2021, 3, 872–886. [Google Scholar] [CrossRef]
- Naturvårdsverket. Nationella marktäckedata 2018 tilläggsskikt Låg fjällskog. Available online: https://www.geodata.se/geodataportalen (accessed on 20 October 2022).
- Lantmäteriet. Produktbeskrivning: GSD-Vegetationsdata. Available online: https://www.geodata.se/geodataportalen (accessed on 9 October 2022).
- Lantmäteriet. Product Description: GSD-Elevation Data, Grid 2+. Available online: https://www.geodata.se/geodataportalen (accessed on 10 September 2022).
- Alaska Satellite Facility. ASF Data Search Vertex. Available online: https://search.asf.alaska.edu/#/ (accessed on 12 September 2022).
- Small, D.; Zuberbühler, L.; Schubert, A.; Meier, E. Terrain-Flattened Gamma Nought Radarsat-2 Backscatter. Can. J. Remote Sens. 2011, 37, 493–499. [Google Scholar] [CrossRef]
- Small, D.; Rohner, C.; Miranda, N.; Ruetschi, M.; Schaepman, M.E. Wide-Area Analysis-Ready Radar Backscatter Composites. IEEE Trans. Geosci. Remote Sens. 2022, 60, 1–14. [Google Scholar] [CrossRef]
- Lee, J.-S.; Wen, J.-H.; Ainsworth, T.L.; Chen, K.-S.; Chen, A.J. Improved Sigma Filter for Speckle Filtering of SAR Imagery. IEEE Trans. Geosci. Remote Sens. 2009, 47, 202–213. [Google Scholar] [CrossRef]
- Marin, C.; Bertoldi, G.; Premier, V.; Callegari, M.; Brida, C.; Hürkamp, K.; Tschiersch, J.; Zebisch, M.; Notarnicola, C. Use of Sentinel-1 Radar Observations to Evaluate Snowmelt Dynamics in Alpine Regions. Cryosphere 2020, 14, 935–956. [Google Scholar] [CrossRef]
- Schmidt, K.; Schwerdt, M.; Miranda, N.; Reimann, J. Radiometric Comparison within the Sentinel-1 SAR Constellation over a Wide Backscatter Range. Remote Sens. 2020, 12, 854. [Google Scholar] [CrossRef]
- Lund, J.; Forster, R.R.; Deeb, E.J.; Liston, G.E.; Skiles, S.M.; Marshall, H.-P. Interpreting Sentinel-1 SAR Backscatter Signals of Snowpack Surface Melt/Freeze, Warming, and Ripening, through Field Measurements and Physically-Based SnowModel. Remote Sens. 2022, 14, 4002. [Google Scholar] [CrossRef]
- Swedish Meteorological and Hydrological Institute. Snödjup (In Swedish). Available online: https://www.smhi.se/vader/observationer/snodjup/ (accessed on 20 February 2024).
- Moriana-Armendariz, M.; Nilsen, L.; Cooper, E.J. Natural Variation in Snow Depth and Snow Melt Timing in the High Arctic Have Implications for Soil and Plant Nutrient Status and Vegetation Composition. Arct. Sci. 2022, 8, 767–785. [Google Scholar] [CrossRef]
- Swedish Meteorological and Hydrological Institute. Sommaren 2018—Extremt Varm Och Solig (In Swedish). Available online: https://www.smhi.se/klimat/klimatet-da-och-nu/arets-vader/sommaren-2018-extremt-varm-och-solig-1.138134 (accessed on 25 May 2024).
