
The Variations of Land Surface Phenology in Northeast China and Its Responses to Climate Change from 1982 to 2013

 , ,
, ,  ,
,
Abstract
:
1. Introduction
2. Materials and Methods
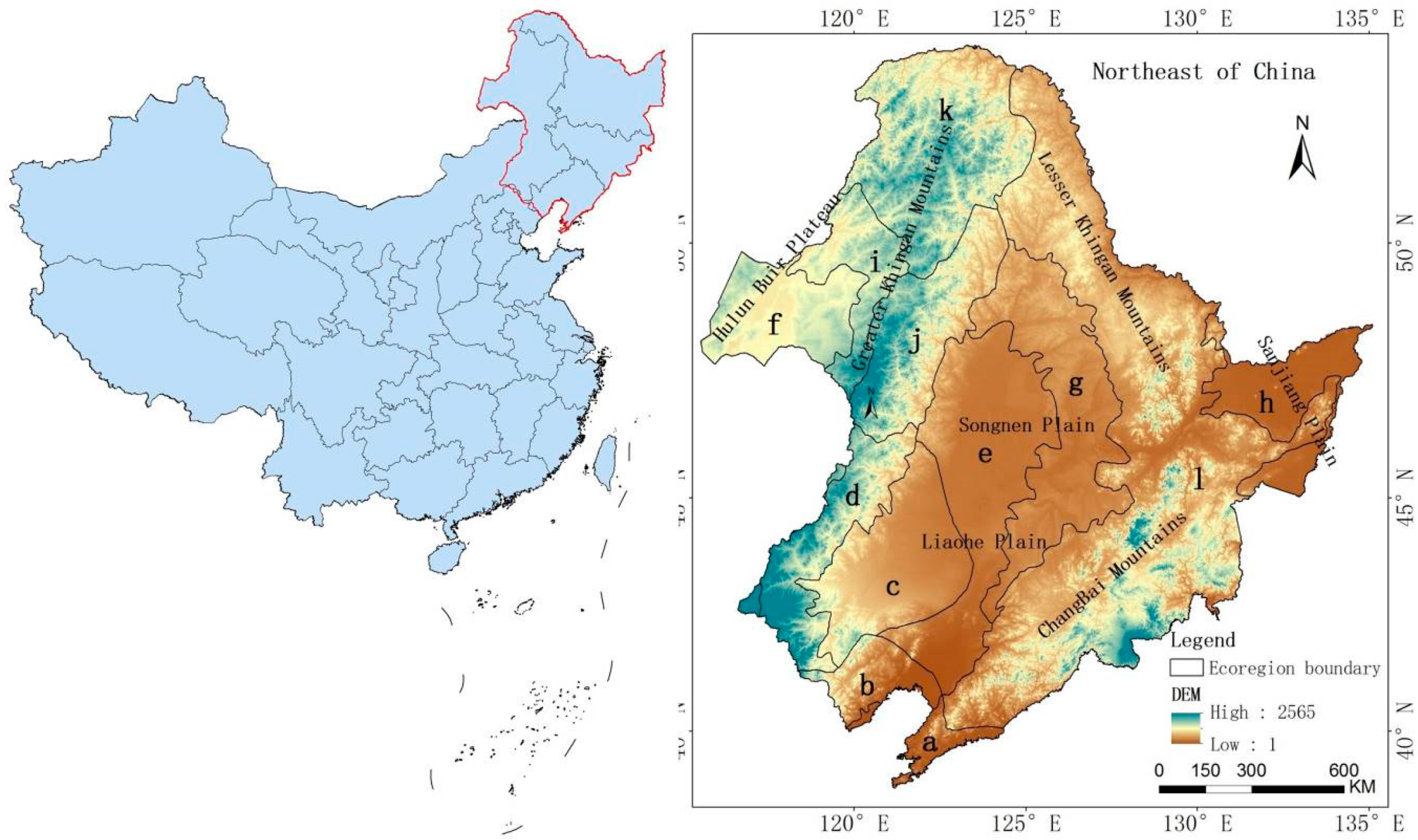
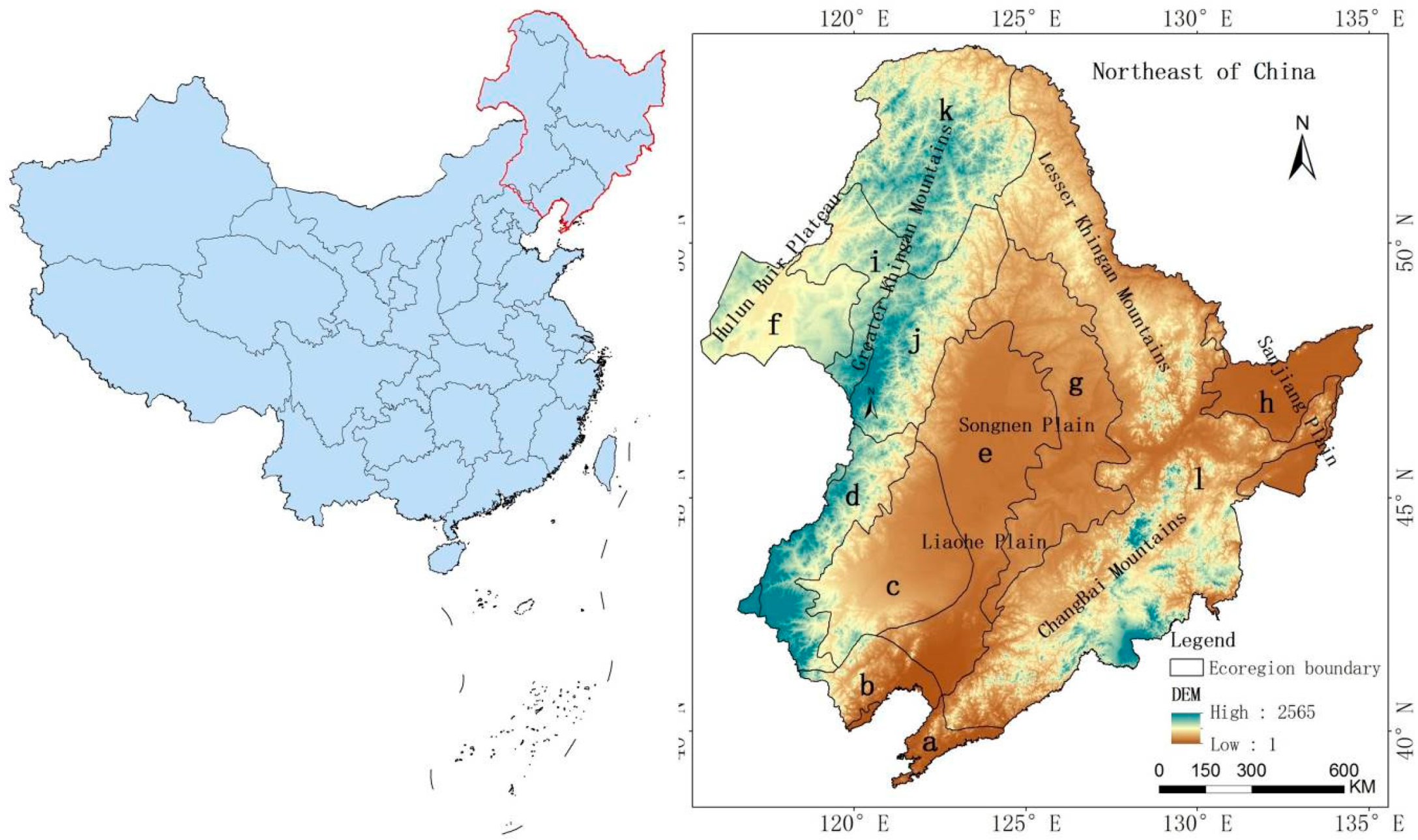
2.1. Study Area
2.2. GIMMS NDVI3g Dataset
2.3. Climate Data
2.4. Phenology Metrics
3. Results
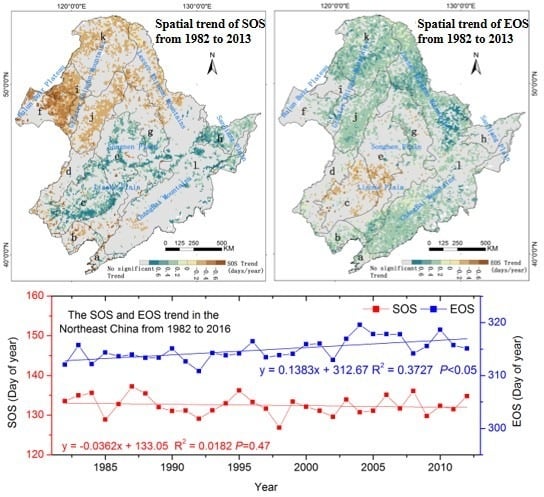
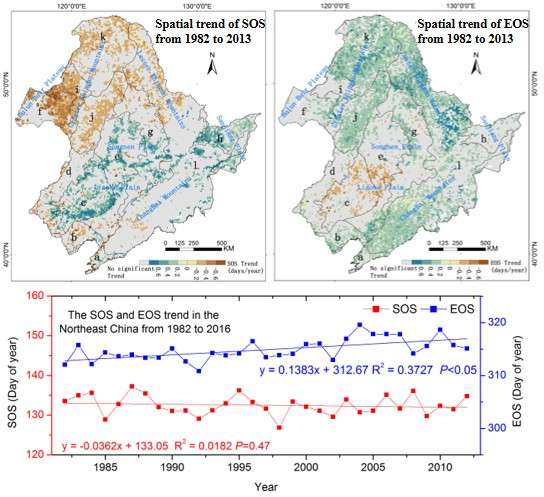
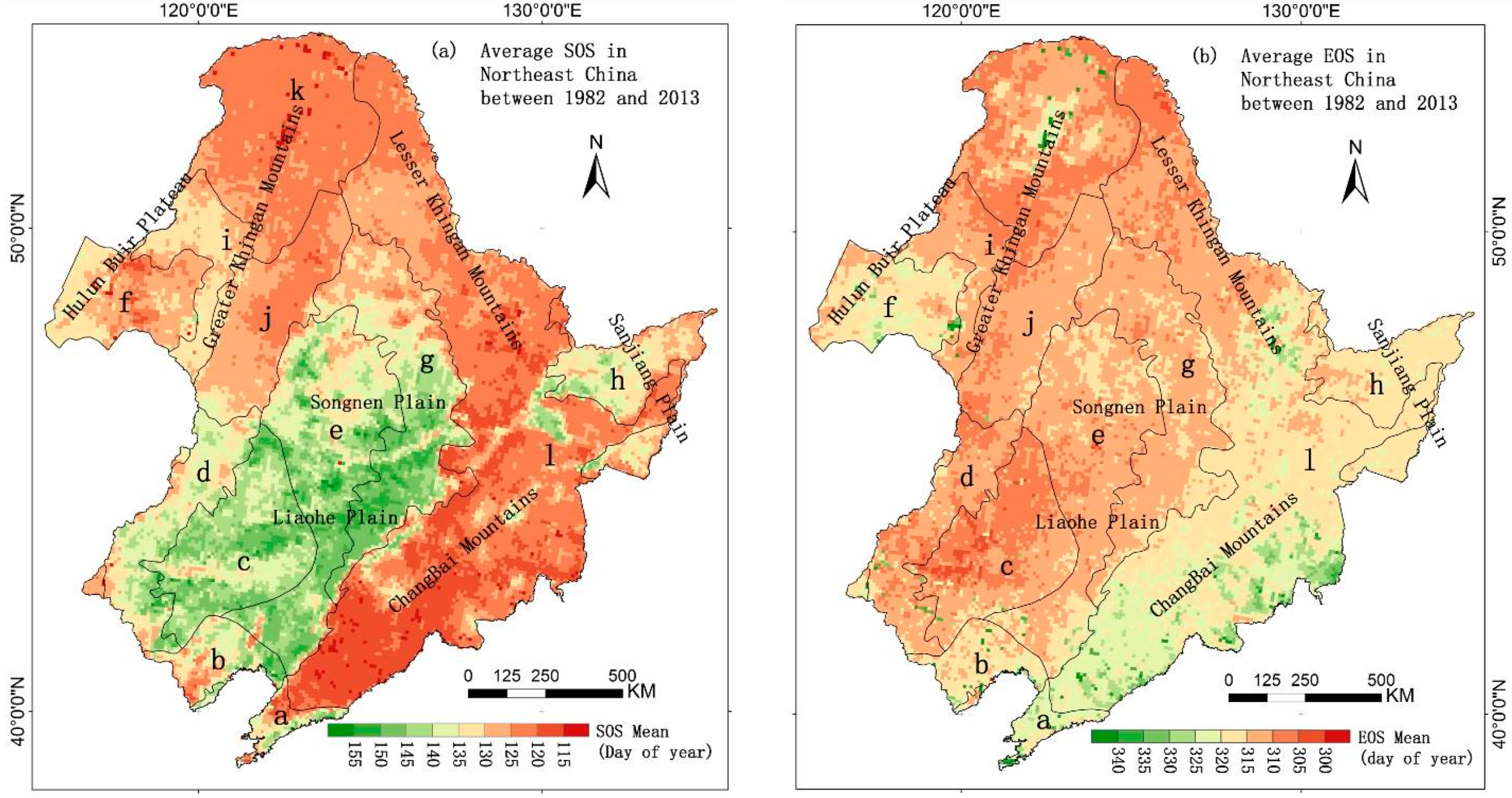
3.1. The Mean Spatial Distribution of Land Surface Phenology
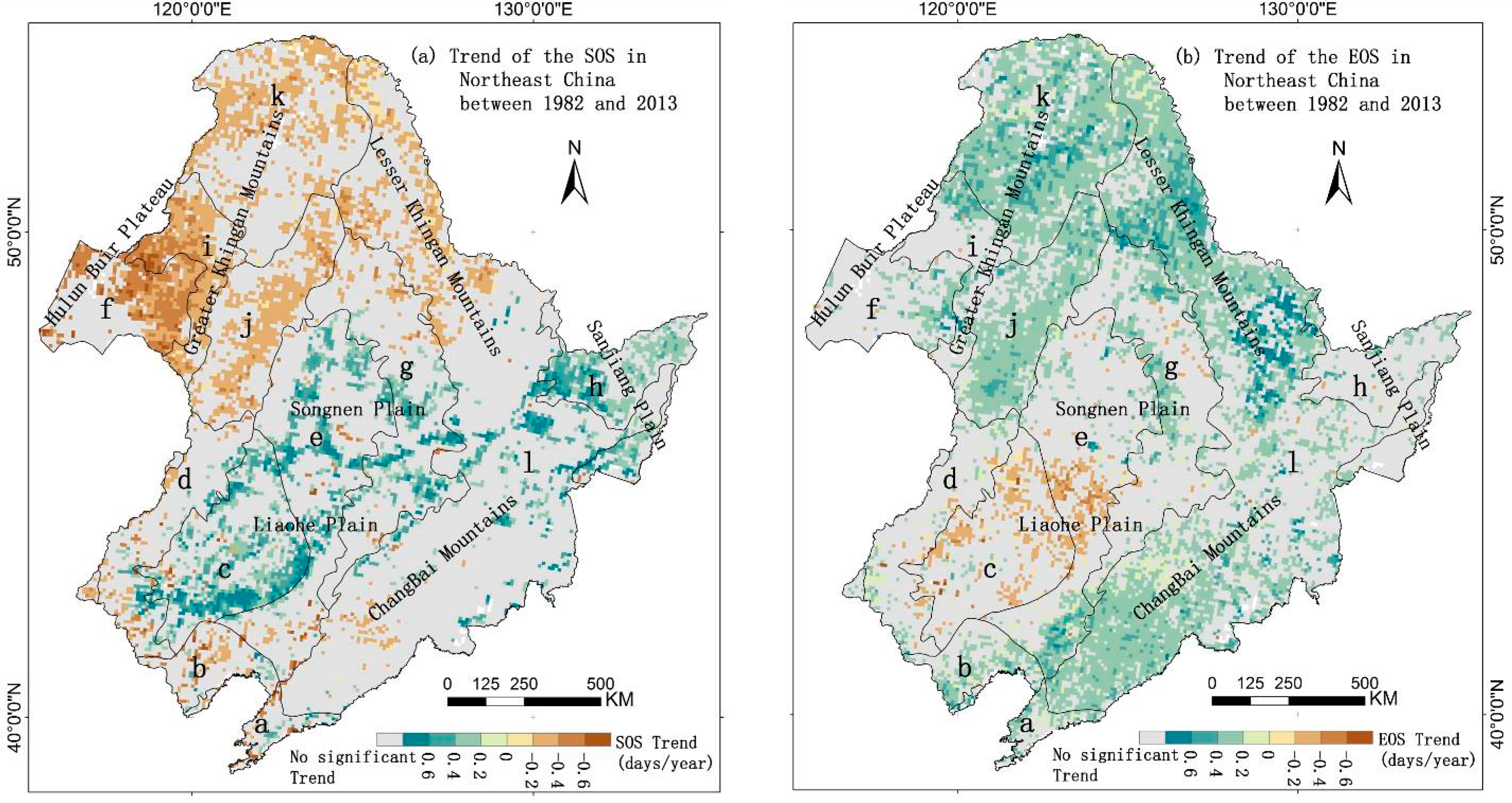
3.2. Spatial Phenology Trends
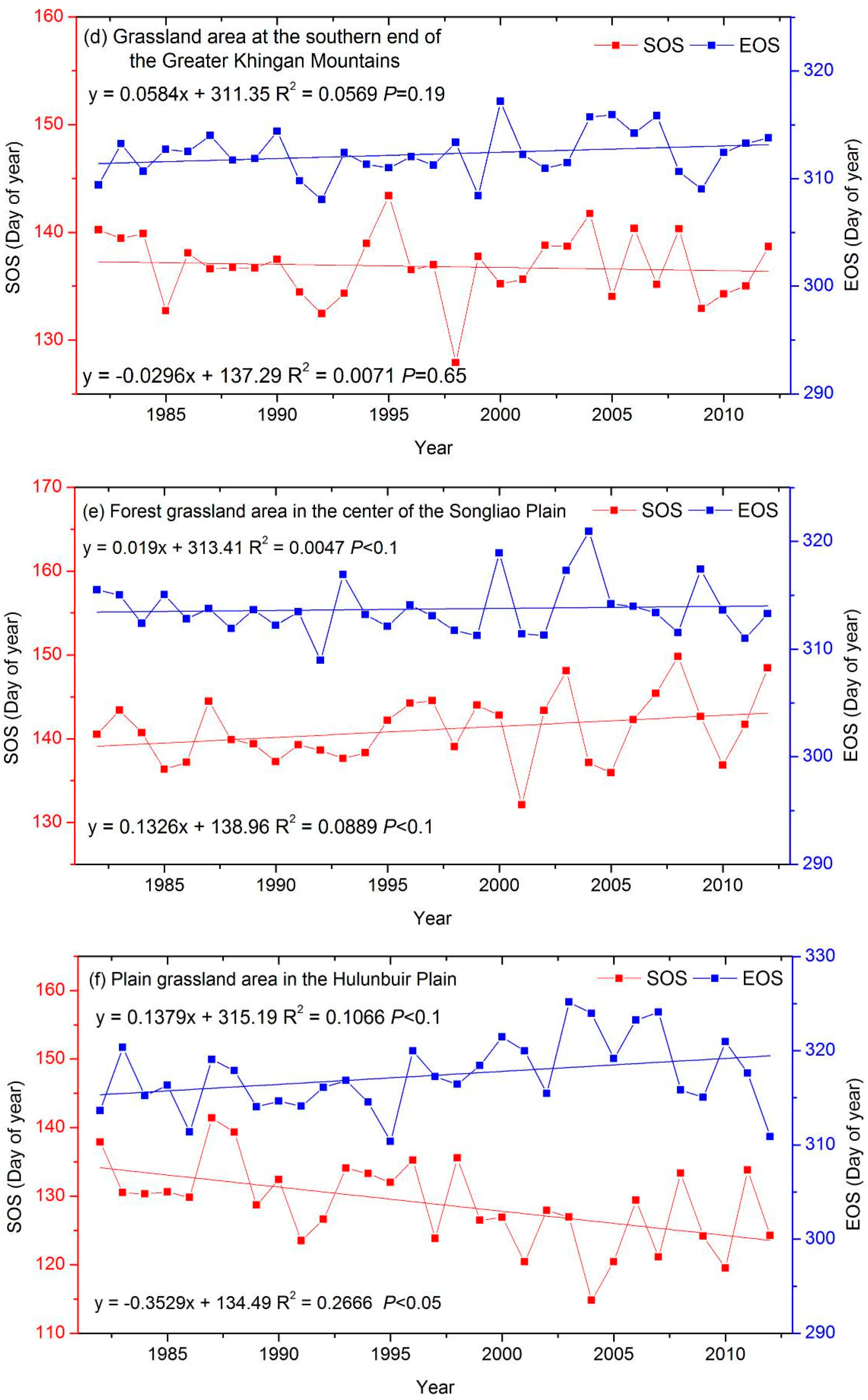
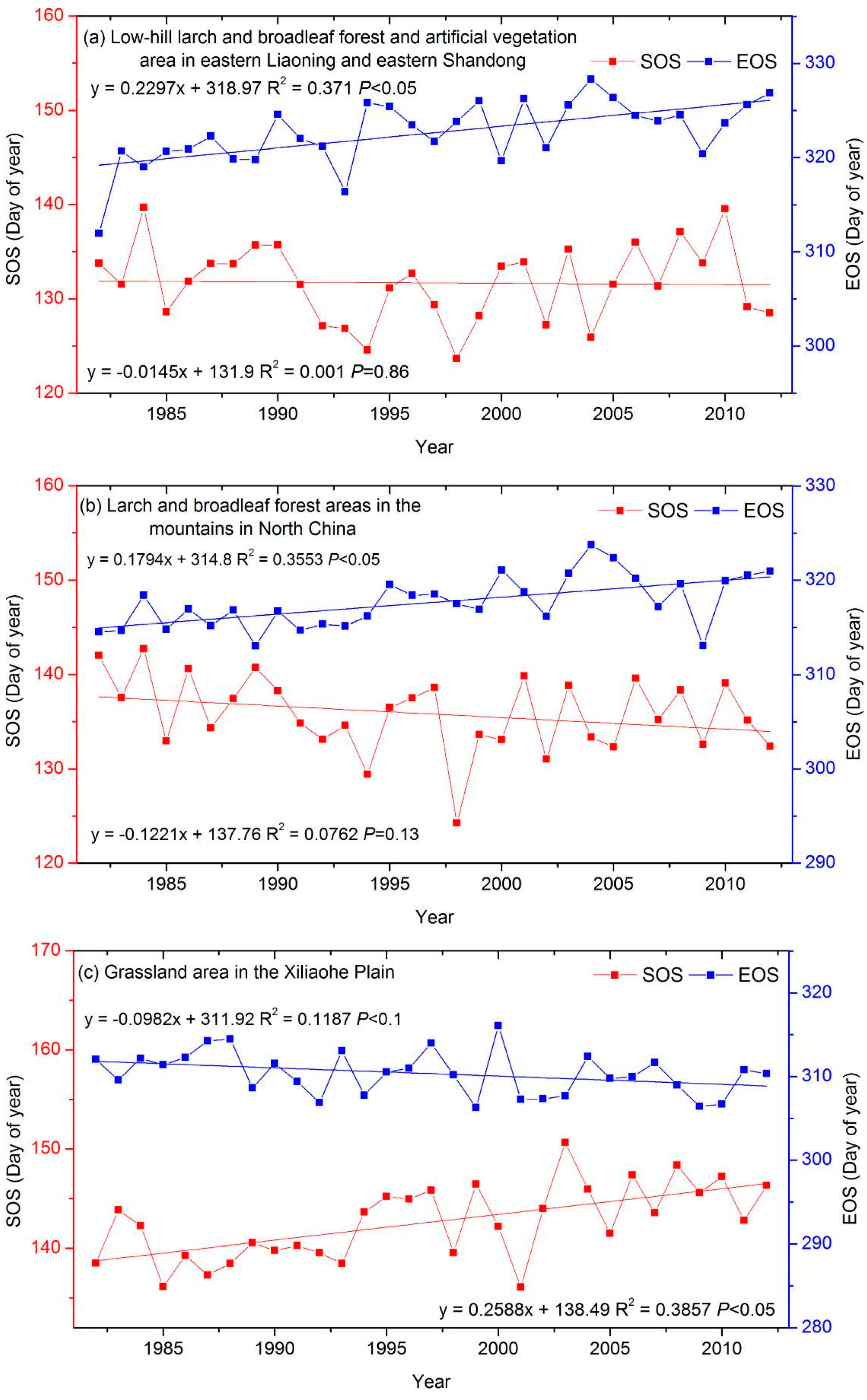
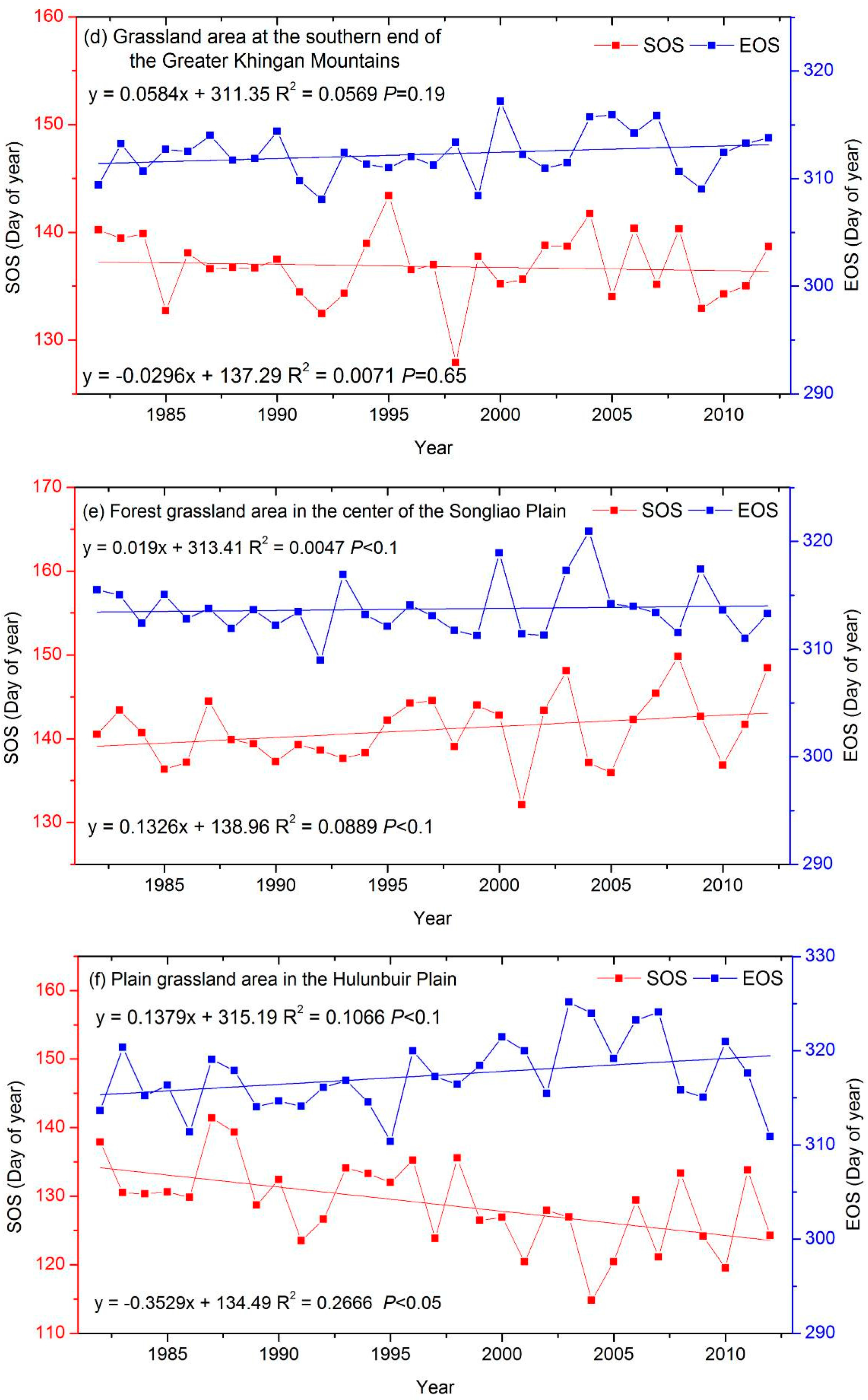
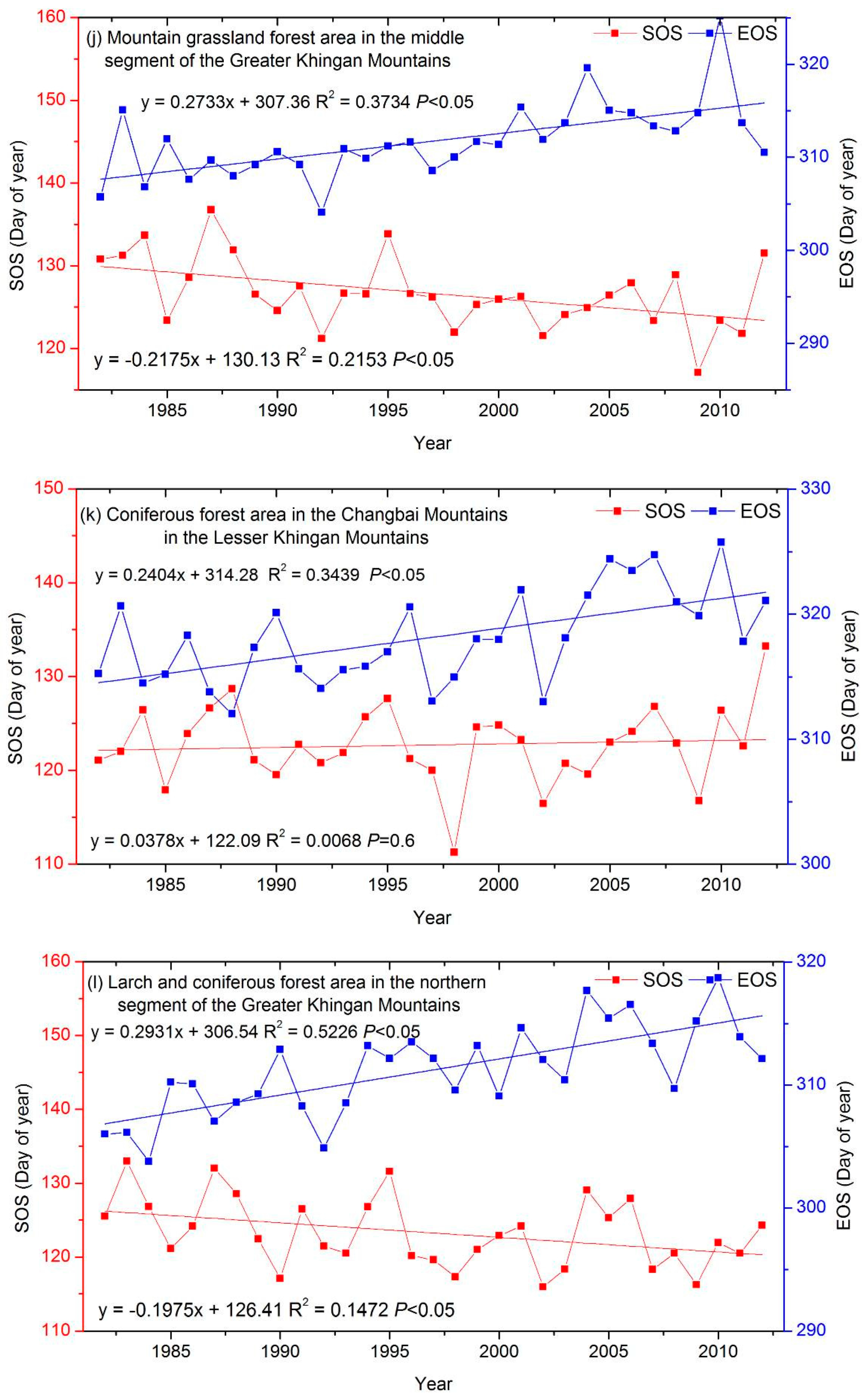
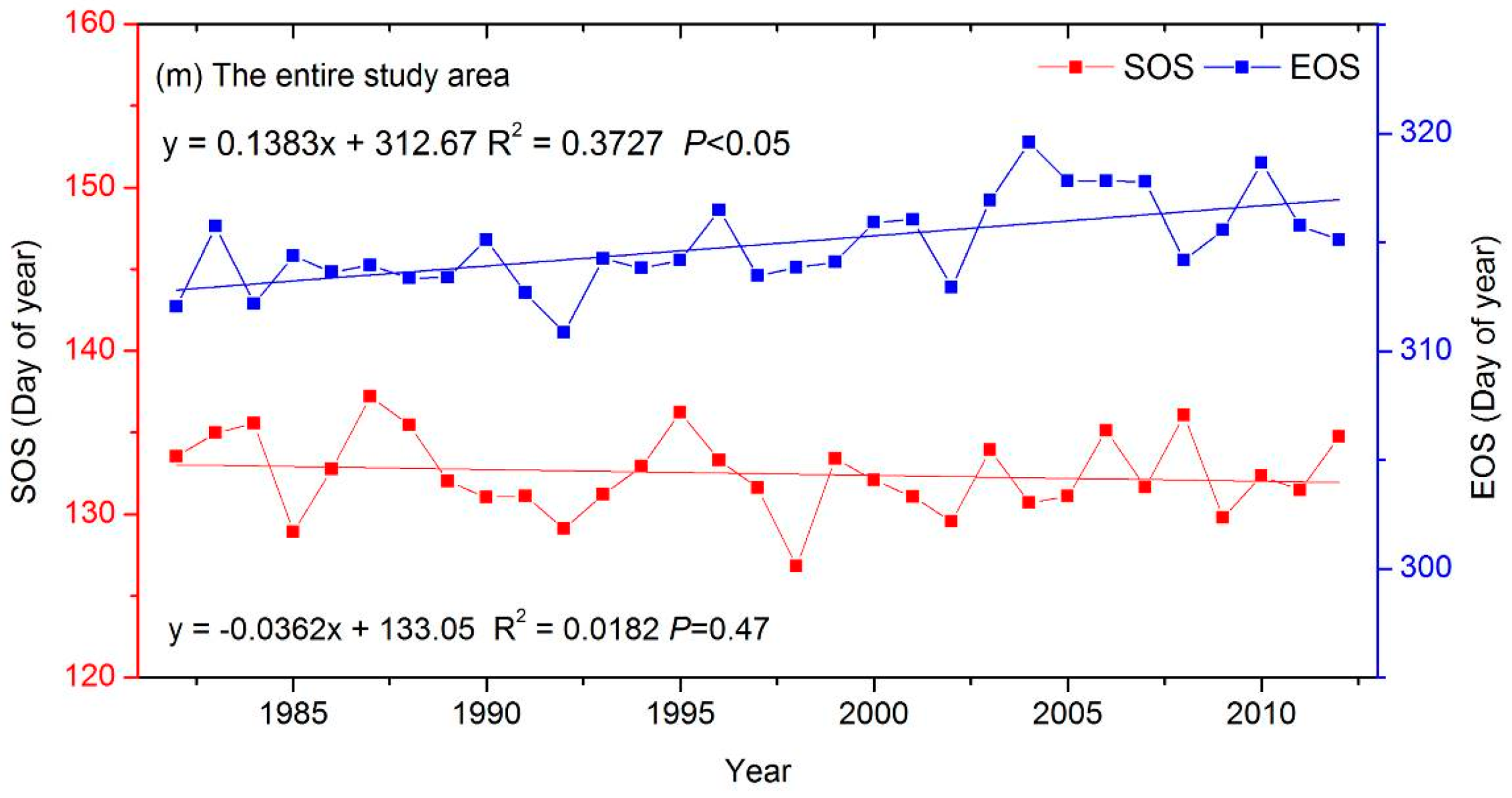
3.3. The Interannual Variability and Trends of LSP Metrics in Different Ecological Areas
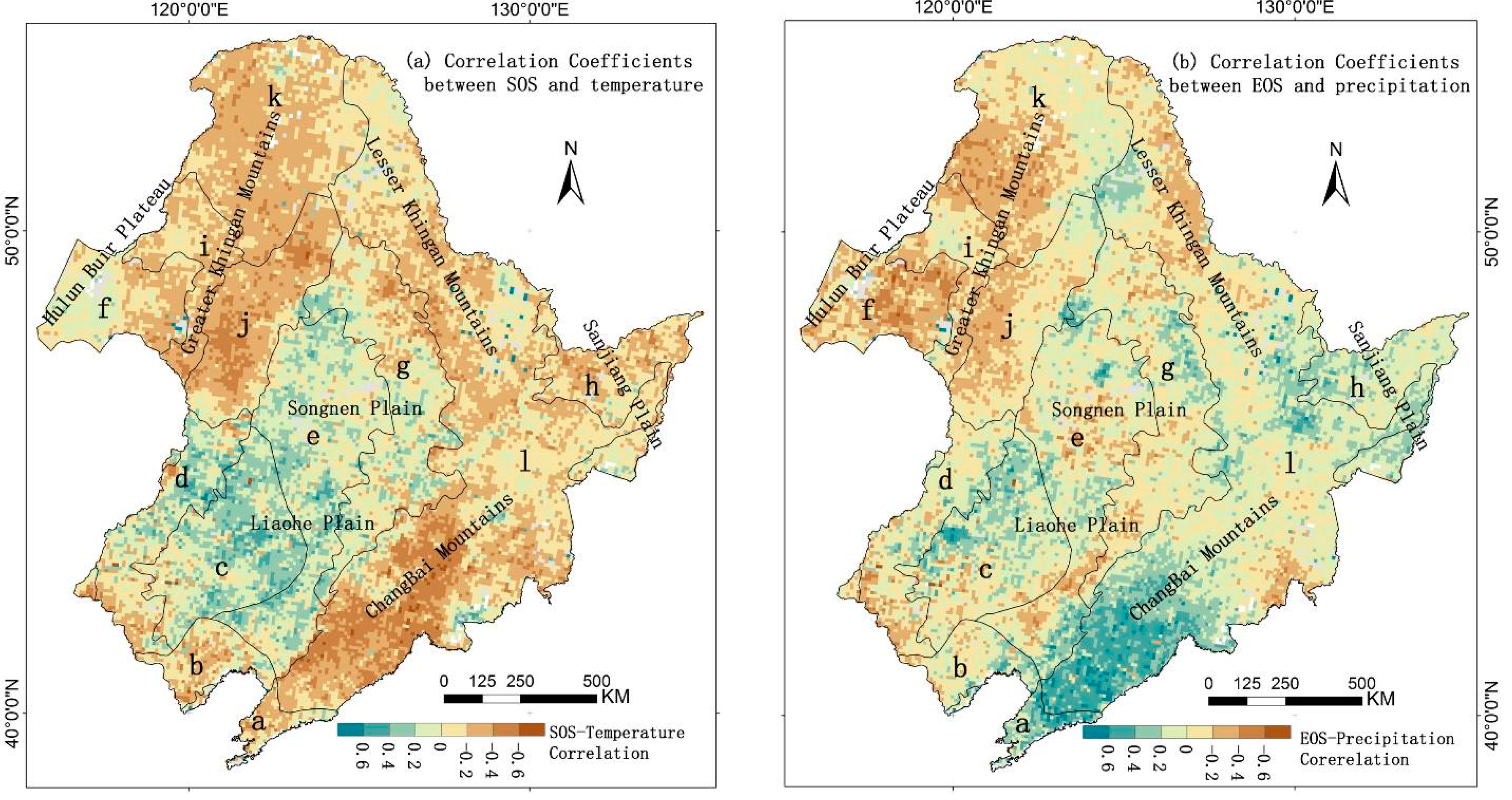
3.4. Direct Effects of Local Climate Factors on LSP
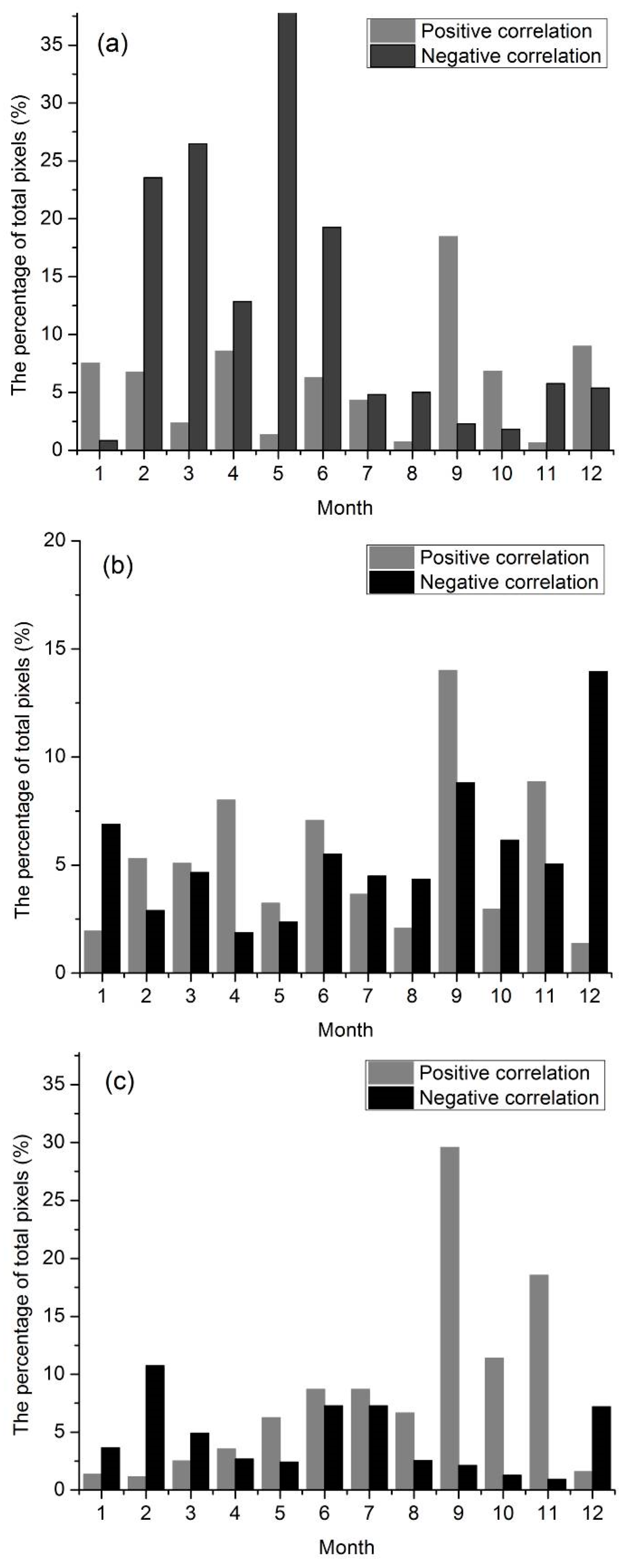
3.5. Lag Effect of Climate Change on Land Surface Phenology
4. Discussion
4.1. Variations of Land Surface Phenology
4.2. Relationships between LSP Metrics and Climatic Factors
4.3. Uncertainty
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Stöckli, R.; Vidale, P.L. European plant phenology and climate as seen in a 20-year AVHRR land-surface parameter dataset. Int. J. Remote Sens. 2004, 25, 3303–3330. [Google Scholar] [CrossRef]
- De Beurs, K.M.; Henebry, G.M. Land surface phenology, climatic variation, and institutional change: Analyzing agricultural land cover change in Kazakhstan. Remote Sens. Environ. 2004, 89, 497–509. [Google Scholar] [CrossRef]