- Kudo, G.; Nordenhäll, U.; Molau, U. Effects of Snowmelt Timing on Leaf Traits, Leaf Production, and Shoot Growth of Alpine Plants: Comparisons along a Snowmelt Gradient in Northern Sweden. Écoscience 1999, 6, 439–450. [Google Scholar] [CrossRef]
- Foster, J.L.; Robinson, D.A.; Hall, D.K.; Estilow, T.W. Spring Snow Melt Timing and Changes over Arctic Lands. Polar Geogr. 2008, 31, 145–157. [Google Scholar] [CrossRef]
- Callaghan, T.V.; Johansson, M.; Brown, R.D.; Groisman, P.Y.; Labba, N.; Radionov, V.; Barry, R.G.; Bulygina, O.N.; Essery, R.L.H.; Frolov, D.M.; et al. The Changing Face of Arctic Snow Cover: A Synthesis of Observed and Projected Changes. AMBIO 2011, 40, 17–31. [Google Scholar] [CrossRef]
- Schimanke, S.; Joelsson, M.; Andersson, S.; Carlund, T.; Wern, L.; Hellström, S.; Kjellström, E. Observerad Klimatförändring i Sverige 1860–2021; SMHI: Norrköping, Sweden, 2022; p. 89. ISSN 1654-2258.
- Kivinen, S.; Kaarlejärvi, E.; Jylhä, K.; Räisänen, J. Spatiotemporal Distribution of Threatened High-Latitude Snowbed and Snow Patch Habitats in Warming Climate. Environ. Res. Lett. 2012, 7, 034024. [Google Scholar] [CrossRef]
- Vowles, T.; Gunnarsson, B.; Molau, U.; Hickler, T.; Klemedtsson, L.; Björk, R.G. Expansion of Deciduous Tall Shrubs but Not Evergreen Dwarf Shrubs Inhibited by Reindeer in Scandes Mountain Range. J. Ecol. 2017, 105, 1547–1561. [Google Scholar] [CrossRef]
- Chagnon, C.; Boudreau, S. Shrub Canopy Induces a Decline in Lichen Abundance and Diversity in Nunavik (Québec, Canada). Arct. Antarct. Alp. Res. 2019, 51, 521–532. [Google Scholar] [CrossRef]
- Bjorkman, A.D.; Myers-Smith, I.H.; Elmendorf, S.C.; Normand, S.; Rüger, N.; Beck, P.S.A.; Blach-Overgaard, A.; Blok, D.; Cornelissen, J.H.C.; Forbes, B.C.; et al. Plant Functional Trait Change across a Warming Tundra Biome. Nature 2018, 562, 57–62. [Google Scholar] [CrossRef]
- Xie, J.; Jonas, T.; Rixen, C.; de Jong, R.; Garonna, I.; Notarnicola, C.; Asam, S.; Schaepman, M.E.; Kneubühler, M. Land Surface Phenology and Greenness in Alpine Grasslands Driven by Seasonal Snow and Meteorological Factors. Sci. Total Environ. 2020, 725, 138380. [Google Scholar] [CrossRef]
- Zong, S.; Lembrechts, J.J.; Du, H.; He, H.S.; Wu, Z.; Li, M.; Rixen, C. Upward Range Shift of a Dominant Alpine Shrub Related to 50 Years of Snow Cover Change. Remote Sens. Environ. 2022, 268, 112773. [Google Scholar] [CrossRef]
- Cooper, E.J.; Dullinger, S.; Semenchuk, P. Late Snowmelt Delays Plant Development and Results in Lower Reproductive Success in the High Arctic. Plant Sci. 2011, 180, 157–167. [Google Scholar] [CrossRef] [PubMed]
- Dunne, J.A.; Harte, J.; Taylor, K.J. Subalpine Meadow Flowering Phenology Responses to Climate Change: Integrating Experimental and Gradient Methods. Ecol. Monogr. 2003, 73, 69–86. [Google Scholar] [CrossRef]
- Frei, E.R.; Henry, G.H.R. Long-Term Effects of Snowmelt Timing and Climate Warming on Phenology, Growth, and Reproductive Effort of Arctic Tundra Plant Species. Arct. Sci. 2022, 8, 700–721. [Google Scholar] [CrossRef]
- Galen, C.; Stanton, M.L. Short-Term Responses of Alpine Buttercups to Experimental Manipulations of Growing Season Length. Ecology 1993, 74, 1052–1058. [Google Scholar] [CrossRef]
- Wipf, S. Phenology, Growth, and Fecundity of Eight Subarctic Tundra Species in Response to Snowmelt Manipulations. Plant Ecol. 2010, 207, 53–66. [Google Scholar] [CrossRef]
- Hallinger, M.; Manthey, M.; Wilmking, M. Establishing a Missing Link: Warm Summers and Winter Snow Cover Promote Shrub Expansion into Alpine Tundra in Scandinavia. New Phytol. 2010, 186, 890–899. [Google Scholar] [CrossRef]
- Rumpf, S.B.; Gravey, M.; Brönnimann, O.; Luoto, M.; Cianfrani, C.; Mariethoz, G.; Guisan, A. From White to Green: Snow Cover Loss and Increased Vegetation Productivity in the European Alps. Science 2022, 376, 1119–1122. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SOSM | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2017 | 2018 | 2019 | 2020 | 2021 | ||||||||||
| Date | VV | VH | Date | VV | VH | Date | VV | VH | Date | VV | VH | Date | VV | VH |
| 2-Mar | 0% | 4% | 3-Mar | 1% | 1% | 4-Mar | 3% | 5% | 4-Mar | 1% | 1% | 5-Mar | 0% | 2% |
| 8-Mar | 0% | 2% | 9-Mar | 1% | 4% | 10-Mar | 2% | 2% | 10-Mar | 1% | 2% | 11-Mar | 0% | 2% |
| 14-Mar | 0% | 2% | 15-Mar | 1% | 1% | 16-Mar | 3% | 5% | 16-Mar | 0% | 0% | 17-Mar | 0% | 3% |
| 20-Mar | 56% | 1% | 21-Mar | 1% | 2% | 22-Mar | 1% | 1% | 22-Mar | 1% | 2% | 23-Mar | 0% | 1% |
| 26-Mar | 0% | 2% | 27-Mar | 1% | 1% | 28-Mar | 4% | 3% | 28-Mar | 0% | 0% | 29-Mar | 0% | 1% |
| 1-Apr | 0% | 1% | 2-Apr | 1% | 2% | 3-Apr | 1% | 1% | 3-Apr | 1% | 2% | 4-Apr | 0% | 1% |
| 7-Apr | 0% | 1% | 8-Apr | 1% | 0% | 9-Apr | 2% | 3% | 9-Apr | 1% | 0% | 10-Apr | 0% | 1% |
| 13-Apr | 0% | 1% | 14-Apr | 4% | 5% | 15-Apr | 3% | 2% | 15-Apr | 1% | 1% | 16-Apr | 2% | 6% |
| 19-Apr | 0% | 1% | 20-Apr | 21% | 11% | 21-Apr | 19% | 25% | 21-Apr | 4% | 5% | 22-Apr | 0% | 4% |
| 25-Apr | 0% | 2% | 26-Apr | 19% | 21% | 27-Apr | 14% | 5% | 27-Apr | 1% | 3% | 28-Apr | 0% | 0% |
| 1-May | 0% | 2% | 2-May | 6% | 2% | 3-May | 4% | 6% | 3-May | 5% | 3% | 4-May | 0% | 1% |
| 7-May | 0% | 4% | 8-May | 6% | 14% | 9-May | 5% | 3% | 9-May | 2% | 3% | 10-May | 1% | 3% |
| 13-May | 0% | 7% | 14-May | 21% | 10% | 15-May | 9% | 9% | 15-May | 0% | 0% | 16-May | 56% | 78% |