- Bhatt, U.S.; Walker, D.A.; Raynolds, M.K.; Bieniek, P.A.; Epstein, H.E.; Comiso, J.C.; Pinzon, J.E.; Tucker, C.J.; Polyakov, I.V. Recent declines in warming and vegetation greening trends over Pan-Arctic tundra. Remote Sens. 2013, 5, 4229–4254. [Google Scholar] [CrossRef]
- Badeck, F.-W.; Bondeau, A.; Böttcher, K.; Doktor, D.; Lucht, W.; Schaber, J.; Sitch, S. Responses of spring phenology to climate change. New Phytol. 2004, 162, 295–309. [Google Scholar] [CrossRef]
- Cong, N.; Wang, T.; Nan, H.; Ma, Y.; Wang, X.; Myneni, R.B.; Piao, S. Changes in satellite-derived spring vegetation green-up date and its linkage to climate in China from 1982 to 2010: A multimethod analysis. Glob. Chang. Biol. 2013, 19, 881–891. [Google Scholar] [CrossRef] [PubMed]
- Heumann, B.W.; Seaquist, J.W.; Eklundh, L.; Jönsson, P. AVHRR derived phenological change in the Sahel and Soudan, Africa, 1982–2005. Remote Sens. Environ. 2007, 108, 385–392. [Google Scholar] [CrossRef]
- Justice, C.O.; Townshend, J.R.G.; Holben, B.N.; Tucker, C.J. Analysis of the phenology of global vegetation using meteorological satellite data. Int. J. Remote Sens. 1985, 6, 1271–1318. [Google Scholar] [CrossRef]
- Myneni, R.B.; Keeling, C.D.; Tucker, C.J.; Asrar, G.; Nemani, R.R. Increased plant growth in the northern high latitudes from 1981 to 1991. Nature 1997, 386, 698–702. [Google Scholar] [CrossRef]
- Lloyd, D. A phenological classification of terrestrial vegetation cover using shortwave vegetation index imagery. Int. J. Remote Sens. 1990, 11, 2269–2279. [Google Scholar] [CrossRef]
- Fischer, A. A model for the seasonal variations of vegetation indices in coarse resolution data and its inversion to extract crop parameters. Remote Sens. Environ. 1994, 48, 220–230. [Google Scholar] [CrossRef]