| 19-May | 18% | 19% | 20-May | 3% | 11% | 21-May | 10% | 5% | 21-May | 19% | 21% | 22-May | 2% | 3% |
| 25-May | 6% | 14% | 26-May | 1% | 0% | 27-May | 10% | 12% | 27-May | 38% | 23% | 28-May | 3% | 11% |
| 31-May | 1% | 2% | 1-Jun | 1% | 1% | 2-Jun | 3% | 3% | 2-Jun | 10% | 21% | 3-Jun | 3% | 1% |
| 6-Jun | 10% | 22% | 7-Jun | 1% | 1% | 8-Jun | 1% | 2% | 8-Jun | 6% | 3% | 9-Jun | 1% | 1% |
| 12-Jun | 1% | 1% | 13-Jun | 2% | 4% | 14-Jun | 0% | 0% | 14-Jun | 1% | 3% | 15-Jun | 0% | 0% |
| 18-Jun | 2% | 3% | 19-Jun | 0% | 0% | 20-Jun | 1% | 1% | 20-Jun | 2% | 0% | 21-Jun | 0% | 1% |
| 24-Jun | 1% | 0% | 25-Jun | 3% | 6% | 26-Jun | 0% | 0% | 26-Jun | 0% | 1% | 27-Jun | 0% | 0% |
| 30-Jun | 2% | 1% | 1-Jul | 1% | 0% | 2-Jul | 2% | 3% | 2-Jul | 1% | 1% | 4-Jul | 0% | 0% |
| 6-Jul | 0% | 0% | 7-Jul | 1% | 1% | 7-Jul | 0% | 0% | 8-Jul | 1% | 1% | 9-Jul | 0% | 0% |
| 12-Jul | 0% | 1% | 13-Jul | 0% | 0% | 14-Jul | 1% | 0% | 14-Jul | 0% | 0% | 15-Jul | 53% | 0% |
| 18-Jul | 0% | 1% | 19-Jul | 0% | 0% | 20-Jul | 0% | 0% | 20-Jul | 0% | 1% | 21-Jul | 0% | 1% |
| 24-Jul | 1% | 0% | 25-Jul | 0% | 0% | 26-Jul | 0% | 0% | 26-Jul | 0% | 0% | 27-Jul | 0% | 0% |
| Year | Date | AWS Snow Free Date | Polarisation |
|---|---|---|---|
| 2017 | 12 June | 15 June | VV |
| 12 June and 18 June | VH | ||
| 2018 | 20 May | 22 May | VV |
| 26 May and 1 June | VH | ||
| 2019 | 7 August | 22 May | VV |
| 2 May and 13 August | VH | ||
| 2020 | 9 June | 19 June | VV |
| 21 June | VH |
| SOSM | |||||||||||||
| 2017 | 2018 | ||||||||||||
| VV | VH | VV | VH | ||||||||||
| Vegetation | Area ** | Mean DOY | STD * | Mean DOY | STD * | Mean DOY | STD * | Mean DOY | STD * | ||||
| Dry heath (e.g., B. nana) | 131.3 | 98 | ± | 30 | 125 | ± | 34 | 115 | ± | 19 | 114 | ± | 20 |
| Alpine low grass meadow (e.g., C. Bigelowii) | 132.0 | 103 | ± | 31 | 134 | ± | 29 | 117 | ± | 18 | 118 | ± | 19 |
| Low-growing shrubs | 37.2 | 101 | ± | 31 | 128 | ± | 29 | 116 | ± | 13 | 115 | ± | 14 |
| Grassland (e.g., D. flexuosa) | 104.7 | 110 | ± | 35 | 144 | ± | 28 | 124 | ± | 20 | 128 | ± | 23 |
| Open marsh vegetation | 14.2 | 90 | ± | 29 | 118 | ± | 36 | 118 | ± | 19 | 113 | ± | 21 |
| Heath (e.g., heather) | 80.0 | 93 | ± | 28 | 122 | ± | 33 | 113 | ± | 19 | 111 | ± | 20 |
| Extreme dry heath (e.g., B. nana) | 21.6 | 93 | ± | 26 | 123 | ± | 36 | 116 | ± | 17 | 118 | ± | 17 |
| Alpine tall grass meadow | 9.2 | 101 | ± | 31 | 132 | ± | 27 | 114 | ± | 20 | 108 | ± | 19 |
| Moist–wet heath (e.g., Sweetgale) | 3.9 | 121 | ± | 31 | 139 | ± | 19 | 117 | ± | 13 | 116 | ± | 15 |
| 2019 | 2020 | ||||||||||||
| VV | VH | VV | VH | ||||||||||
| Vegetation | Area ** | Mean DOY | STD * | Mean DOY | STD * | Mean DOY | STD * | Mean DOY | STD * | ||||