- Reed, B.C.; Brown, J.F.; VanderZee, D.; Loveland, T.R.; Merchant, J.W.; Ohlen, D.O. Measuring phenological variability from satellite imagery. J. Veg. Sci. 1994, 5, 703–714. [Google Scholar] [CrossRef]
- Zhou, L.; Tucker, C.J.; Kaufmann, R.K.; Slayback, D.; Shabanov, N.V.; Myneni, R.B. Variations in northern vegetation activity inferred from satellite data of vegetation index during 1981 to 1999. J. Geophys. Res. 2001, 106, 20069–20083. [Google Scholar] [CrossRef]
- Shabanov, N.V.; Zhou, L.; Knyazikhin, Y.; Myneni, R.B.; Tucker, C.J. Analysis of interannual changes in northern vegetation activity observed in AVHRR data from 1981 to 1994. IEEE Trans. Geosci. Remote Sens. 2002, 40, 115–130. [Google Scholar] [CrossRef]
- Zhou, L.; Kaufmann, R.K.; Tian, Y.; Myneni, R.B.; Tucker, C.J. Relation between interannual variations in satellite measures of northern forest greenness and climate between 1982 and 1999. J. Geophys. Res. 2003, 108. [Google Scholar] [CrossRef]
- Chen, X.; Hu, B.; Yu, R. Spatial and temporal variation of phenological growing season and climate change impacts in temperate eastern China. Glob. Chang. Biol. 2005, 11, 1118–1130. [Google Scholar] [CrossRef]
- Zhao, J.; Zhang, H.; Zhang, Z.; Guo, X.; Li, X.; Chen, C. Spatial and temporal changes in vegetation phenology at middle and high latitudes of the Northern Hemisphere over the past three decades. Remote Sens. 2015, 7, 10973–10995. [Google Scholar] [CrossRef]
- Cleland, E.E.; Chuine, I.; Menzel, A.; Mooney, H.A.; Schwartz, M.D. Shifting plant phenology in response to global change. Trends Ecol. Evol. 2007, 22, 357–365. [Google Scholar] [CrossRef] [PubMed]
- Julien, Y.; Sobrino, J.A. Global land surface phenology trends from GIMMS database. Int. J. Remote Sens. 2009, 30, 3495–3513. [Google Scholar] [CrossRef]
- Jeong, S.-J.; Ho, C.-H.; Gim, H.-J.; Brown, M.E. Phenology shifts at start vs. end of growing season in temperate vegetation over the Northern Hemisphere for the period 1982–2008. Glob. Chang. Biol. 2011, 17, 2385–2399. [Google Scholar] [CrossRef]
- White, M.A.; Running, S.W.; Thornton, P.E. The impact of growing-season length variability on carbon assimilation and evapotranspiration over 88 years in the eastern US deciduous forest. Int. J. Biometeorol. 1999, 42, 139–145. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Friedl, M.A.; Schaaf, C.B.; Strahler, A.H.; Liu, Z. Monitoring the response of vegetation phenology to precipitation in Africa by coupling MODIS and TRMM instruments. J. Geophys. Res. 2005, 110. [Google Scholar] [CrossRef]
- Fitter, A.H.; Fitter, R.S.R.; Harris, I.T.B.; Williamson, M.H. Relationships between first flowering date and temperature in the flora of a locality in central England. Funct. Ecol. 1995, 9, 55–60. [Google Scholar] [CrossRef]
- Rötzer, T.; Chmielewski, F.-M. Phenological maps of Europe. Clim. Res. 2001, 18, 249–257. [Google Scholar] [CrossRef]
- Piao, S.; Fang, J.; Zhou, L.; Ciais, P.; Zhu, B. Variations in satellite-derived phenology in China’s temperate vegetation. Glob. Chang. Biol. 2006, 12, 672–685. [Google Scholar] [CrossRef]
- Zhao, J.; Wang, Y.; Hashimoto, H.; Melton, F.S.; Hiatt, S.H.; Zhang, H.; Nemani, R.R. The variation of land surface phenology from 1982 to 2006 along the Appalachian Trail. IEEE Trans. Geosci. Remote Sens. 2013, 51, 2087–2095. [Google Scholar] [CrossRef]