| Dry heath (e.g., B. nana) | 131.3 | 112 | ± | 26 | 108 | ± | 25 | 140 | ± | 21 | 136 | ± | 25 |
| Alpine low grass meadow (e.g., C. Bigelowii) | 132.0 | 119 | ± | 23 | 115 | ± | 24 | 144 | ± | 17 | 140 | ± | 23 |
| Low-growing shrubs | 37.2 | 113 | ± | 20 | 109 | ± | 19 | 142 | ± | 15 | 139 | ± | 18 |
| Grassland (e.g., D. flexuosa) | 104.7 | 125 | ± | 26 | 128 | ± | 27 | 145 | ± | 19 | 146 | ± | 23 |
| Open marsh vegetation | 14.2 | 110 | ± | 35 | 99 | ± | 31 | 136 | ± | 25 | 129 | ± | 24 |
| Heath (e.g., heather) | 80.0 | 110 | ± | 27 | 105 | ± | 24 | 137 | ± | 23 | 132 | ± | 26 |
| Extreme dry heath (e.g., B. nana) | 21.6 | 110 | ± | 27 | 107 | ± | 26 | 137 | ± | 23 | 132 | ± | 28 |
| Alpine tall grass meadow | 9.2 | 119 | ± | 26 | 111 | ± | 24 | 140 | ± | 22 | 137 | ± | 19 |
| Moist–wet heath (e.g., Sweetgale) | 3.9 | 122 | ± | 18 | 117 | ± | 18 | 147 | ± | 8 | 145 | ± | 13 |
| 2021 | |||||||||||||
| VV | VH | ||||||||||||
| Vegetation | Area ** | Mean DOY | STD * | Mean DOY | STD * | ||||||||
| Dry heath (e.g., B. nana) | 131.3 | 161 | ± | 30 | 128 | ± | 21 | ||||||
| Alpine low grass meadow (e.g., C. Bigelowii) | 132.0 | 158 | ± | 29 | 134 | ± | 16 | ||||||
| Low-growing shrubs | 37.2 | 151 | ± | 27 | 131 | ± | 16 | ||||||
| Grassland (e.g., D. flexuosa) | 104.7 | 157 | ± | 27 | 137 | ± | 13 | ||||||
| Open marsh vegetation | 14.2 | 157 | ± | 37 | 116 | ± | 29 | ||||||
| Heath (e.g., heather) | 80.0 | 163 | ± | 31 | 126 | ± | 22 | ||||||
| Extreme dry heath (e.g., B. nana) | 21.6 | 166 | ± | 30 | 128 | ± | 21 | ||||||
| Alpine tall grass meadow | 9.2 | 170 | ± | 31 | 124 | ± | 21 | ||||||
| Moist–wet heath (e.g., Sweetgale) | 3.9 | 144 | ± | 21 | 134 | ± | 15 | ||||||
| EOS | |||||||||||||
| 2017 | 2018 | ||||||||||||
| VV | VH | VV | VH | ||||||||||
| Vegetation | Area ** | Mean DOY | STD * | Mean DOY | STD * | Mean DOY | STD * | Mean DOY | STD * | ||||
| Dry heath (e.g., B. nana) | 131.3 | 117 | ± | 40 | 126 | ± | 34 | 125 | ± | 60 | 127 | ± | 60 |
| Alpine low grass meadow (e.g., C. Bigelowii) | 132.0 | 126 | ± | 38 | 130 | ± | 29 | 127 | ± | 56 | 130 | ± | 63 |
| Low-growing shrubs | 37.2 | 121 | ± | 39 | 137 | ± | 29 | 128 | ± | 41 | 129 | ± | 38 |
| Grassland (e.g., D. flexuosa) | 104.7 | 133 | ± | 42 | 114 | ± | 28 | 124 | ± | 72 | 119 | ± | 91 |
| Open marsh vegetation | 14.2 | 97 | ± | 49 | 108 | ± | 36 | 121 | ± | 55 | 120 | ± | 62 |
| Heath (e.g., heather) | 80.0 | 112 | ± | 39 | 128 | ± | 33 | 123 | ± | 59 | 126 | ± | 53 |
| Extreme dry heath (e.g., B. nana) | 21.6 | 110 | ± | 37 | 104 | ± | 36 | 118 | ± | 71 | 121 | ± | 75 |
| Alpine tall grass meadow | 9.2 | 120 | ± | 39 | 123 | ± | 27 | 120 | ± | 67 | 122 | ± | 62 |