- White, M.A.; Nemani, R.R.; Thornton, P.E.; Running, S.W. Satellite evidence of phenological differences between urbanized and rural areas of the eastern United States deciduous broadleaf forest. Ecosystems 2002, 5, 260–273. [Google Scholar] [CrossRef]
- Dash, J.; Jeganathan, C.; Atkinson, P.M. The use of MERIS Terrestrial Chlorophyll Index to study spatio-temporal variation in vegetation phenology over India. Remote Sens. Environ. 2010, 114, 1388–1402. [Google Scholar] [CrossRef]
- Busetto, L.; Colombo, R.; Migliavacca, M.; Cremonese, E.; Meroni, M.; Galvagno, M.; Rossini, M.; Siniscalco, C.; Morra di Cella, U.; Pari, E. Remote sensing of larch phenological cycle and analysis of relationships with climate in the Alpine region. Glob. Chang. Biol. 2010, 16, 2504–2517. [Google Scholar] [CrossRef]
- Yu, X.; Wang, Q.; Yan, H.; Wang, Y.; Wen, K.; Zhuang, D.; Wang, Q. Forest phenology dynamics and its responses to meteorological variations in Northeast China. Adv. Meteorol. 2014. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, W.-C.; Fang, X.; Ye, Y.; Zheng, J. Vegetation of Northeast China during the late seventeenth to early twentieth century as revealed by historical documents. Reg. Environ. Chang. 2011, 11, 869–882. [Google Scholar] [CrossRef]
- Zheng, D. China’s Eco-Geographical Region Map; The Commercial Press: Beijing, China, 2008. [Google Scholar]
- Zhu, Z.; Bi, J.; Pan, Y.; Ganguly, S.; Anav, A.; Xu, L.; Samanta, A.; Piao, S.; Nemani, R.R.; Myneni, R.B. Global data sets of vegetation leaf area index (LAI) 3g and Fraction of Photosynthetically Active Radiation (FPAR) 3g derived from Global Inventory Modeling and Mapping Studies (GIMMS) Normalized Difference Vegetation Index (NDVI3g) for the period 1981 to 2011. Remote Sens. 2013, 5, 927–948. [Google Scholar]
- Dardel, C.; Kergoat, L.; Hiernaux, P.; Mougin, E.; Grippa, M.; Tucker, C.J. Re-greening Sahel: 30 years of remote sensing data and field observations (Mali, Niger). Remote Sens. Environ. 2014, 140, 350–364. [Google Scholar] [CrossRef]
- NASA Ames Ecological Forecasting Lab. Available online: http://ecocast.arc.nasa.gov/data/pub/gimms/3g.v0/ (accessed on 15 September 2014).
- Chinese Meteorological Science Data Sharing Service Network. Available online: http://www.ncdc.noaa.gov/cag/time-series/global/ (accessed on 1 May 2015).
- Holben, B.N. Characteristics of maximum-value composite images from temporal AVHRR data. Int. J. Remote Sens. 2007, 7, 1417–1434. [Google Scholar] [CrossRef]
- Tucker, C.J.; Pinzon, J.E.; Brown, M.E.; Slayback, D.A.; Pak, E.W.; Mahoney, R. An extended AVHRR 8-km NDVI dataset compatible with MODIS and SPOT vegetation NDVI data. Int. J. Remote Sens. 2005, 26, 4485–4498. [Google Scholar] [CrossRef]
- Viovy, N.; Arino, O.; Belward, A.S. The Best Index Slope Extraction (BISE): A method for reducing noise in NDVI time-series. Int. J. Remote Sens. 1992, 13, 1585–1590. [Google Scholar] [CrossRef]
- Jiang, N.; Zhu, W.; Mou, M.; Wang, L.; Zhang, J. A phenology-preserving filtering method to reduce noise in NDVI time series. In Proceedings of the IEEE International Geoscience and Remote Sensing Symposium (IGARSS), Munich, Germany, 22–27 July 2012; pp. 2384–2387.
- Song, Y.; Chen, P.; Wan, Y.; Shen, S. Application of hybrid classification method based on fourier transform to time-series NDVI images. In Proceedings of the Congress on Image and Signal Processing, CISP ’08, Sanya, China, 27–30 May 2008; pp. 634–638.