| Moist−wet heath (e.g., Sweetgale) | 3.9 | 142 | ± | 36 | 150 | ± | 19 | 131 | ± | 30 | 129 | ± | 34 |
| 2019 | 2020 | ||||||||||||
| VV | VH | VV | VH | ||||||||||
| Vegetation | Area ** | Mean DOY | STD * | Mean DOY | STD * | Mean DOY | STD * | Mean DOY | STD * | ||||
| Dry heath (e.g., B. nana) | 131.3 | 125 | ± | 79 | 133 | ± | 58 | 144 | ± | 61 | 142 | ± | 70 |
| Alpine low grass meadow (e.g., C. Bigelowii) | 132.0 | 130 | ± | 77 | 138 | ± | 63 | 150 | ± | 51 | 145 | ± | 71 |
| Low-growing shrubs | 37.2 | 123 | ± | 63 | 126 | ± | 45 | 147 | ± | 48 | 148 | ± | 48 |
| Grassland (e.g., D. flexuosa) | 104.7 | 125 | ± | 90 | 123 | ± | 97 | 145 | ± | 68 | 133 | ± | 93 |
| Open marsh vegetation | 14.2 | 109 | ± | 87 | 115 | ± | 70 | 134 | ± | 70 | 131 | ± | 77 |
| Heath (e.g., heather) | 80.0 | 120 | ± | 82 | 130 | ± | 52 | 142 | ± | 62 | 144 | ± | 59 |
| Extreme dry heath (e.g., B. nana) | 21.6 | 118 | ± | 91 | 129 | ± | 71 | 137 | ± | 73 | 130 | ± | 88 |
| Alpine tall grass meadow | 9.2 | 121 | ± | 86 | 130 | ± | 68 | 137 | ± | 71 | 145 | ± | 58 |
| Moist−wet heath (e.g., Sweetgale) | 3.9 | 132 | ± | 42 | 131 | ± | 42 | 155 | ± | 26 | 152 | ± | 41 |
| 2021 | |||||||||||||
| VV | VH | ||||||||||||
| Vegetation | Area ** | Mean DOY | STD * | Mean DOY | STD * | ||||||||
| Dry heath (e.g., B. nana) | 131.3 | 182 | ± | 27 | 151 | ± | 13 | ||||||
| Alpine low grass meadow (e.g., C. Bigelowii) | 132.0 | 182 | ± | 27 | 154 | ± | 13 | ||||||
| Low-growing shrubs | 37.2 | 170 | ± | 29 | 147 | ± | 10 | ||||||
| Grassland (e.g., D. flexuosa) | 104.7 | 180 | ± | 26 | 156 | ± | 13 | ||||||
| Open marsh vegetation | 14.2 | 184 | ± | 29 | 143 | ± | 18 | ||||||
| Heath (e.g., heather) | 80.0 | 184 | ± | 27 | 148 | ± | 13 | ||||||
| Extreme dry heath (e.g., B. nana) | 21.6 | 186 | ± | 26 | 152 | ± | 14 | ||||||
| Alpine tall grass meadow | 9.2 | 191 | ± | 23 | 150 | ± | 16 | ||||||
| Moist−wet heath (e.g., Sweetgale) | 3.9 | 163 | ± | 26 | 148 | ± | 8 | ||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carlsson, I.; Rosqvist, G.; Wennbom, J.M.; Brown, I.A. Synthetic Aperture Radar Monitoring of Snow in a Reindeer-Grazing Landscape. Remote Sens. 2024, 16, 2329. https://doi.org/10.3390/rs16132329
Carlsson I, Rosqvist G, Wennbom JM, Brown IA. Synthetic Aperture Radar Monitoring of Snow in a Reindeer-Grazing Landscape. Remote Sensing. 2024; 16(13):2329. https://doi.org/10.3390/rs16132329
Chicago/Turabian StyleCarlsson, Ida, Gunhild Rosqvist, Jenny Marika Wennbom, and Ian A. Brown. 2024. "Synthetic Aperture Radar Monitoring of Snow in a Reindeer-Grazing Landscape" Remote Sensing 16, no. 13: 2329. https://doi.org/10.3390/rs16132329