- Chen, J.; Jönsson, P.; Tamura, M.; Gu, Z.; Matsushita, B.; Eklundh, L. A simple method for reconstructing a high-quality NDVI time-series data set based on the Savitzky-Golay filter. Remote Sens. Environ. 2004, 91, 332–344. [Google Scholar] [CrossRef]
- White, M.A.; de Beurs, K.M.; Didan, K.; Inouye, D.W.; Richardson, A.D.; Jensen, O.P.; O’Keefe, J.; Zhang, G.; Nemani, R.R.; van Leeuwen, W.J.D. Intercomparison, interpretation, and assessment of spring phenology in North America estimated from remote sensing for 1982–2006. Glob. Chang. Biol. 2009, 15, 2335–2359. [Google Scholar] [CrossRef]
- Brown, M.E.; Beurs, K.D.; Vrieling, A. The response of African land surface phenology to large scale climate oscillations. Remote Sens. Environ. 2010, 114, 2286–2296. [Google Scholar] [CrossRef]
- Jonsson, P.; Eklundh, L. TIMESAT—A program for analyzing time-series of satellite sensor data. Comput. Geosci. 2004, 30, 833–845. [Google Scholar] [CrossRef]
- Martínez, B.; Gilabert, M.A. Vegetation dynamics from NDVI time series analysis using the wavelet transform. Remote Sens. Environ. 2009, 113, 1823–1842. [Google Scholar] [CrossRef]
- Metsämäki, S.; Vepsäläinen, J.; Pulliainen, J.; Sucksdorff, Y. Improved linear interpolation method for the estimation of snow-covered area from optical data. Remote Sens. Environ. 2002, 82, 64–78. [Google Scholar] [CrossRef]
- Jonsson, P.; Eklundh, L. Seasonality extraction by function fitting to time-series of satellite sensor data. IEEE Trans. Geosci. Remote Sens. 2002, 40, 1824–1832. [Google Scholar] [CrossRef]
- Jönsson, A.M.; Eklundh, L.; Hellström, M.; Bärring, L.; Jönsson, P. Annual changes in MODIS vegetation indices of Swedish coniferous forests in relation to snow dynamics and tree phenology. Remote Sens. Environ. 2010, 114, 2719–2730. [Google Scholar] [CrossRef]
- Van Leeuwen, W.J.D. Monitoring the effects of forest restoration treatments on post-fire vegetation recovery with MODIS multitemporal data. Sensors 2008, 8, 2017–2042. [Google Scholar] [CrossRef]
- Bachoo, A.; Archibald, S. Influence of using date-specific values when extracting phenological metrics from 8-day composite NDVI data. In Proceedings of the IEEE International Workshop on the Analysis of Multi-temporal Remote Sensing Images, MultiTemp 2007, Leuven, Belgium, 18–20 July 2007; pp. 1–4.
- Hird, J.N.; McDermid, G.J. Noise reduction of NDVI time series: An empirical comparison of selected techniques. Remote Sens. Environ. 2009, 113, 248–258. [Google Scholar] [CrossRef]
- Peppin, D.; Fulé, P.Z.; Sieg, C.H.; Beyers, J.L.; Hunter, M.E. Post-wildfire seeding in forests of the western United States: An evidence-based review. For. Ecol. Manag. 2010, 260, 573–586. [Google Scholar] [CrossRef]
- Steenkamp, K.; Wessels, K.; Archibald, S.; von Maltitz, G. Long-term phenology and variability of Southern African vegetation. In Proceedings of the IEEE International Geoscience and Remote Sensing Symposium, IGARSS 2008, Boston, MA, USA, 7–11 July 2008; Volume 3, pp. 816–819.
- Eklundh, L.; Jonsson, P. TIMESAT 3.0 Software Manual; Malmö University: Malmö, Sweden, 2010. [Google Scholar]
- Gao, F.; Morisette, J.T.; Wolfe, R.E.; Ederer, G.; Pedelty, J.; Masuoka, E.; Myneni, R.; Tan, B.; Nightingale, J. An algorithm to produce temporally and spatially continuous MODIS-LAI time series. IEEE Geosci. Remote Sens. Lett. 2008, 5, 60–64. [Google Scholar] [CrossRef]
- Wang, M.; Tao, F. Comparison of three NDVI time-series fitting methods in crop phenology detection in Northeast China. In Proceedings of the IOP Conference Series: Earth and Environmental Science, Beijing, China, 22–26 April 2013; IOP Publishing: Beijing, China, 2014; Volume 17. [Google Scholar]
- Hou, X.H.; Niu, Z.; Gao, S. Phenology of forest vegetation in northeast of China in ten years using remote sensing. Spectrosc. Spectr. Anal. 2014, 34, 515–519. [Google Scholar]
- Fang, Y.X.; Fang, Z.D. Monitoring forest phenophases of Northeast China based on MODIS NDVI data. Resour. Sci. 2006, 28, 111–117. [Google Scholar]
- Gong, P.; Chen, Z.X. Regional vegetation phenology monitoring based on MODIS. Chin. J. Soil Sci. 2009, 40, 213–217. [Google Scholar]
- Luo, X.; Chen, X.; Wang, L.; Xu, L.; Tian, Y. Modeling and predicting spring land surface phenology of the deciduous broadleaf forest in northern China. Agric. For. Meteorol. 2014, 198–199, 33–41. [Google Scholar] [CrossRef]
- Liu, Q.; Fu, Y.H.; Zeng, Z.; Huang, M.; Li, X.; Piao, S. Temperature, precipitation, and insolation effects on autumn vegetation phenology in temperate China. Glob. Chang. Biol. 2016, 22, 644–655. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Guan, H.; Shen, M.; Liang, W.; Jiang, L. Changes in autumn vegetation dormancy onset date and the climate controls across temperate ecosystems in China from 1982 to 2010. Glob. Chang. Biol. 2015, 21, 652–665. [Google Scholar] [CrossRef] [PubMed]
- Xia, J.; Yan, Z. Changes in the local growing season in Eastern China during 1909–2012. Sola 2014, 10, 163–166. [Google Scholar] [CrossRef]
- Lucht, W.; Prentice, I.C.; Myneni, R.B.; Sitch, S.; Friedlingstein, P.; Cramer, W.; Bousquet, P.; Buermann, W.; Smith, B. Climatic Control of the High-Latitude Vegetation Greening Trend and Pinatubo Effect. Science 2002, 296, 1687–1689. [Google Scholar] [CrossRef] [PubMed]
- Jeong, S.-J.; Ho, C.-H.; Kim, B.-M.; Feng, S.; Medvigy, D. Non-linear response of vegetation to coherent warming over northern high latitudes. Remote Sens. Lett. 2013, 4, 123–130. [Google Scholar] [CrossRef]
- Barichivich, J.; Briffa, K.R.; Myneni, R.B.; Osborn, T.J.; Melvin, T.M.; Ciais, P.; Piao, S.; Tucker, C. Large-scale variations in the vegetation growing season and annual cycle of atmospheric CO2 at high northern latitudes from 1950 to 2011. Glob. Chang. Biol. 2013, 19, 3167–3183. [Google Scholar] [CrossRef] [PubMed]
- Berner, L.T.; Beck, P.S.; Bunn, A.G.; Lloyd, A.H.; Goetz, S.J. High-latitude tree growth and satellite vegetation indices: Correlations and trends in Russia and Canada (1982–2008). J. Geophys. Res. Biogeosci. 2011, 116. [Google Scholar] [CrossRef]
- Zeng, H.; Jia, G.; Epstein, H. Recent changes in phenology over the northern high latitudes detected from multi-satellite data. Environ. Res. Lett. 2011. [Google Scholar] [CrossRef]
- Uttam Babu, S.; Shiva, G.; Bawa, K.S. Widespread climate change in the Himalayas and associated changes in local ecosystems. PLoS ONE 2012, 7, e36741. [Google Scholar]
- Shen, Z.; Yin, R.; Qi, J. Land cover changes in Northeast China from the late 1970s to 2004. Appl. Ecol. Environ. Res. 2009, 11, 55–67. [Google Scholar]
- Yun, Y.R.; Fang, X.Q.; Wang, Y.; Tao, J.D.; Qiao, D.F. Main grain crops structural change and its climate background in Heilongjiang Province during the past two decades. J. Nat. Resour. 2005, 20, 697–705. [Google Scholar]
- Zhu, X.X.; Fang, X.Q.; Wang, Y. Responses of corn and rice planting area to temperature changes based on RS in the west of Heilongjiang Province. Sci. Geogr. Sin. 2008, 28, 66–71. [Google Scholar]
- Ma, S.Q.; Wang, Q.; Luo, X.L. Effect of climate change on maize (Zea mays) growth and yield based on stage sowing. Acta Ecol. Sin. 2008, 28, 2131–2139. [Google Scholar]
- Fang, X.Q.; Sheng, J.F. Human adaptation to climate change: A case study of changes in paddy planting area in Heilongjiang Province. J. Nat. Resour. 2000, 15, 213–217. [Google Scholar]
- Li, Z.-G.; Yang, P.; Tang, H.J.; Wu, W.-B.; Chen, Z.X.; Zhou, Q.B.; Zou, J.Q.; Zhang, L. Trend analysis of typical phenophases of major crops under climate change in the three provinces of Northeast China. Sci. Agric. Sin. 2011, 44, 4180–4189. [Google Scholar]
- Li, R.; Zhou, G.S. Responses of woody plants phenology to air temperature in Northeast China in 1980–2005. Chin. J. Ecol. 2010, 29, 2317–2326. [Google Scholar]
- Guo, Z.-X.; Zhang, X.-N.; Wang, Z.-M.; Fang, W.-H. Responses of vegetation phenology in Northeast China to climate change. Chin. J. Ecol. 2010, 29, 578–585. [Google Scholar]
- Li, Z.; Yang, P.; Tang, H.; Wu, W.; Yin, H.; Liu, Z.; Zhang, L. Response of maize phenology to climate warming in Northeast China between 1990 and 2012. Reg. Environ. Chang. 2014, 14, 39–48. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
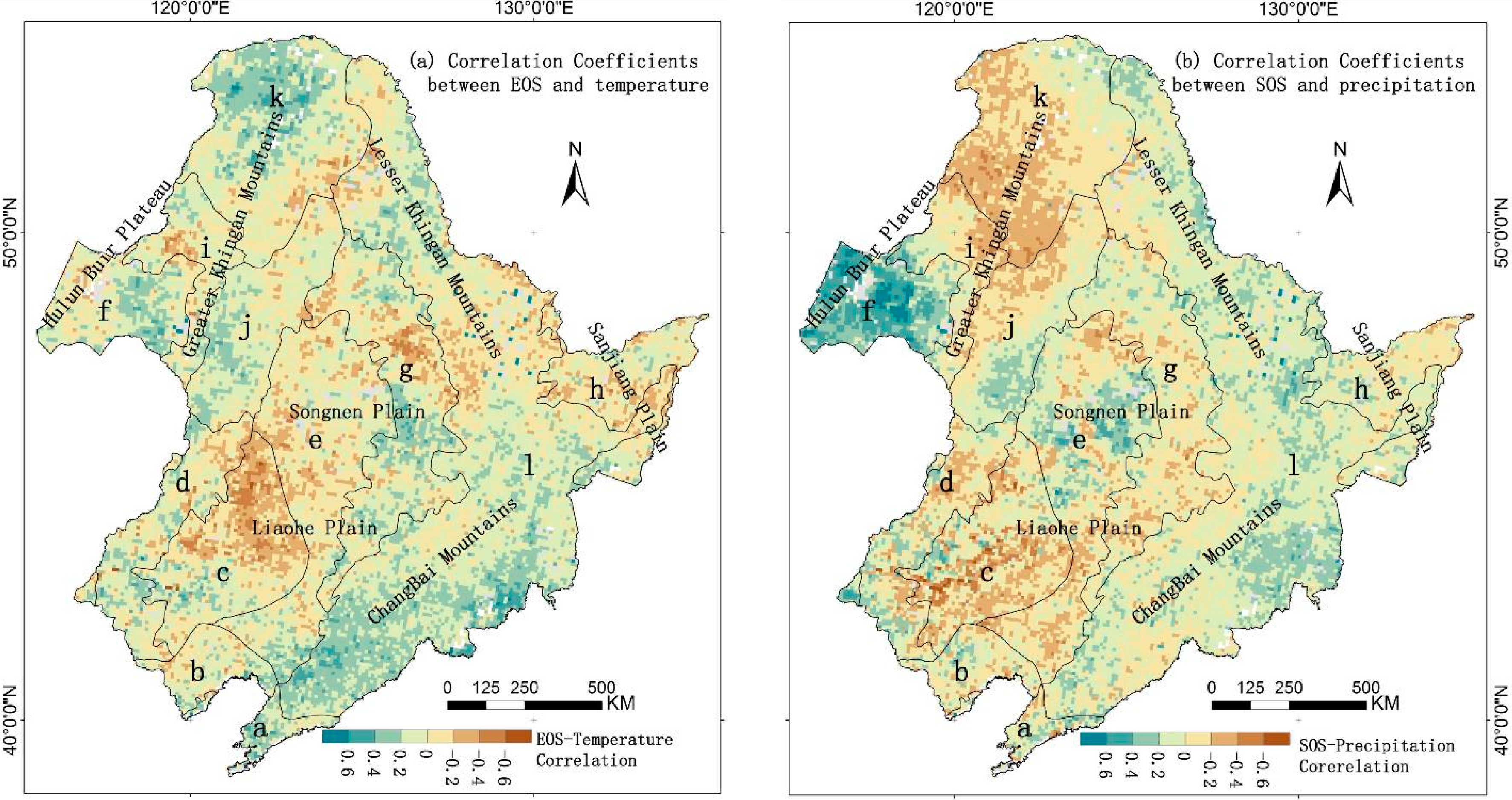{kind=link}
{kind=link}
{kind=link}
| Ecoregions | Trends (Days/Year) | |
|---|---|---|
| SOS | EOS | |
| low-hill larch and broadleaf forest and artificial vegetation in eastern Liaoning and eastern Shandong (LLS) | −0.01 | 0.23 ‡ |
| larch and broadleaf forest areas in the mountains of North China (LMN) | −0.12 | 0.18 ‡ |
| a grassland area in the Xiliaohe Plain (GXP) | 0.26 ‡ | −0.10 * |
| a grassland area at the southern end of the Greater Khingan Mountains (GGM) | −0.03 | 0.06 |
| a forest grassland area in the center of the Songliao Plain (FXP) | 0.13 * | 0.02 |
| a plain grassland area in the Hulunbuir Plain (PHP) | −0.35 ‡ | 0.14 * |
| a coniferous and broadleaf mixed forest area in the eastern piedmont tableland of the Songliao Plain (CSP) | 0.13 * | 0.09* |
| wetlands of the Sanjiang Plain (WSP) | 0.25 ‡ | 0.08 |
| a forest grassland area on the western side of the northern segment of the Greater Khingan Mountains (FGM) | −0.31 ‡ | 0.16 † |
| a mountain grassland forest area in the middle segment of the Greater Khingan Mountains (MGM) | −0.22 ‡ | 0.27 ‡ |
| a coniferous forest area in the Changbai Mountains in the Lesser Khingan Mountains (CLM) | 0.04 | 0.24 ‡ |
| a larch and coniferous forest area in the northern segment of the Greater Khingan Mountains (LGM) | −0.2 † | 0.29 ‡ |
| entire study area | −0.04 | 0.14 ‡ |
| Correlation | Spring | Summer | Fall | Winter | Year | |
|---|---|---|---|---|---|---|
| Temperature | Significant Positive Correlation | 3% | 3% | 2% | 5% | 4% |
| No Significant Positive Correlation | 26% | 40% | 42% | 67% | 32% | |
| No Significant Negative Correlation | 38% | 44% | 51% | 28% | 58% | |
| Significant Negative Correlation | 33% | 13% | 5% | 0% | 6% | |
| Precipitation | Significant Positive Correlation | 6% | 3% | 5% | 2% | 3% |
| No Significant Positive Correlation | 57% | 35% | 50% | 46% | 43% | |
| No Significant Negative Correlation | 33% | 59% | 42% | 50% | 51% | |
| Significant Negative Correlation | 4% | 3% | 3% | 2% | 2% |
| Correlation | Spring | Summer | Fall | Winter | Year | |
|---|---|---|---|---|---|---|
| Temperature | Significant Positive Correlation | 0% | 10% | 16% | 2% | 4% |
| No Significant Positive Correlation | 34% | 62% | 63% | 53% | 54% | |
| No Significant Negative Correlation | 60% | 27% | 20% | 42% | 40% | |
| Significant Negative Correlation | 6% | 1% | 1% | 3% | 2% | |
| Precipitation | Significant Positive Correlation | 11% | 1% | 5% | 18% | 3% |
| No Significant Positive Correlation | 57% | 45% | 32% | 58% | 41% | |
| No Significant Negative Correlation | 30% | 51% | 55% | 23% | 52% | |
| Significant Negative Correlation | 3% | 3% | 8% | 1% | 4% |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, J.; Wang, Y.; Zhang, Z.; Zhang, H.; Guo, X.; Yu, S.; Du, W.; Huang, F. The Variations of Land Surface Phenology in Northeast China and Its Responses to Climate Change from 1982 to 2013. Remote Sens. 2016, 8, 400. https://doi.org/10.3390/rs8050400
Zhao J, Wang Y, Zhang Z, Zhang H, Guo X, Yu S, Du W, Huang F. The Variations of Land Surface Phenology in Northeast China and Its Responses to Climate Change from 1982 to 2013. Remote Sensing. 2016; 8(5):400. https://doi.org/10.3390/rs8050400
Chicago/Turabian StyleZhao, Jianjun, Yanying Wang, Zhengxiang Zhang, Hongyan Zhang, Xiaoyi Guo, Shan Yu, Wala Du, and Fang Huang. 2016. "The Variations of Land Surface Phenology in Northeast China and Its Responses to Climate Change from 1982 to 2013" Remote Sensing 8, no. 5: 400. https://doi.org/10.3390/rs8050400
APA StyleZhao, J., Wang, Y., Zhang, Z., Zhang, H., Guo, X., Yu, S., Du, W., & Huang, F. (2016). The Variations of Land Surface Phenology in Northeast China and Its Responses to Climate Change from 1982 to 2013. Remote Sensing, 8(5), 400. https://doi.org/10.3390/rs8050400